Persistence of Norovirus GII Genome in Drinking Water and Wastewater at Different Temperatures


Abstract
:1. Introduction
2. Results
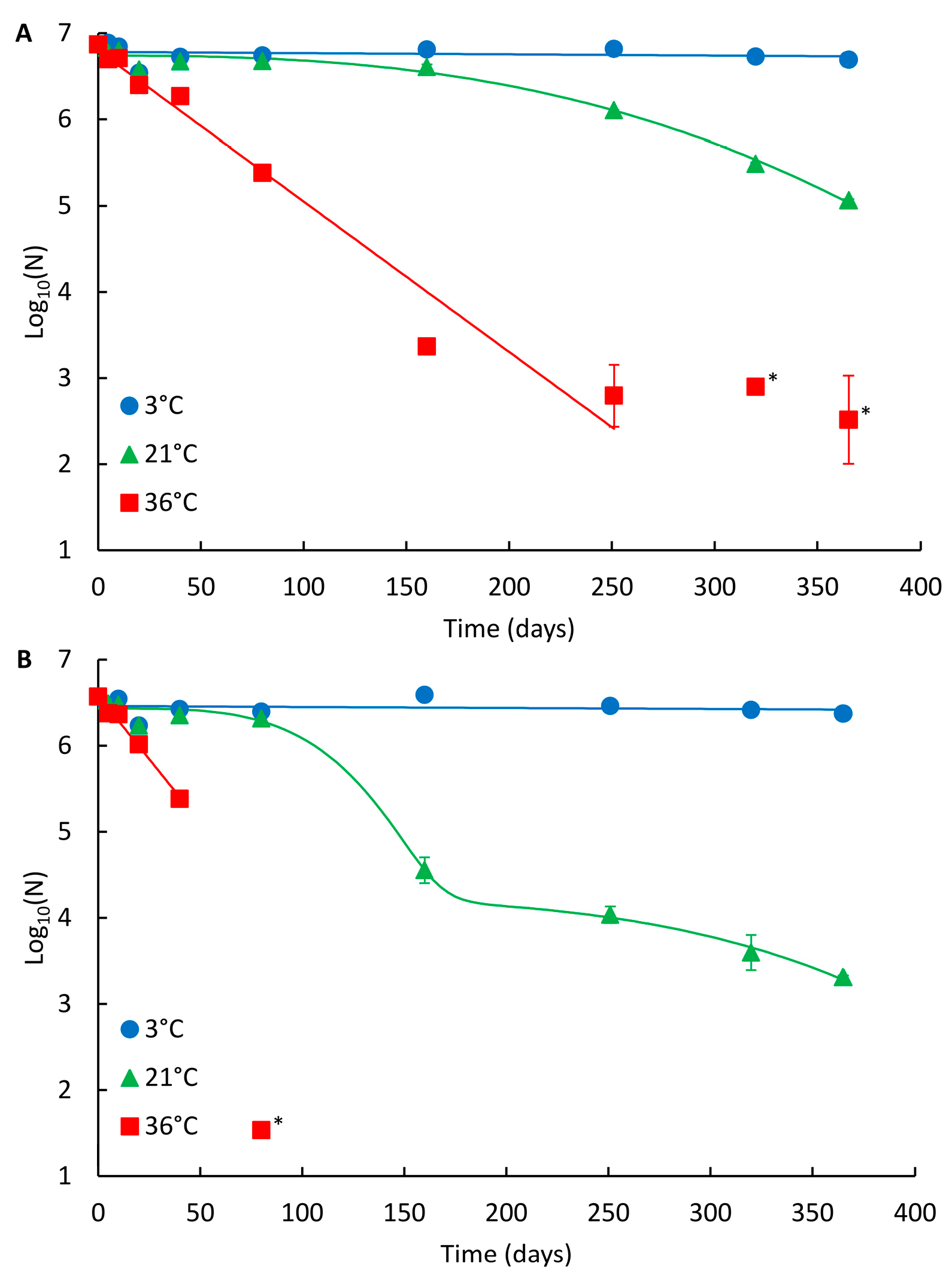
2.1. Persistence of the NoV Genome in Drinking Water at Different Temperatures
2.2. Persistence of the NoV Genome in Wastewater at Different Temperatures
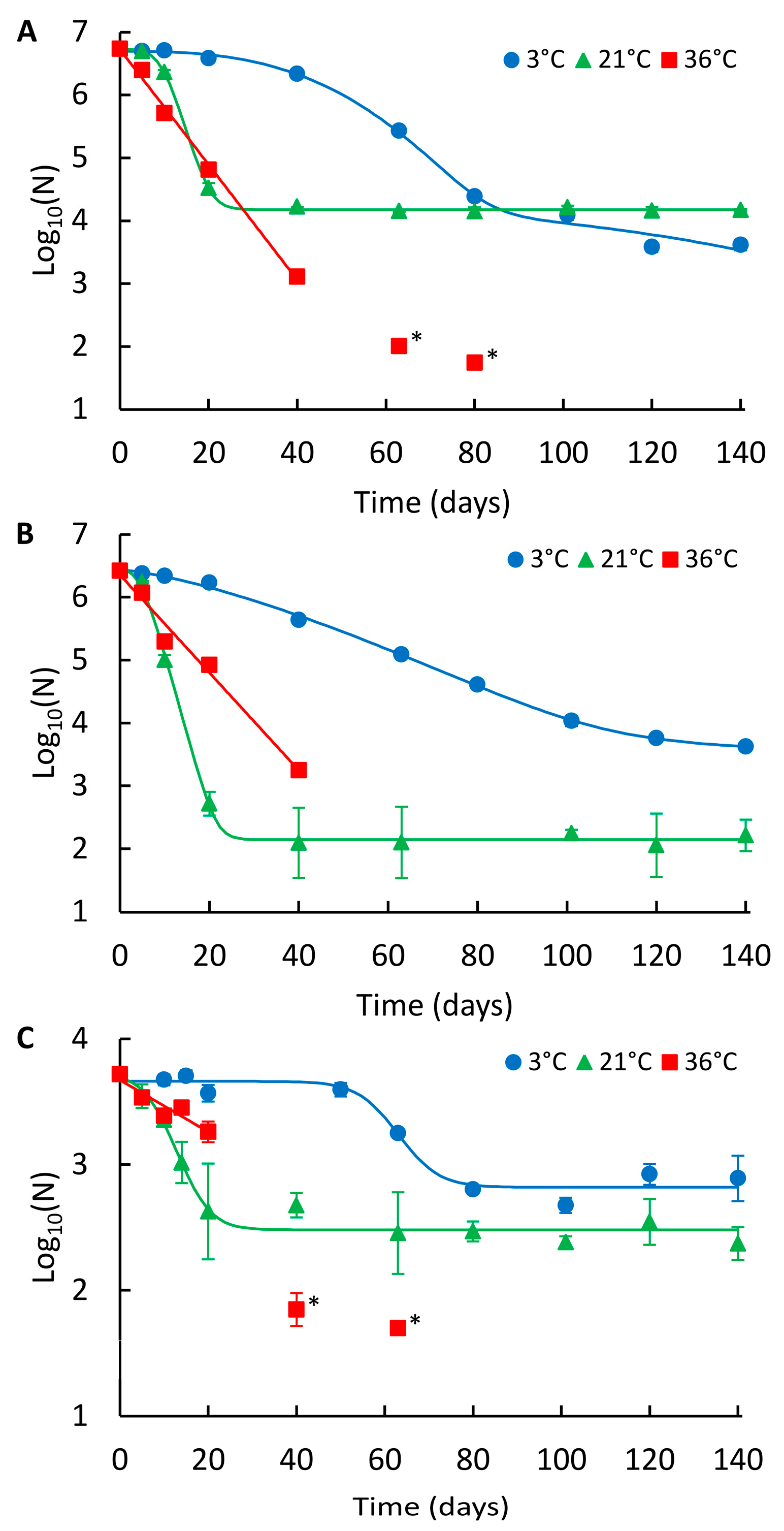
2.3. Effect of the Matrix on NoV Genome Persistence
3. Discussion
4. Materials and Methods
4.1. Viruses and Water Matrices
4.2. Experimental Design and Sampling
4.3. Quantitative Detection of NoV
4.4. Sequencing
4.5. Modelling of Decay Curves
4.6. Data Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zacheus, O.; Miettinen, I.T. Increased information on waterborne outbreaks through efficient notification system enforces actions towards safe drinking water. J. Water Health 2011, 9, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Belliot, G.; Lopman, B.A.; Ambert-Balay, K.; Pothier, P. The burden of norovirus gastroenteritis: An important foodborne and healthcare-related infection. Clin. Microbiol. Infect. 2014, 20, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Maunula, L.; Klemola, P.; Kauppinen, A.; Soderberg, K.; Ngujen, T.; Pitkänen, T.; Kaijalainen, S.; Simonen, M.L. Enteric viruses in a large waterborne outbreak of acute gastroenteritis in Finland. Food Environ. Virol. 2009, 1, 31–36. [Google Scholar] [CrossRef]
- Sinclair, R.G.; Jones, E.L.; Gerba, C.P. Viruses in recreational water-borne disease outbreaks: A review. J. Appl. Microbiol. 2009, 107, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Kauppinen, A.; Al-Hello, H.; Zacheus, O.; Kilponen, J.; Maunula, L.; Huusko, S.; Lappalainen, M.; Miettinen, I.; Blomqvist, S.; Rimhanen-Finne, R. Increase in outbreaks of gastroenteritis linked to bathing water in Finland in summer 2014. Euro Surveill. 2017, 22, 30470. [Google Scholar] [CrossRef] [PubMed]
- Kokkinos, P.; Kozyra, I.; Lazic, S.; Söderberg, K.; Vasickova, P.; Bouwknegt, M.; Rutjes, S.; Willems, K.; Moloney, R.; de Roda Husman, A.M.; et al. Virological Quality of Irrigation Water in Leafy Green Vegetables and Berry Fruits Production Chains. Food Environ. Virol. 2017, 9, 72–78. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific opinion on norovirus (NoV) in oysters: Methods, limits and control options. EFSA J. 2012, 10, 2500. [Google Scholar]
- Murphy, H. Persistence of pathogens in sewage and other water types. In Global Water Pathogens Project. Part 4 Management of Risk from Excreta and Wastewater; Rose, J.B., Jiménez-Cisneros, B., Yates, M., Eds.; Michigan State University: E. Lansing, MI, USA, 2017; Available online: http://www.waterpathogens.org/book/persistence-in-sewage (accessed on 2 October 2017).
- John, D.E.; Rose, J.B. Review of factors affecting microbial survival in groundwater. Environ. Sci. Technol. 2005, 39, 7345–7356. [Google Scholar] [CrossRef] [PubMed]
- Gerba, C.P. Virus occurrence and survival in the environmental waters. In Human Viruses in Water; Bosch, A., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 91–108. [Google Scholar]
- Yates, M. Persistence of pathogenic microorganisms in fecal wastes and wastewater matrices: An introduction and overview of data considerations. In Global Water Pathogens Project. Part 4 Management of Risk from Excreta and Wastewater; Rose, J.B., Jiménez-Cisneros, B., Yates, M., Eds.; Michigan State University: E. Lansing, MI, USA, 2017; Available online: http://www.waterpathogens.org/book/persistence-of-pathogens-introduction (accessed on 2 October 2017).
- Jones, M.K.; Grau, K.R.; Costantini, V.; Kolawole, A.O.; de Graaf, M.; Freiden, P.; Graves, C.L.; Koopmans, M.; Wallet, S.M.; Tibbetts, S.A.; et al. Human norovirus culture in B cells. Nat. Protoc. 2015, 10, 1939–1947. [Google Scholar] [CrossRef] [PubMed]
- Ettayebi, K.; Crawford, S.E.; Murakami, K.; Broughman, J.R.; Karandikar, U.; Tenge, V.R.; Neill, F.H.; Blutt, S.E.; Zeng, X.L.; Qu, L.; et al. Replication of human noroviruses in stem cell-derived human enteroids. Science 2016, 353, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Knight, A.; Haines, J.; Stals, A.; Li, D.; Uyttendaele, M.; Knight, A.; Jaykus, L.A. A systematic review of human norovirus survival reveals a greater persistence of human norovirus RT-qPCR signals compared to those of cultivable surrogate viruses. Int. J. Food Microbiol. 2016, 216, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Knight, A.; Li, D.; Uyttendaele, M.; Jaykus, L.A. A Critical Review of Methods for Detecting Human Noroviruses and Predicting Their Infectivity. Crit. Rev. Microbiol. 2013, 39, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.; Schwab, K.J. Evaluation of murine norovirus, feline calicivirus, poliovirus, and MS2 as surrogates for human norovirus in a model of viral persistence in surface water and groundwater. Appl. Environ. Microbiol. 2008, 74, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Charles, K.J.; Shore, J.; Sellwood, J.; Laverick, M.; Hart, A.; Pedley, S. Assessment of the stability of human viruses and coliphage in groundwater by PCR and infectivity methods. J. Appl. Microbiol. 2009, 106, 1827–1837. [Google Scholar] [CrossRef] [PubMed]
- Skraber, S.; Ogorzaly, L.; Helmi, K.; Maul, A.; Hoffmann, L.; Cauchie, H.M.; Gantzer, C. Occurrence and persistence of enteroviruses, noroviruses and F-specific RNA phages in natural wastewater biofilms. Water Res. 2009, 43, 4780–4789. [Google Scholar] [CrossRef] [PubMed]
- Seitz, S.R.; Leon, J.S.; Schwab, K.J.; Lyon, G.M.; Dowd, M.; McDaniels, M.; Abdulhafid, G.; Fernandez, M.L.; Lindesmith, L.C.; Baric, R.S.; et al. Norovirus infectivity in humans and persistence in water. Appl. Environ. Microbiol. 2011, 77, 6884–6888. [Google Scholar] [CrossRef] [PubMed]
- Ngazoa, E.S.; Fliss, I.; Jean, J. Quantitative study of persistence of human norovirus genome in water using TaqMan real-time RT-PCR. J. Appl. Microbiol. 2008, 104, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Jaykus, L.A.; Wong, E.; Moe, C. Persistence of Norwalk virus, male-specific coliphage, and Escherichia coli on stainless steel coupons and in phosphate-buffered saline. J. Food Prot. 2012, 75, 2151–2157. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, I.; Schijven, J.F.; Sánchez, G.; Wyn-Jones, P.; Ottoson, J.; Morin, T.; Muscillo, M.; Verani, M.; Nasser, A.; de Roda Husman, A.M.; et al. The impact of temperature on the inactivation of enteric viruses in food and water: A review. J. Appl. Microbiol. 2012, 112, 1059–1074. [Google Scholar] [CrossRef] [PubMed]
- Richards, G.P.; Watson, M.A.; Meade, G.K.; Hovan, G.L.; Kingsley, D.H. Resilience of Norovirus GII.4 to freezing and thawing: Implications for virus infectivity. Food Environ. Virol. 2012, 4, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Teunis, P.F.; Moe, C.L.; Liu, P.; Miller, S.E.; Lindesmith, L.; Baric, R.S.; Le Pendu, J.; Calderon, R.L. Norwalk virus: How infectious is it? J. Med. Virol. 2008, 80, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Mathijs, E.; Denayer, S.; Palmeira, L.; Botteldoorn, N.; Scipioni, A.; Vanderplasschen, A.; Thiry, E.; Dierick, K. Novel norovirus recombinants and of GII.4 sub-lineages associated with outbreaks between 2006 and 2010 in Belgium. Virol. J. 2011, 8, 310. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, D.; Mauroy, A.; Seebach, J.; Simon, V.; Wantia, N.; Protzer, U. New norovirus classified as a recombinant GII.g/GII.1 causes an extended foodborne outbreak at a university hospital in Munich. J. Clin. Virol. 2013, 58, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Loury, P.; Le Guyader, F.S.; Le Saux, J.C.; Ambert-Balay, K.; Parrot, P.; Hubert, B. A norovirus oyster-related outbreak in a nursing home in France, January 2012. Epidemiol. Infect. 2015, 143, 2486–2493. [Google Scholar] [CrossRef] [PubMed]
- Bruggink, L.D.; Dunbar, N.L.; Marshall, J.A. Emergence of GII.Pg norovirus in gastroenteritis outbreaks in Victoria, Australia. J. Med. Virol. 2016, 88, 1521–1528. [Google Scholar] [CrossRef] [PubMed]
- Verhaelen, K.; Bouwknegt, M.; Lodder-Verschoor, F.; Rutjes, S.A.; de Roda Husman, A.M. Persistence of human norovirus GII.4 and GI.4, murine norovirus, and human adenovirus on soft berries as compared with PBS at commonly applied storage conditions. Int. J. Food Microbiol. 2012, 160, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, H.; D’Souza, D.H.; Davidson, P.M. A comparison of the thermal inactivation kinetics of human norovirus surrogates and hepatitis A virus in buffered cell culture medium. Food Microbiol. 2014, 42, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Kim, S.J.; Kang, D.H. Inactivation modeling of human enteric virus surrogates, MS2, Qβ, and ΦX174, in water using UVC-LEDs, a novel disinfecting system. Food Res. Int. 2017, 91, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Kauppinen, A.; Pitkänen, T.; Miettinen, I.T. Persistent norovirus contamination of groundwater supplies in two waterborne outbreaks. Food Environ. Virol. 2017. [Google Scholar] [CrossRef]
- Lopman, B.; Gastañaduy, P.; Park, G.W.; Hall, A.J.; Parashar, U.D.; Vinjé, J. Environmental transmission of norovirus gastroenteritis. Curr. Opin. Virol. 2012, 2, 96–102. [Google Scholar] [CrossRef] [PubMed]
- The Finnish Meteorological Institute’s Open Data. Available online: http://suja.kapsi.fi/fmi-tilastot.php (accessed on 10 August 2017).
- Drinking Water Quality in Kuopio in the Year 2016. Available online: http://www.kuopionvesi.fi/c/document_library/get_file?uuid=e825c3bf-f87a-4012-b4f3-65e3c6453c03&groupId=518539 (accessed on 7 September 2017).
- Loisy, F.; Atmar, R.L.; Guillon, P.; Le Cann, P.; Pommepuy, M.; Le Guyader, F.S. Real-time RT-PCR for norovirus screening in shellfish. J. Virol. Methods 2005, 123, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, T.; Kojima, S.; Shinohara, M.; Uchida, K.; Fukushi, S.; Hoshino, F.B.; Takeda, N.; Katayama, K. Broadly reactive and highly sensitive assay for Norwalk-like viruses based on real-time quantitative reverse transcription-PCR. J. Clin. Microbiol. 2003, 41, 1548–1557. [Google Scholar] [CrossRef] [PubMed]
- Vinjé, J.; Hamidjaja, R.A.; Sobsey, M.D. Development and application of a capsid VP1 (region D) based reverse transcription PCR assay for genotyping of genogroup I and II noroviruses. J. Virol. Methods 2004, 116, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Kojima, S.; Kageyama, T.; Fukushi, S.; Hoshino, F.B.; Shinohara, M.; Uchida, K.; Natori, K.; Takeda, N.; Katayama, K. Genogroup-specific PCR primers for detection of Norwalk-like viruses. J. Virol. Methods 2002, 100, 107–114. [Google Scholar] [CrossRef]
- Kroneman, A.; Vennema, H.; Deforche, K.; Avoort, H.V.; Peñaranda, S.; Oberste, M.S.; Vinjé, J.; Koopmans, M. An automated genotyping tool for enteroviruses and noroviruses. J. Clin. Virol. 2011, 51, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Geeraerd, A.H.; Valdramidis, V.P.; Van Impe, J.F. GInaFiT, a freeware tool to assess non-log-linear microbial survivor curves. Int. J. Food. Microbiol. 2005, 102, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Ratkowsky, D.A. Model Fitting and Uncertainty. In Modeling Microbial Responses in Food; McKellar, R.C., Lu, X., Eds.; CRC Press: Boca Raton, FL, USA, 2003; pp. 151–196. [Google Scholar]
- Geeraerd, A.H.; Herremans, C.H.; Van Impe, J.F. Structural model requirements to describe microbial inactivation during a mild heat treatment. Int. J. Food Microbiol. 2000, 59, 185–209. [Google Scholar] [CrossRef]
- Mafart, P.; Couvert, O.; Gaillard, S.; Leguerinel, I. On calculating sterility in thermal preservation methods: Application of the Weibull frequency distribution model. Int. J. Food Microbiol. 2002, 72, 107–113. [Google Scholar] [CrossRef]
- Van Boekel, M.A. On the use of the Weibull model to describe thermal inactivation of microbial vegetative cells. Int. J. Food Microbiol. 2002, 74, 139–159. [Google Scholar] [CrossRef]
- Coroller, L.; Leguerinel, I.; Mettler, E.; Savy, N.; Mafart, P. General model, based on two mixed weibull distributions of bacterial resistance, for describing various shapes of inactivation curves. Appl. Environ. Microbiol. 2006, 72, 6493–6502. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Log-Linear Model | Non-Linear Model | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| T | Water | Virus | kmax | R2 | RMSE | R2 | RMSE | ||||
| 3 °C | DW | GII_A | N/A 1 | N/A | |||||||
| GII_B | N/A | N/A | |||||||||
| Double Weibull | |||||||||||
| α | δ1 | p | δ2 | ||||||||
| WW | GII_A | 0.06 ± 0.001 | 0.957 | 0.30 | 2.46 ± 0.20 | 57.06 ± 2.68 | 2.80 ± 0.44 | 158.5 ± 21.9 | 0.996 | 0.11 | |
| GII_B | 0.05 ± 0.002 | 0.986 | 0.14 | 2.63 ± 0.56 | 50.06 ± 1.94 | 1.38 ± 0.11 | 453.3 ± 959.1 | 0.999 | 0.05 | ||
| Log-linear shoulder tail | |||||||||||
| S1 | kmax | Log10(Nres) | |||||||||
| GII_ind | 0.02 ± 0.003 | 0.801 | 0.20 | 58.85 ± 3.64 | 0.23 ± 0.17 | 2.82 ± 0.05 | 0.967 | 0.09 | |||
| Weibull | |||||||||||
| δ | p | ||||||||||
| 21 °C | DW | GII_A | 0.01 ± 0.001 | 0.885 | 0.22 | 298.2 ± 8.6 | 2.63 ± 0.36 | 0.983 | 0.09 | ||
| Double Weibull | |||||||||||
| α | δ1 | p | δ2 | ||||||||
| GII_B | 0.02 ± 0.002 | 0.962 | 0.28 | 2.20 ± 0.20 | 131.6 ± 10.3 | 3.84 ± 1.24 | 370.3 ± 25.7 | 0.995 | 0.11 | ||
| Log-linear shoulder tail | |||||||||||
| S1 | kmax | Log10(Nres) | |||||||||
| WW | GII_A | 0.04 ± 0.010 | 0.577 | 0.80 | 9.42 ± 0.30 | 0.54 ± 0.02 | 4.18 ± 0.01 | 0.999 | 0.03 | ||
| GII_B | 0.06 ± 0.020 | 0.544 | 1.35 | 4.71 ± 0.73 | 0.59 ± 0.03 | 2.14 ± 0.04 | 0.998 | 0.10 | |||
| GII_ind | 0.02 ± 0.005 | 0.636 | 0.31 | 8.40 ± 2.25 | 0.30 ± 0.09 | 2.48 ± 0.04 | 0.972 | 0.10 | |||
| 36 °C | DW | GII_A | 0.04 ± 0.003 | 0.967 | 0.31 | N/A | |||||
| GII_B | 0.07 ± 0.004 | 0.988 | 0.06 | N/A | |||||||
| WW | GII_A | 0.21 ± 0.010 | 0.996 | 0.11 | N/A | ||||||
| GII_B | 0.18 ± 0.010 | 0.982 | 0.19 | N/A | |||||||
| GII_ind | 0.05 ± 0.010 | 0.880 | 0.07 | N/A | |||||||
| Log-Linear | Non-Linear | ||||||
|---|---|---|---|---|---|---|---|
| T | Water | Virus | Best Fitting Model | T90 | T99.99 | TFL | T4L |
| 3 °C | DW | GII_A | N/A 1 | Na2 | Na | Na | Na |
| GII_B | N/A | Na | Na | Na | Na | ||
| WW | GII_A | Double Weibull | 38 | 154 | 57 | 185 | |
| GII_B | Double Weibull | 45 | 179 | 50 | 569 | ||
| GII_ind | Log-linear shoulder tail | 115 | 461 | Na | Na | ||
| 21 °C | DW | GII_A | Weibull | 230 | 921 | 298 | 505 |
| GII_B | Double Weibull | 115 | 461 | 132 | 431 | ||
| WW | GII_A | Log-linear shoulder tail | 58 | 230 | 14 | Na | |
| GII_B | Log-linear shoulder tail | 38 | 154 | 8 | 21 | ||
| GII_ind | Log-linear shoulder tail | 115 | 461 | 19 | Na | ||
| 36 °C | DW | GII_A | Log-linear | 58 | 230 | N/A | N/A |
| GII_B | Log-linear | 33 | 132 | N/A | N/A | ||
| WW | GII_A | Log-linear | 11 | 44 | N/A | N/A | |
| GII_B | Log-linear | 13 | 51 | N/A | N/A | ||
| GII_ind | Log-linear | 46 | 184 | N/A | N/A | ||
| GII_A | GII_B | |
|---|---|---|
| Patient age and gender | 29 years Female | 2 years Female |
| Assumed exposure | 1 April 2011 | Not known |
| Symptoms started | 2 April 2011 | 23–24 February 2012 |
| Stool sample taken | 3 April 2011 | 25 February 2012 |
| Symptoms relieved | 4 April 2011 | 26 February 2012 |
| Storage prior to preparation of inoculum | Immediately at −20 °C (five years and 26 days) | One week outdoors at an average temperature of −4.6 °C (range −20–5.2 °C) [34], including three putative freeze thaw cycles, and then 4 March 2012 at −20 °C (four years, one month and 25 days) |
| Drinking Water [35] | Wastewater | |
|---|---|---|
| Turbidity, FTU | 0.1 | 23.15 |
| Color, mg Pt L−1 | <5 | Nd 1 |
| pH | 7.7 | 7.03 |
| Conductivity, μS cm−1 | 263 | 1026 |
| TOC 2, mg C L−1 | 2.1 | Nd |
| Free chlorine, mg Cl2 L−1 | 0.33 | Nd |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kauppinen, A.; Miettinen, I.T. Persistence of Norovirus GII Genome in Drinking Water and Wastewater at Different Temperatures. Pathogens 2017, 6, 48. https://doi.org/10.3390/pathogens6040048
Kauppinen A, Miettinen IT. Persistence of Norovirus GII Genome in Drinking Water and Wastewater at Different Temperatures. Pathogens. 2017; 6(4):48. https://doi.org/10.3390/pathogens6040048
Chicago/Turabian StyleKauppinen, Ari, and Ilkka T. Miettinen. 2017. "Persistence of Norovirus GII Genome in Drinking Water and Wastewater at Different Temperatures" Pathogens 6, no. 4: 48. https://doi.org/10.3390/pathogens6040048
APA StyleKauppinen, A., & Miettinen, I. T. (2017). Persistence of Norovirus GII Genome in Drinking Water and Wastewater at Different Temperatures. Pathogens, 6(4), 48. https://doi.org/10.3390/pathogens6040048