Species C Rotaviruses in Children with Diarrhea in India, 2010–2013: A Potentially Neglected Cause of Acute Gastroenteritis


 ,
, 

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Samples
2.2. Viral RNA Extraction and cDNA Synthesis
2.3. Diagnostic VP6 Gene-Specific RT-PCR Assay
2.4. Amplification, Cloning and Sequencing of VP6, VP4, VP7 and NSP4 Genes
2.5. Phylogenetic Analysis and Pair-Wise Sequence Distance for VP6, VP4 and NSP4 Genes
2.6. Determination of N-Glycosylation Sites
3. Results and Discussion
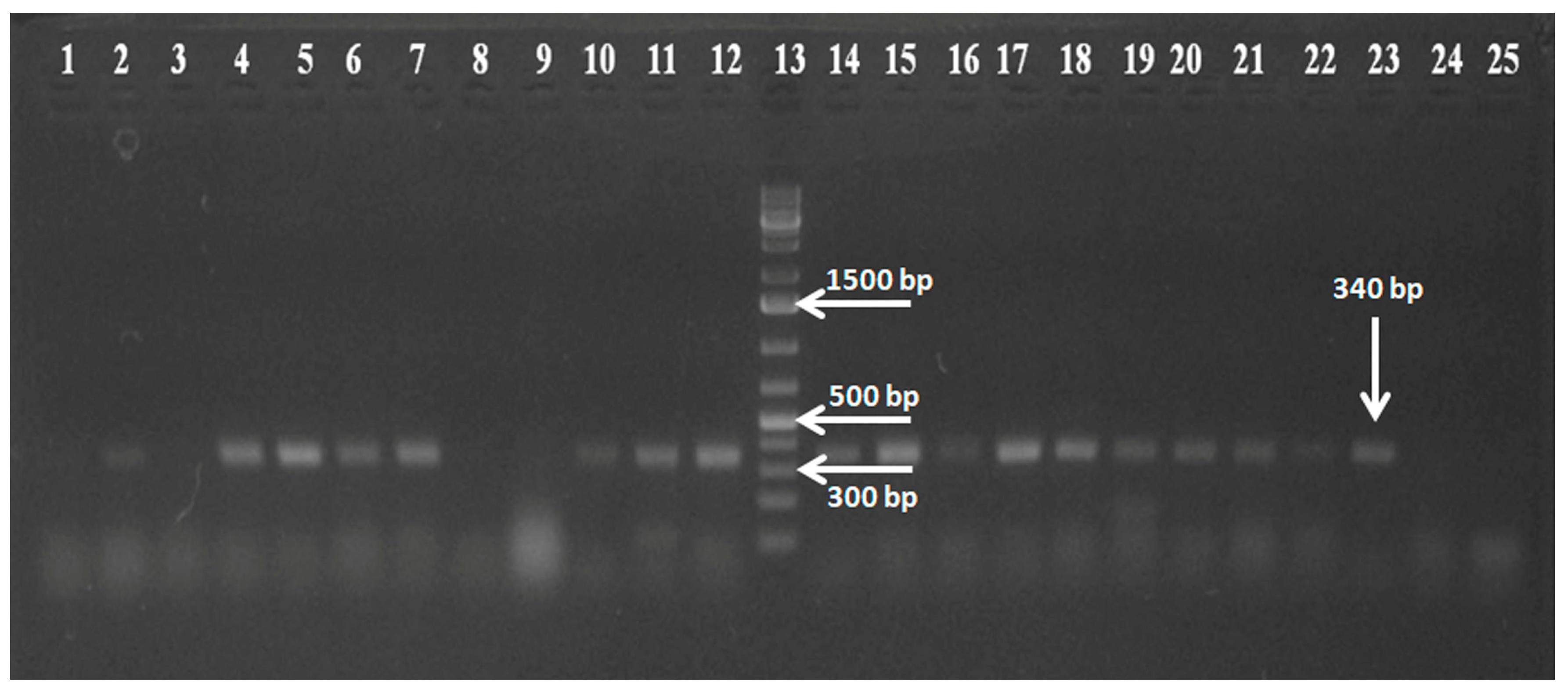
3.1. Detection of RVC
3.2. Nucleotide and Amino Acid Sequence Analysis
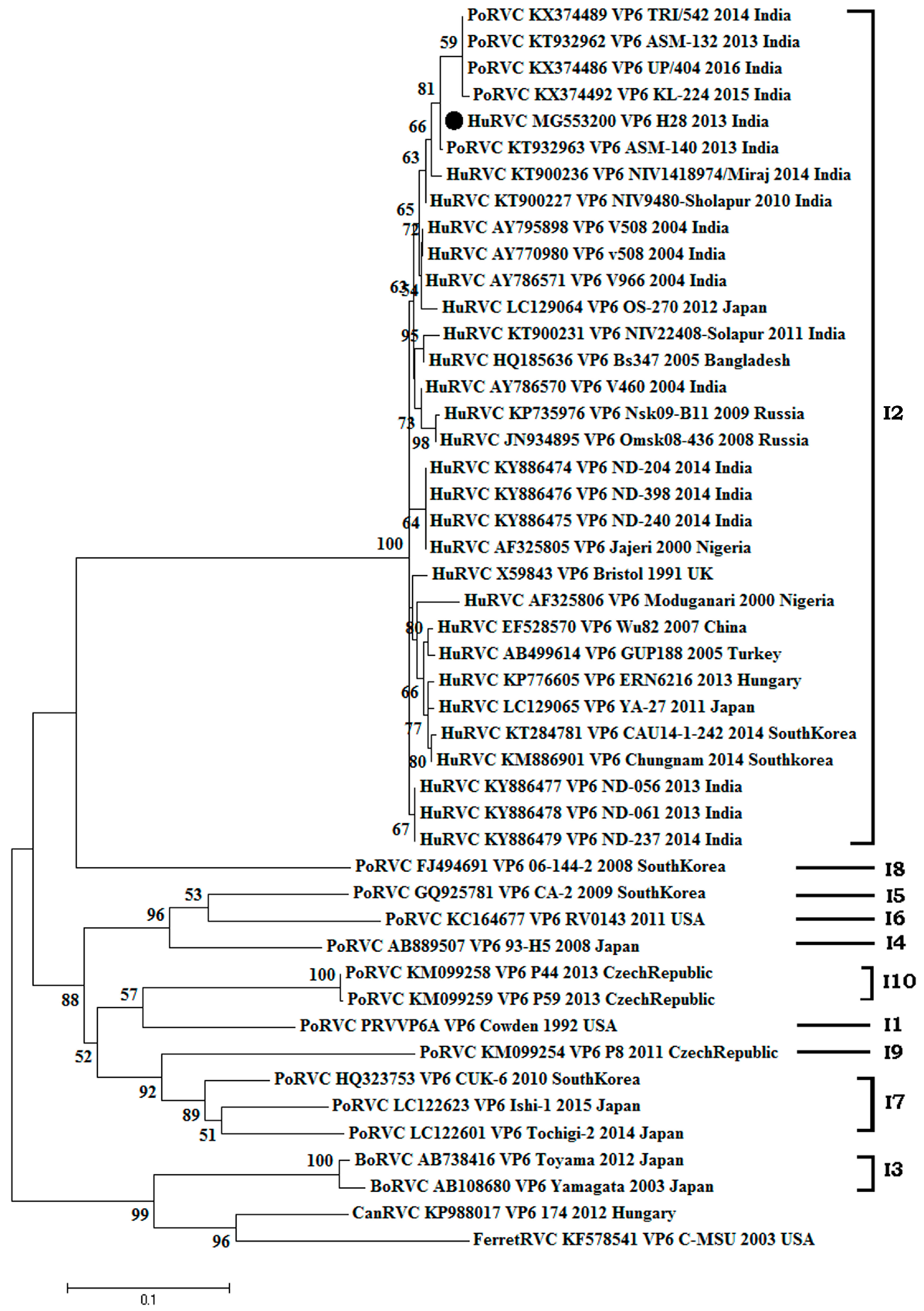
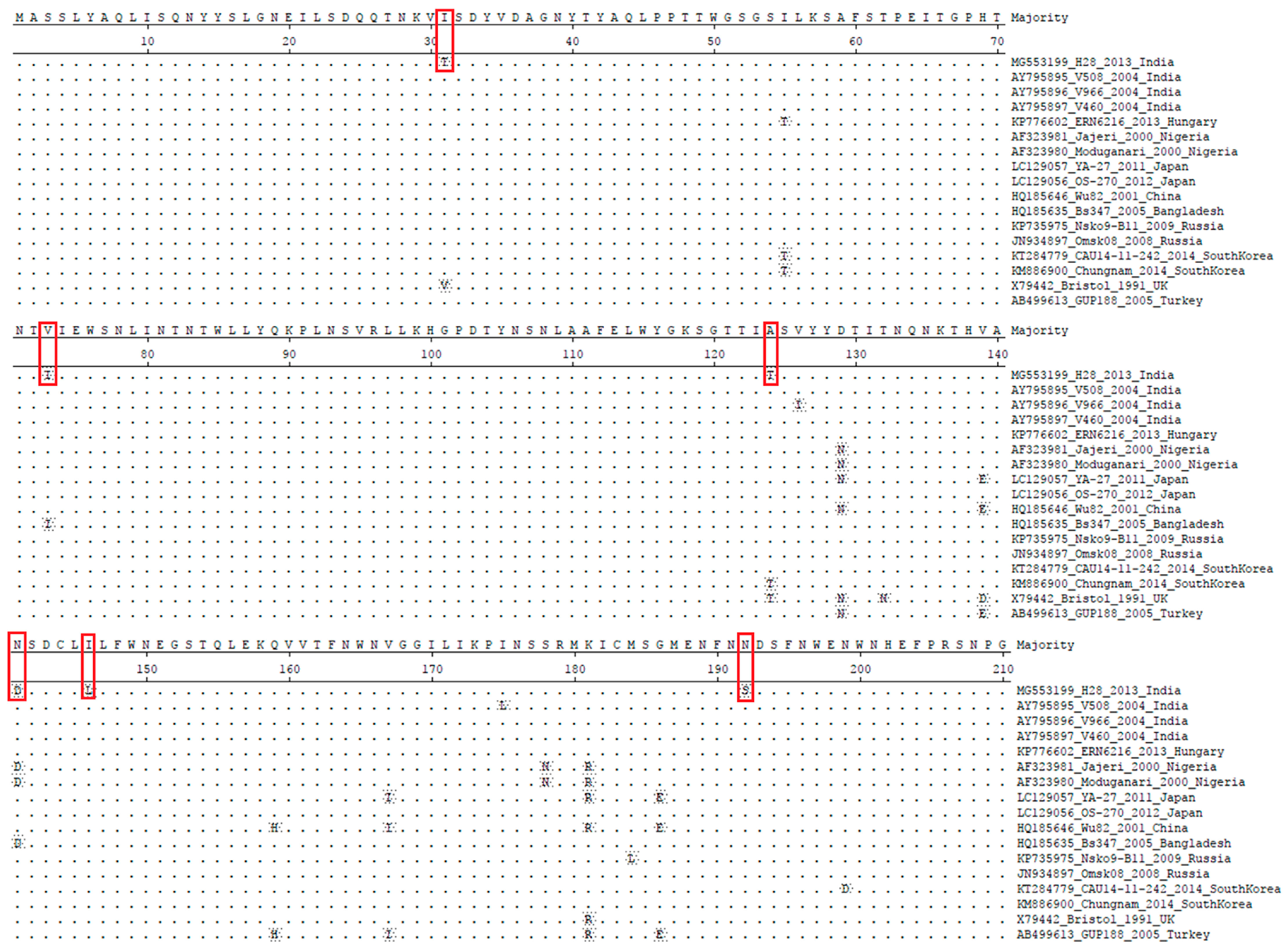
3.2.1. Structural Gene (VP6)
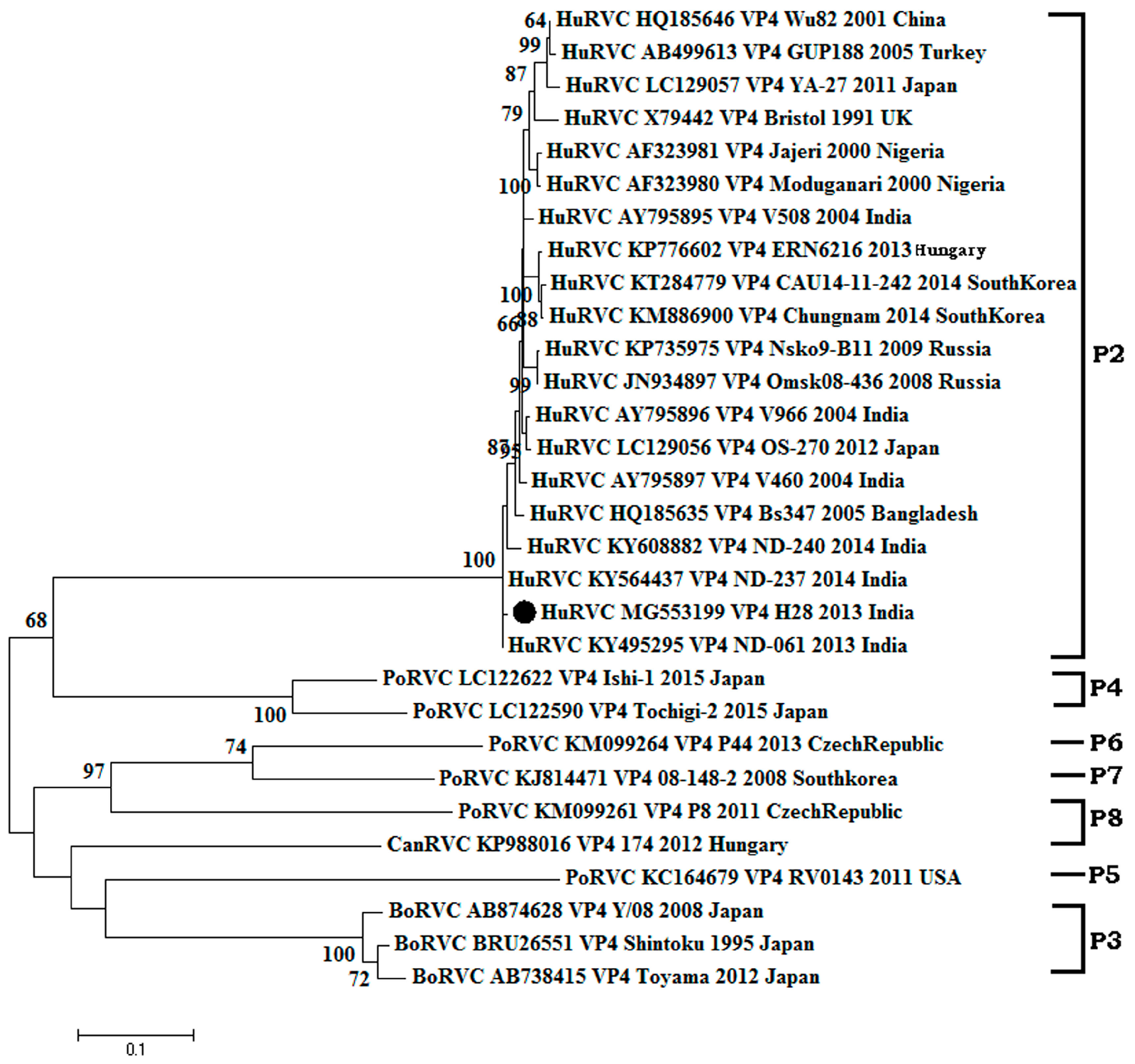
3.2.2. Structural Gene (VP4)
3.2.3. Non-Structural Gene NSP4
3.2.4. Structural Gene VP7
3.3. Prediction of N-Linked Glycosylation Pattern
4. Conclusions and Future Perspectives
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Joshi, M.S.; Jare, V.M.; Gopalkrishna, V. Group C rotavirus infection in patients with acute gastroenteritis in outbreaks in western India between 2006 and 2014. Epidemiol. Infect. 2017, 145, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Monavari, S.H.; Hadifar, S.; Mostafaei, S.; Miri, A.; Keshavarz, M.; Babaei, F.; Moghoofei, M. Epidemiology of rotavirus in the Iranian children: A systematic review and meta-analysis. J. Glob. Infect. Dis. 2017, 9, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Desselberger, U. Rotaviruses. Virus Res. 2014, 190, 75–96. [Google Scholar] [CrossRef] [PubMed]
- Sanaei Dashti, A.; Ghahremani, P.; Hashempoor, T.; Karimi, A. Molecular epidemiology of enteric adenovirus gastroenteritis in under-five-year-old children in Iran. Gastroenterol. Res. Pract. 2016, 2045697. [Google Scholar] [CrossRef] [PubMed]
- Hoa-Tran, T.N.; Nakagomi, O.; Dao, A.T.; Nguyen, A.T.; Agbemabiese, C.A.; Vu, H.M.; Nakagomi, T.; Thanh, N.T. Molecular epidemiology of noroviruses detected in Vietnamese children with acute gastroenteritis from 2012 to 2015. J. Med. Microbiol. 2017, 66, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Pinto, R.M.; Guix, S. Human astroviruses. Clin. Microbiol. Rev. 2014, 27, 1048–1074. [Google Scholar] [CrossRef] [PubMed]
- Estes, M.K.; Greenberg, H.B. Rotaviruses. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Cohen, J.I., Griffin, D.E., Lamb, R.A., Martin, M.A., Racaniello, V.R., Roizman, B., Eds.; Wolters Kluwer Health/Lippincott, Williams and Wilkins: Philadelphia, PA, USA; Buenos Aires, Argentina, 2013; pp. 1347–1401. [Google Scholar]
- Matthijnssens, J.; Otto, P.H.; Ciarlet, M.; Desselberger, U.; Van Ranst, M.; Johne, R. VP6-sequence-based cutoff values as a criterion for rotavirus species demarcation. Arch. Virol. 2012, 157, 1177–1182. [Google Scholar] [CrossRef] [PubMed]
- Ramig, F.; Ciarlet, M.; Mertens, P.P.C.; Dermody, T.S. Rotavirus. In Virus Taxonomy: Eight Report of the International Committee on Taxonomy of Viruses; Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005; pp. 484–496. [Google Scholar]
- Bányai, K.; Kemenesi, G.; Budinski, I.; Földes, F.; Zana, B.; Marton, S.; Varga-Kugler, R.; Oldal, M.; Kurucz, K.; Jakab, F. Candidate new rotavirus species in Schreiber’s bats, Serbia. Infect. Genet. Evol. 2017, 48, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, S.; Kobayashi, N.; Ishino, M.; Alam, M.M.; Ahmed, M.U.; Paul, S.K.; Ganesh, B.; Chawla-Sarkar, M.; Krishnan, T.; Naik, T.N.; et al. Whole genomic characterization of a human rotavirus strain B219 belonging to a novel group of the genus Rotavirus. J. Med. Virol. 2008, 80, 2023–2033. [Google Scholar] [CrossRef] [PubMed]
- Bohl, E.H.; Saif, L.J.; Theil, K.W.; Agnes, A.G.; Cross, R.F. Porcine pararotavirus: Detection, differentiation from rotavirus, and pathogenesis in gnotobiotic pigs. J. Clin. Microbiol. 1982, 15, 312–319. [Google Scholar] [PubMed]
- Dhama, K.; Saminathan, M.; Karthik, K.; Tiwari, R.; Shabbir, M.Z.; Kumar, N.; Malik, Y.S.; Singh, R.K. Avian rotavirus enteritis—An updated review. Vet. Q. 2015, 35, 142–158. [Google Scholar] [CrossRef] [PubMed]
- Kattoor, J.J.; Saurabh, S.; Malik, Y.S.; Sircar, S.; Dhama, K.; Ghosh, S.; Bányai, K.; Kobayashi, N.; Singh, R.K. Unexpected detection of porcine rotavirus C strains carrying human origin VP6 gene. Vet. Q. 2017, 37, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Kattoor, J.J.; Malik, Y.S.; Sharma, K.; Kumar, N.; Batra, M.; Jindal, N.; Yadav, A.S. Molecular evidence of group D rotavirus in commercial broiler chicks in India. Avian Biol. Res. 2013, 6, 313–316. [Google Scholar] [CrossRef]
- Malik, Y.P.S.; Chandrashekar, K.M.; Sharma, K.; Prasad, M.; Prasad, G. Evidence for Occurrence of Human group B rotavirus in Central India Based on Characterization of NSP2 Gene. Indian J. Virol. 2011, 22, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Deol, P.; Kattoor, J.J.; Sircar, S.; Ghosh, S.; Bányai, K.; Dhama, K.; Malik, Y.S. Avian Group D Rotaviruses: Structure, Epidemiology, Diagnosis, and Perspectives on Future Research Challenges. Pathogens 2017, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Mihalov-Kovács, E.; Gellért, Á.; Marton, S.; Farkas, S.L.; Fehér, E.; Oldal, M.; Jakab, F.; Martella, V.; Bányai, K. Candidate new rotavirus species in sheltered dogs, Hungary. Emerg. Infect. Dis. 2015, 21, 660–663. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.G.; Leutenegger, C.M.; Chan, R.; Delwart, E. Rotavirus I in feces of a cat with diarrhea. Virus Genes 2017, 53, 487–490. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Fauquier, A.; Roman, E.; Colomina, J.; Wilhelmi, I.; Glass, R.I.; Jiang, B. First detection of group C rotavirus in children with acute diarrhea in Spain. Arch. Virol. 2003, 148, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Papp, H.; László, B.; Jakab, F.; Ganesh, B.; De Grazia, S.; Matthijnssens, J.; Ciarlet, M.; Martella, V.; Bányai, K. Review of group A rotavirus strains reported in swine and cattle. Vet. Microbiol. 2013, 165, 190–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusumakar, A.L.; Malik, Y.S.; Prasad, G. Genomic diversity among group A rotaviruses from diarrheic children, piglets, buffalo and cow calves of Madhya Pradesh. Indian J. Microbiol. 2010, 50, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.; Svenungsson, B.; Hedlund, K.O.; Uhnoo, I.; Lagergren, A.; Akre, T.; Svensson, L. Incidence and genetic diversity of group C rotavirus among adults. J. Infect. Dis. 2000, 182, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Saif, L.J.; Bohl, E.H.; Theil, K.W.; Cross, R.F.; House, J.A. Rotavirus-like, calicivirus-like, and 23-nm virus-like particles associated with diarrhea in young pigs. J. Clin. Microbiol. 1980, 12, 105–111. [Google Scholar] [PubMed]
- Rodger, S.M.; Bishop, R.F.; Holmes, I.H. Detection of a rotavirus-like agent associated with diarrhea in an infant. J. Clin. Microbiol. 1982, 16, 724–726. [Google Scholar] [PubMed]
- Mawatari, T.; Taneichi, A.; Kawagoe, T.; Hosokawa, M.; Togashi, K.; Tsunemitsu, H. Detection of a bovine group C rotavirus from adult cows with diarrhea and reduced milk production. J. Vet. Med. Sci. 2004, 66, 887–890. [Google Scholar] [CrossRef] [PubMed]
- Wise, A.G.; Smedley, R.C.; Kiupel, M.; Maes, R.K. Detection of Group C Rotavirus in Juvenile Ferrets (Mustela putorius furo) with Diarrhea by Reverse Transcription Polymerase Chain Reaction: Sequencing and Analysis of the Complete Coding Region of the VP6 Gene. Vet. Pathol. 2009, 46, 985–991. [Google Scholar] [CrossRef] [PubMed]
- Marton, S.; Mihalov-Kovács, E.; Dóró, R.; Csata, T.; Fehér, E.; Oldal, M.; Jakab, F.; Matthijnssens, J.; Martella, V.; Bányai, K. Canine rotavirus C strain detected in Hungary shows marked genotype diversity. J. Gen. Virol. 2015, 96, 3059–3071. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Ciarlet, M.; Heiman, E.; Arijs, I.; Delbeke, T.; McDonald, S.M.; Palombo, E.A.; Iturriza-Gómara, M.; Maes, P.; Patton, J.T.; et al. Full genome-based classification of rotaviruses reveals a common origin between human Wa-Like and porcine rotavirus strains and human DS-1-like and bovine rotavirus strains. J. Virol. 2008, 82, 3204–3219. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Ciarlet, M.; Rahman, M.; Attoui, H.; Bányai, K.; Estes, M.K.; Gentsch, J.R.; Iturriza-Gómara, M.; Kirkwood, C.D.; Martella, V.; et al. Recommendations for the classification of group A rotaviruses using all 11 genomic RNA segments. Arch. Virol. 2008, 153, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Maes, P.; Matthijnssens, J.; Rahman, M.; Van Ranst, M. RotaC: A web-based tool for the complete genome classification of group A rotaviruses. BMC Microbiol. 2009, 9, 238. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.-J.; Matthijnssens, J.; Kim, D.-S.; Kim, J.-Y.; Alfajaro, M.M.; Park, J.-G.; Hosmillo, M.; Son, K.-Y.; Soliman, M.; Baek, Y.-B.; et al. Genetic diversity of the VP7, VP4 and VP6 genes of Korean porcine group C rotaviruses. Vet. Microbiol. 2015, 176, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Matthijnssens, J.; Ciarlet, M.; McDonald, S.M.; Attoui, H.; Bányai, K.; Brister, J.R.; Buesa, J.; Esona, M.D.; Estes, M.K.; Gentsch, J.R.; et al. Uniformity of rotavirus strain nomenclature proposed by the Rotavirus Classification Working Group (RCWG). Arch. Virol. 2011, 156, 1397–1413. [Google Scholar] [CrossRef] [PubMed]
- Marthaler, D.; Rossow, K.; Culhane, M.; Collins, J.; Goyal, S.; Ciarlet, M.; Matthijnssens, J. Identification, phylogenetic analysis and classification of porcine group C rotavirus VP7 sequences from the United States and Canada. Virology 2013, 446, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, D.; Ghosh, S.; Kuzuya, M.; Wang, Y.-H.; Zhou, X.; Chawla-Sarkar, M.; Paul, S.K.; Ishino, M.; Kobayashi, N. Whole-genome characterization of human group C rotaviruses: Identification of two lineages in the VP3 gene. J. Gen. Virol. 2011, 92, 361–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oishi, I.; Yamazaki, K.; Minekawa, Y. An occurrence of diarrheal cases associated with group C rotavirus in adults. Microbiol. Immunol. 1993, 37, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Tiku, V.R.; Jiang, B.; Kumar, P.; Aneja, S.; Bagga, A.; Bhan, M.K.; Ray, P. First study conducted in Northern India that identifies group C rotavirus as the etiological agent of severe diarrhea in children in Delhi. Virol. J. 2017, 14, 100. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Dennehy, P.H.; Spangenberger, S.; Gentsch, J.R.; Glass, R.I. First detection of group C rotavirus in fecal specimens of children with diarrhea in the United States. J. Infect. Dis. 1995, 172, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.W.; Mathan, M.M.; Mathew, M.; Martin, R.; Beards, G.M.; Mathan, V.I. Rotavirus epidemiology in Vellore, south India: Group, subgroup, serotype, and electrophoretype. J. Clin. Microbiol. 1988, 26, 2410–2414. [Google Scholar] [PubMed]
- Mukhopadhya, I.; Anbu, D.; Iturriza-Gomara, M.; Gray, J.J.; Brown, D.W.; Kavanagh, O.; Estes, M.K.; Kang, G. Anti-VP6 IgG antibodies against group A and group C rotaviruses in South India. Epidemiol. Infect. 2010, 138, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Malik, Y.P.S.; Rawat, V.; Sharma, K.; Vaid, N.; Kumar, N.; Sharma, A.K.; Rakholia, R.; Ganesh, B. Molecular evidences of mixed viral infection in an infant hospitalized with acute gastroenteritis in northern India. J. Pure Appl. Microbiol. 2014, 8, 733–737. [Google Scholar]
- Gabbay, Y.B.; Borges, A.A.; Oliveira, D.S.; Linhares, A.C.; Mascarenhas, J.D.P.; Barardi, C.R.M.; Simões, C.M.O.; Wang, Y.; Glass, R.I.; Jiang, B. Evidence for zoonotic transmission of group C rotaviruses among children in Belém, Brazil. J. Med. Virol. 2008, 80, 1666–1674. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Banik, S.; Faruque, A.S.G.; Taniguchi, K.; Sack, D.A.; Van Ranst, M.; Azim, T. Detection and characterization of human group C rotaviruses in Bangladesh. J. Clin. Microbiol. 2005, 43, 4460–4465. [Google Scholar] [CrossRef] [PubMed]
- Zhirakovskaia, E.; Tikunov, A.; Klemesheva, V.; Loginovskikh, N.; Netesov, S.; Tikunova, N. First genetic characterization of rotavirus C in Russia. Infect. Genet. Evol. 2016, 39, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rasool, N.B.; Hamzah, M.; Jegathesan, M.; Wong, Y.H.; Qian, Y.; Green, K.Y. Identification of a human group C rotavirus in Malaysia. J. Med. Virol. 1994, 43, 209–211. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.; Humphrey, C.D.; Kim, J.S.; Baek, L.J.; Song, J.-W.; Song, K.-J.; Jiang, B. First detection of group C rotavirus in children with acute gastroenteritis in South Korea. Clin. Microbiol. Infect. 2011, 17, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Peñaranda, M.E.; Cubitt, W.D.; Sinarachatanant, P.; Taylor, D.N.; Likanonsakul, S.; Saif, L.; Glass, R.I. Group C rotavirus infections in patients with diarrhea in Thailand, Nepal, and England. J. Infect. Dis. 1989, 160, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Mwenda, J.M.; Peenze, I.; Omollo, E.; Galo, M.; Steele, A.D. Human group C rotaviruses identified in Kenya. East Afr. Med. J. 2003, 80, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Malik, Y.S.; Kumar, N.; Sharma, K.; Saurabh, S.; Dhama, K.; Prasad, M.; Ghosh, S.; Bányai, K.; Kobayashi, N.; Singh, R.K. Multispecies reassortant bovine rotavirus strain carries a novel simian G3-like VP7 genotype. Infect. Genet. Evol. 2016, 41, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Huang, P.W.; Zhong, W.M.; Farkas, T.; Cubitt, D.W.; Matson, D.O. Designand evaluation of a primer pair that detects both Norwalk- and Sapporo-like caliciviruses by RT-PCR. J. Virol. Methods 1999, 83, 145–154. [Google Scholar] [CrossRef]
- Finkbeiner, S.R.; Holtz, L.R.; Jiang, Y.; Rajendran, P.; Franz, C.J.; Zhao, G.; Kang, G.; Wang, D. Human stool contains a previously unrecognized diversity of novel astroviruses. Virol. J. 2009, 6, 161. [Google Scholar] [CrossRef] [PubMed]
- Rosen, B.I.; Fang, Z.Y.; Glass, R.I.; Monroe, S.S. Cloning of human picobirnavirus genomic segments and development of an RT-PCR detection assay. Virology 2000, 277, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Cooke, S.J.; Clarke, I.N.; Freitas, R.B.; Gabbay, Y.B.; Lambden, P.R. The correct sequence of the porcine group C/Cowden rotavirus major inner capsid protein shows close homology with human isolates from Brazil and the U.K. Virology 1992, 190, 531–537. [Google Scholar] [CrossRef]
- Law, A.H.; Lee, D.C.; Cheung, B.K.; Yim, H.C.; Lau, A.S. Role for nonstructural protein 1 of severe acute respiratory syndrome coronavirus in chemokine dysregulation. J. Virol. 2007, 81, 416–422. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information (NCBI)[Internet]. Bethesda (MD): National Library of Medicine (US), National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 20 November 2017).
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Brunak, S. Prediction of glycosylation across the human proteome and the correlation to protein function. In Proceedings of the Pacific Symposium, Kauai, HI, USA, 3–7 January 2002. [Google Scholar]
- Iturriza-Gómara, M.; Kang, G.; Gray, J. Rotavirus genotyping: Keeping up with an evolving population of human rotaviruses. J. Clin. Virol. 2004, 31, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Malik, Y.S.; Sharma, K.; Vaid, N.; Batra, M.; Prasad, M.; Prasad, G.; Gulati, B.R.; Bhilegaonkar, K.N. Designing and validation of new primers for efficient genotyping of animal G3 rotaviruses. Vet. Arh. 2013, 83, 611–622. [Google Scholar]
- Barman, P.; Naik, T.N. Human rotavirus C strain V508 major inner capsid protein VP6 (VP6) gene, complete cds. Available online: https://www.ncbi.nlm.nih.gov/nuccore/AY795898 (accessed on 15 November 2017).
- Srivastava, S.; Jain, A. Rotavirus Nonstructural Protein 4 (NSP4)-Viral Enterotoxin with Multiple roles in Pathogenesis of Diarrhoea in Children. J. Appl. Pharm. Sci. 2015, 5, 146–153. [Google Scholar] [CrossRef]
- Soma, J.; Tsunemitsu, H.; Miyamoto, T.; Suzuki, G.; Sasaki, T.; Suzuki, T. Whole-genome analysis of two bovine rotavirus C strains:Shintoku and Toyama. J. Gen. Virol. 2013, 94, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Luchs, A.; do Carmo Sampaio Tavares Timenetsky, M. Phylogenetic analysis of human group C rotavirus circulating in Brazil reveals a potential unique NSP4 genetic variant and high similarity with Asian strains. Mol. Genet. Genom. 2015, 290, 969–986. [Google Scholar] [CrossRef] [PubMed]
- Meleg, E.; Banyai, K.; Martella, V.; Jiang, B.; Kocsis, B.; Kisfali, P.; Melegh, B.; Szucs, G. Detection and Quantification of Group C Rotaviruses in Communal Sewage. Appl. Environ. Microbiol. 2008, 74, 3394–3399. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Hasebe, A.; Miyazaki, A.; Tsunemitsu, H. Phylogenetic characterization of VP6 gene (inner capsid) of porcine rotavirus C collected in Japan. Infect. Genet. Evol. 2014, 26, 223–227. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Human Strains | Porcine | Bovine | Canine | Ferret | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gene | Nucleotide % | Amino Acid % | Nucleotide % | Amino Acid % | Nucleotide % | Amino Acid % | Nucleotide % | Amino Acid % | Nucleotide % | Amino Acid % |
| VP6 | 95.5–98.6 | 98.3–99.1 | 73.6–99.9 | 84.3–99.1 | 78.1–78.5 | 88.7 | 64 | 90 | 64.3 | 88.3 |
| VP4 | 94.3–97 | 95.6–97.6 | 59.8–65.1 | 60.4–65.8 | 61.7–62.6 | 67.5–68.7 | 63.8 | 68.2 | - | - |
| NSP4 | 91.4–96 | 90.2–95.4 | 58.9–61.3 | 59.5–62.1 | 55.2–58.5 | 54.9–56.9 | 60.4 | 59.5 | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhat, S.; Kattoor, J.J.; Malik, Y.S.; Sircar, S.; Deol, P.; Rawat, V.; Rakholia, R.; Ghosh, S.; Vlasova, A.N.; Nadia, T.; et al. Species C Rotaviruses in Children with Diarrhea in India, 2010–2013: A Potentially Neglected Cause of Acute Gastroenteritis. Pathogens 2018, 7, 23. https://doi.org/10.3390/pathogens7010023
Bhat S, Kattoor JJ, Malik YS, Sircar S, Deol P, Rawat V, Rakholia R, Ghosh S, Vlasova AN, Nadia T, et al. Species C Rotaviruses in Children with Diarrhea in India, 2010–2013: A Potentially Neglected Cause of Acute Gastroenteritis. Pathogens. 2018; 7(1):23. https://doi.org/10.3390/pathogens7010023
Chicago/Turabian StyleBhat, Sudipta, Jobin Jose Kattoor, Yashpal Singh Malik, Shubhankar Sircar, Pallavi Deol, Vinita Rawat, Ritu Rakholia, Souvik Ghosh, Anastasia N. Vlasova, Touil Nadia, and et al. 2018. "Species C Rotaviruses in Children with Diarrhea in India, 2010–2013: A Potentially Neglected Cause of Acute Gastroenteritis" Pathogens 7, no. 1: 23. https://doi.org/10.3390/pathogens7010023
APA StyleBhat, S., Kattoor, J. J., Malik, Y. S., Sircar, S., Deol, P., Rawat, V., Rakholia, R., Ghosh, S., Vlasova, A. N., Nadia, T., Dhama, K., & Kobayashi, N. (2018). Species C Rotaviruses in Children with Diarrhea in India, 2010–2013: A Potentially Neglected Cause of Acute Gastroenteritis. Pathogens, 7(1), 23. https://doi.org/10.3390/pathogens7010023