Soil Chemical Properties Barely Perturb the Abundance of Entomopathogenic Fusarium oxysporum: A Case Study Using a Generalized Linear Mixed Model for Microbial Pathogen Occurrence Count Data

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Infection Frequencies
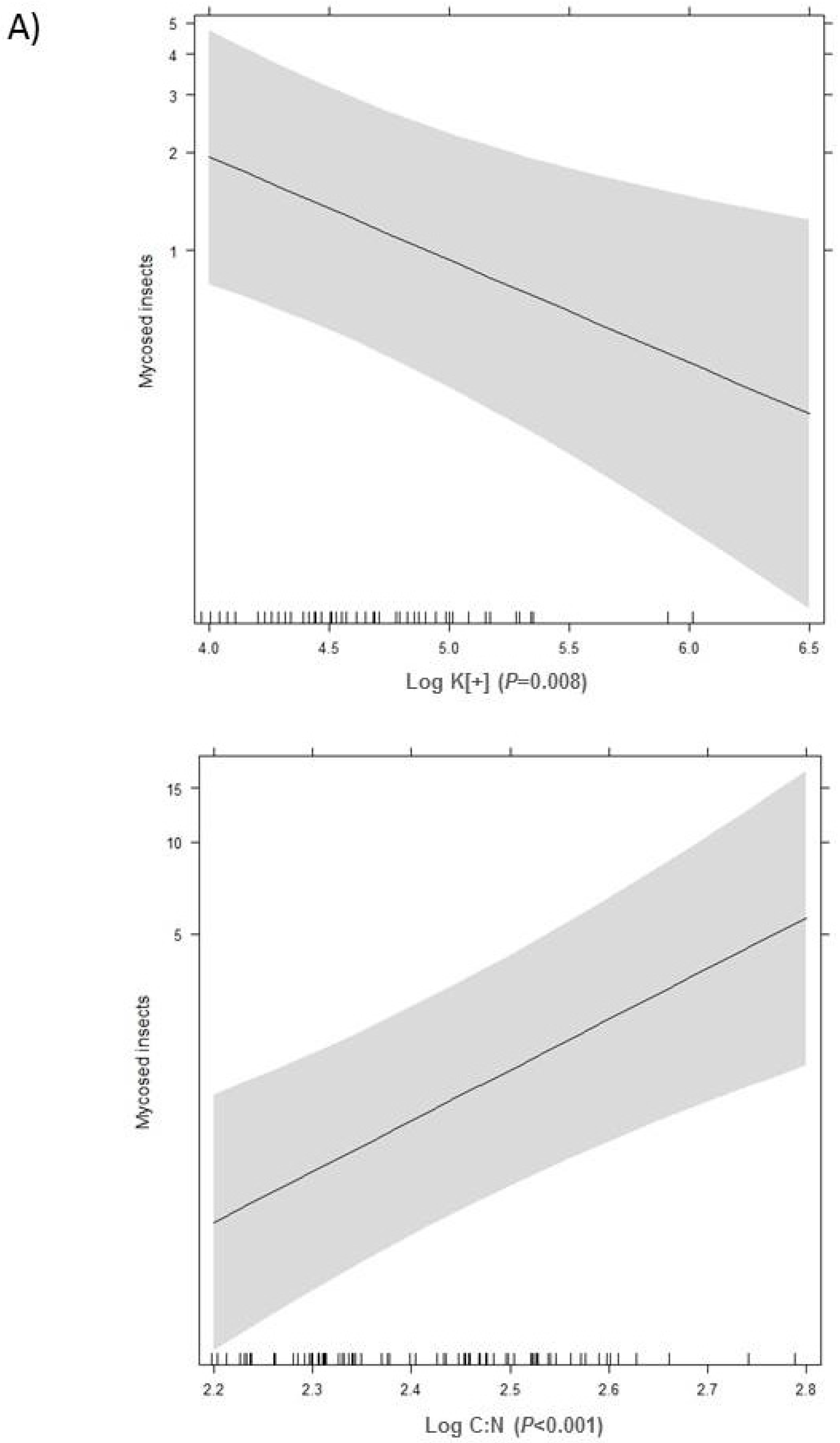
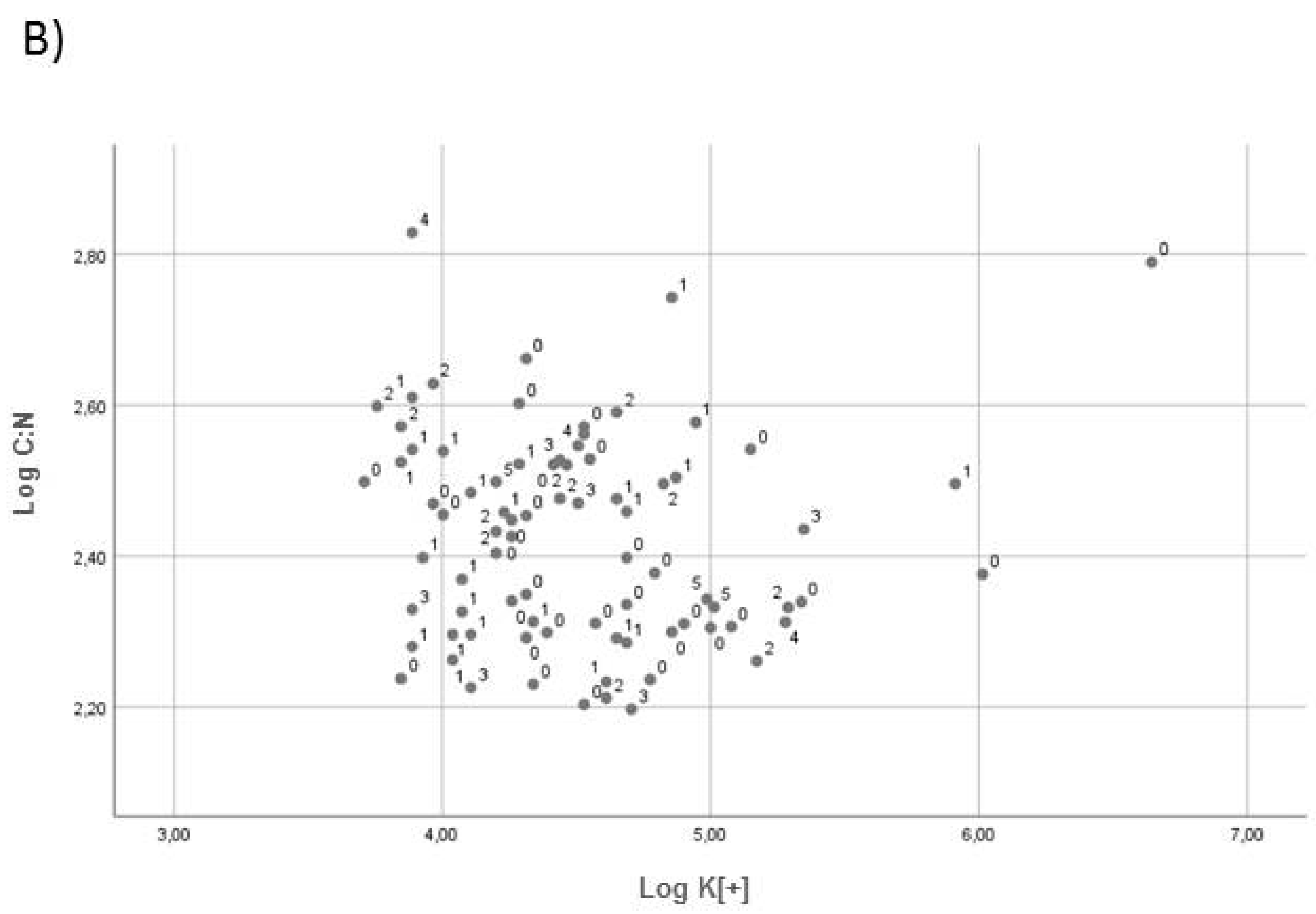
2.2. Effect of Soil Chemical Properties on Fungal Abundance
3. Discussion
4. Materials and Methods
4.1. Sampling Site
4.2. Fungal Isolation, Identification, and Screening
4.3. Soil Analyses and Calculations
4.4. Data Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sharma, L.; Oliveira, I.; Torres, L.; Marques, G. Entomopathogenic fungi in Portuguese vineyards soils: Suggesting a ‘Galleria-Tenebrio-bait method’ as bait-insects Galleria and Tenebrio significantly underestimate the respective recoveries of Metarhizium (robertsii) and Beauveria (bassiana). MycoKeys 2018, 38, 1–23. [Google Scholar] [CrossRef]
- Sharma, L.; Gonçalves, F.; Oliveira, I.; Torres, L.; Marques, G. Insect-associated fungi from naturally mycosed vine mealybug Planococcus ficus (Signoret) (Hemiptera: Pseudococcidae). Biocontrol Sci. Technol. 2018, 28, 122–141. [Google Scholar] [CrossRef]
- Tiago, P.V.; Medeiros, L.V.; Carneiro Leão, M.P.; Santos, A.C.; da Costa, A.F.; de Oliveira, N.T. Polymorphisms in entomopathogenic fusaria based on inter simple sequence repeats. Biocontrol Sci. Technol. 2016, 26, 1401–1410. [Google Scholar] [CrossRef]
- O’Donnell, K.; Humber, R.A.; Geiser, D.M.; Kang, S.; Park, B.; Robert, V.A.R.G.; Crous, P.W.; Johnston, P.R.; Aoki, T.; Rooney, A.P.; et al. Phylogenetic diversity of insecticolous fusaria inferred from multilocus DNA sequence data and their molecular identification via Fusarium-ID and Fusarium MLST. Mycologia 2012, 104, 427–445. [Google Scholar] [CrossRef] [PubMed]
- Teetor-Barsch, G.H.; Roberts, D.W. Entomogenous Fusarium species. Mycopathologia 1983, 84, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Velasco, G.Y.; Prados-Rosales, R.C.; Ortíz-Urquiza, A.; Quesada-Moraga, E.; Di Pietro, A. Galleria mellonella as model host for the trans-kingdom pathogen Fusarium oxysporum. Fungal Genet. Biol. 2011, 48, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Jaronski, S.T. Soil ecology of the entomopathogenic ascomycetes: A critical examination of what we (think) we know. In Use of Entomopathogenic Fungi in Biological Pest Management; Maniana, K., Ekesi, S., Eds.; Research SignPosts: Trivandrum, India, 2007; pp. 91–144. ISBN 978-8-13-080192-6. [Google Scholar]
- Quesada-Moraga, E.; Navas-Cortés, J.A.; Maranhao, E.A.A.; Ortiz-Urquiza, A.; Santiago-Álvarez, C. Factors affecting the occurrence and distribution of entomopathogenic fungi in natural and cultivated soils. Mycol. Res. 2007, 111, 947–966. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, R.; Barbercheck, M.E. Soil management effects on entomopathogenic fungi during the transition to organic agriculture in a feed grain rotation. Biol. Control 2009, 51, 435–443. [Google Scholar] [CrossRef]
- Oddsdottir, E.S.; Nielsen, C.; Sen, R.; Harding, S.; Eilenberg, J.; Halldorsson, G. Distribution patterns of soil entomopathogenic and birch symbiotic ectomycorrhizal fungi across native woodlandand degraded habitats in Iceland. Icel. Agric. Sci. 2010, 23, 37–49. [Google Scholar]
- Garrido-Jurado, I.; Torrent, J.; Barrón, V.; Corpas, A.; Quesada-Moraga, E. Soil properties affect the availability, movement, and virulence of entomopathogenic fungi conidia against puparia of Ceratitis capitata (Diptera: Tephritidae). Biol. Control 2011, 58, 277–285. [Google Scholar] [CrossRef]
- Clifton, E.H.; Jaronski, S.T.; Hodgson, E.W.; Gassmann, A.J. Abundance of soil-borne entomopathogenic fungi in organic and conventional fields in the midwestern USA with an emphasis on the effect of herbicides and fungicides on fungal persistence. PLoS ONE 2015, 10, e0133613. [Google Scholar] [CrossRef] [PubMed]
- Carlos, C.; Gonçalves, F.; Oliveira, I.; Torres, L. Is a biofix necessary for predicting the flight phenology of Lobesia botrana in Douro demarcated region vineyards? Crop Prot. 2018, 110, 57–64. [Google Scholar] [CrossRef]
- Garrido-Jurado, I.; Valverde-García, P.; Quesada-Moraga, E. Use of a multiple logistic regression model to determine the effects of soil moisture and temperature on the virulence of entomopathogenic fungi against pre-imaginal Mediterranean fruit fly Ceratitis capitata. Biol. Control 2011, 59, 366–372. [Google Scholar] [CrossRef]
- Bolker, B.M.; Brooks, M.E.; Clark, C.J.; Geange, S.W.; Poulsen, J.R.; Stevens, M.H.H.; White, J.-S.S. Generalized linear mixed models: A practical guide for ecology and evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, T.; Moral, R.; Hinde, J.; Demétrio, C.; Zocchi, S.S.; Zanardo, A.B.; Delalibera, I. Generalized linear mixed models applied to overdispersed proportion data in a fungal occurrence study. In Proceedings of the 30th International Workshop on Statistical Modelling, Linz, Austria, 6–10 July 2015; Friedl, H., Wagner, H., Eds.; Johannes Kepler Universität: Linz, Austria, 2015; Volume 2. [Google Scholar]
- Barrios, E. Soil biota, ecosystem services and land productivity. Ecol. Econ. 2007, 64, 269–285. [Google Scholar] [CrossRef]
- Birkhofer, K.; Bezemer, T.M.; Bloem, J.; Bonkowski, M.; Christensen, S.; Dubois, D.; Ekelund, F.; Fließbach, A.; Gunst, L.; Hedlund, K.; et al. Long-term organic farming fosters below and aboveground biota: Implications for soil quality, biological control and productivity. Soil Biol. Biochem. 2008, 40, 2297–2308. [Google Scholar] [CrossRef]
- Carlos, C.G.F.; Sousa, S.; Salvação, J.; Sharma, L.; Soares, R.; Manso, J.; Nóbrega, M.; Lopes, A.; Soares, S.; Aranha, J.; et al. Environmentally safe strategies to control the European grapevine moth, Lobesia botrana (Den. & Schiff.) in the Douro demarcated region. Cienc. Tec. Vitivinic. 2013, 1006–1011. [Google Scholar]
- Gonçalves, F.; Carlos, C.; Aranha, J.; Torres, L. Does habitat heterogeneity affect the diversity of epigaeic arthropods in vineyards? Agric. For. Entomol. 2018, 20, 366–379. [Google Scholar] [CrossRef]
- Bednarek, A.; Gaugler, R. Compatibility of soil amendments with entomopathogenic nematodes. J. Nematol. 1997, 29, 220–227. [Google Scholar] [PubMed]
- Sun, B.-D.; Liu, X.-Z. Occurrence and diversity of insect-associated fungi in natural soils in China. Appl. Soil Ecol. 2008, 39, 100–108. [Google Scholar] [CrossRef]
- Goble, T.A.; Dames, J.F.; Hill, M.P.; Moore, S.D. The effects of farming system, habitat type and bait type on the isolation of entomopathogenic fungi from citrus soils in the eastern Cape province, South Africa. BioControl 2010, 55, 399–412. [Google Scholar] [CrossRef]
- Bremner, J.M. Nitrogen-total. In Methods of Soil Analysis Part 3—Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Eds.; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Bower, C.A.; Reitemeier, R.F.; Fireman, M. Exchangeable cation analysis of saline and alkali soils. Soil Sci. 1952, 73, 251–262. [Google Scholar] [CrossRef]
- Thomas, G.W. Soil pH and soil acidity. In Methods of Soil Analysis Part 3—Chemical Methods; Sparks, D.L., Page, A.L., Helmke, P.A., Loeppert, R.H., Eds.; Soil Science Society of America, American Society of Agronomy: Madison, WI, USA, 1996; pp. 475–490. [Google Scholar]
- Crawley, M.J. Generalized linear models. In The R book; Crawley, M.J., Ed.; John Wiley & Sons: Chichester, UK, 2007; pp. 511–526. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 978-0-38-721706-2. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fertil. Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Farm Type | Soil Type | Herbicide Usage | Rapid Texture | Collection Site | OM | P | K | Ca2+ | Mg2+ | K+ | Na+ | ECEC | N | TA | pH | C:N | DBS | Fusarium oxysporum Count |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Carvalhas | Vineyards | 0 | Medium | nh1mCa1A | 6.31 | 22 | 139 | 7.94 | 1.55 | 0.68 | 0.10 | 10.36 | 3.01 | 0.10 | 5.9 | 12.155174 | 99 | 1 |
| Carvalhas | Vineyards | 0 | Medium | nh1mCa2C | 4.57 | 6 | 90 | 4.27 | 1.20 | 0.33 | 0.03 | 5.83 | 2.45 | 0.00 | 6.3 | 10.819543 | 100 | 3 |
| Carvalhas | Vineyards | 1 | Medium | hmCa3B | 2.21 | 221 | 197 | 3.89 | 0.69 | 0.63 | 0.01 | 5.78 | 1.38 | 0.55 | 5.2 | 9.2963577 | 90 | 2 |
| Carvalhas | Vineyards | 1 | Medium | hmCa4A | 1.43 | 46 | 60 | 3.47 | 1.76 | 0.26 | 0.08 | 6.55 | 0.93 | 0.98 | 5.5 | 8.934769 | 85 | 1 |
| Carvalhas | Vineyards | 1 | Medium | hmCa5A | 1.39 | 45 | 56 | 4.94 | 1.17 | 0.19 | 0.18 | 6.71 | 0.90 | 0.23 | 5.8 | 8.9326187 | 97 | 1 |
| Carvalhas | Vineyards | 1 | Gross | hgCa6B | 4.15 | 96 | 84 | 1.09 | 0.37 | 0.26 | 0.03 | 4.15 | 2.21 | 2.40 | 4.4 | 10.895056 | 42 | 2 |
| Carvalhas | Vineyards | 1 | Medium | hmCa7 | 1.24 | 91 | 92 | 4.10 | 1.07 | 0.28 | 0.03 | 6.22 | 0.89 | 0.75 | 5.3 | 8.0555353 | 88 | 0 |
| Carvalhas | Vineyards | 1 | Medium | hmCa8 | 3.17 | 81 | 147 | 5.06 | 0.91 | 0.48 | 0.06 | 6.99 | 2.04 | 0.50 | 5.2 | 9.0237878 | 93 | 0 |
| Carvalhas | Vineyards | 1 | Medium | hmCa9 | 1.76 | 106 | 118 | 3.33 | 0.67 | 0.40 | 0.02 | 5.72 | 1.22 | 1.30 | 4.9 | 8.3599869 | 77 | 0 |
| Carvalhas | Vineyards | 1 | Medium | hmCa10 | 2.78 | 56 | 74 | 3.68 | 1.24 | 0.27 | 0.06 | 6.22 | 1.81 | 0.98 | 5.0 | 8.894316 | 84 | 0 |
| Carvalhas | Vineyards | 1 | Medium | hmCa11A | 2.57 | 70 | 108 | 3.50 | 0.80 | 0.41 | 0.10 | 6.36 | 1.69 | 1.55 | 4.6 | 8.8276961 | 76 | 1 |
| Carvalhas | Vineyards | 1 | Medium | hmCa12A | 2.74 | 64 | 104 | 6.24 | 1.49 | 0.36 | 0.03 | 8.70 | 1.79 | 0.58 | 5.3 | 8.8875571 | 93 | 1 |
| Carvalhas | Hedgerows | 0 | Medium | nh2mCaM1B | 5.71 | 14 | 86 | 8.17 | 1.43 | 0.37 | 0.07 | 10.25 | 2.90 | 0.00 | 6.4 | 11.438218 | 100 | 2 |
| Carvalhas | Hedgerows | 0 | Medium | nh2mCaM2D | 5.74 | 15 | 90 | 8.14 | 1.60 | 0.39 | 0.06 | 10.19 | 2.83 | 0.00 | 6.2 | 11.758777 | 100 | 4 |
| São Luiz | Vineyards | 1 | Medium | hmSl11 | 4.38 | 174 | 82 | 2.41 | 1.87 | 0.30 | 0.10 | 4.93 | 2.22 | 0.25 | 5.5 | 11.44503 | 95 | 0 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl12 | 1.54 | 37 | 40 | 4.27 | 1.04 | 0.13 | 0.08 | 5.53 | 0.80 | 0.00 | 6.1 | 11.161607 | 100 | 0 |
| São Luiz | Vineyards | 1 | Medium | hmSl21 | 6.38 | 34 | 72 | 6.09 | 1.71 | 0.25 | 0.05 | 8.09 | 2.96 | 0.00 | 6.2 | 12.492244 | 100 | 0 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl22A | 3.41 | 28 | 68 | 3.78 | 0.83 | 0.21 | 0.03 | 4.85 | 1.85 | 0.00 | 6.3 | 10.680227 | 100 | 1 |
| São Luiz | Vineyards | 1 | Medium | hmSl3A | 2.09 | 56 | 50 | 8.10 | 1.65 | 0.14 | 0.08 | 9.96 | 1.21 | 0.00 | 6.8 | 9.9992 | 100 | 1 |
| São Luiz | Vineyards | 1 | Medium | hmSl41A | 1.26 | 26 | 58 | 3.46 | 1.09 | 0.14 | 0.06 | 4.74 | 0.79 | 0.00 | 5.9 | 9.2397671 | 100 | 1 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl42 | 2.93 | 43 | 108 | 3.81 | 1.17 | 0.32 | 0.06 | 5.35 | 1.82 | 0.00 | 6.1 | 9.3399121 | 100 | 0 |
| São Luiz | Vineyards | 1 | Medium | hmSl51 | 6.84 | 45 | 74 | 1.34 | 0.69 | 0.24 | 0.06 | 2.74 | 2.98 | 0.40 | 5.6 | 13.314371 | 85 | 0 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl52 | 3.77 | 25 | 70 | 3.37 | 0.40 | 0.21 | 0.06 | 4.04 | 2.12 | 0.00 | 6.3 | 10.310496 | 100 | 0 |
| São Luiz | Vineyards | 1 | Medium | hmSl61B | 4.04 | 28 | 46 | 3.12 | 2.61 | 0.18 | 0.17 | 6.14 | 1.94 | 0.05 | 5.8 | 12.086662 | 99 | 2 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl62B | 3.13 | 23 | 42 | 4.30 | 2.29 | 0.17 | 0.24 | 7.06 | 1.46 | 0.05 | 5.9 | 12.44421 | 99 | 2 |
| São Luiz | Vineyards | 1 | Medium | hmSl71A | 2.48 | 14 | 48 | 2.85 | 2.56 | 0.16 | 0.15 | 5.79 | 1.23 | 0.08 | 5.3 | 11.690122 | 99 | 1 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl72 | 2.07 | 6 | 52 | 2.45 | 2.56 | 0.15 | 0.13 | 5.36 | 1.11 | 0.08 | 5.5 | 10.809946 | 99 | 0 |
| São Luiz | Vineyards | 1 | Medium | hmSl81A | 1.59 | 38 | 76 | 2.58 | 1.36 | 0.19 | 0.06 | 4.18 | 1.01 | 0 | 6.4 | 9.1081822 | 100 | 1 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl82 | 1.90 | 53 | 108 | 1.76 | 0.85 | 0.23 | 0.12 | 2.96 | 1.1 | 0 | 6.0 | 9.9992 | 100 | 0 |
| São Luiz | Vineyards | 1 | Medium | hmSl91 | 1.29 | 42 | 70 | 1.44 | 0.43 | 0.15 | 0.03 | 2.18 | 0.71 | 0.13 | 5.9 | 10.562535 | 94 | 0 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl92 | 1.71 | 51 | 54 | 1.86 | 0.43 | 0.15 | 0.05 | 2.48 | 0.93 | 0 | 6.2 | 10.64431 | 100 | 0 |
| São Luiz | Vineyards | 1 | Medium | hmSl101 | 2.18 | 29 | 74 | 2.43 | 0.67 | 0.22 | 0.06 | 3.38 | 1.19 | 0 | 6.4 | 10.629402 | 100 | 0 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl102 | 4.02 | 20 | 92 | 3.76 | 0.77 | 0.25 | 0.03 | 4.82 | 1.93 | 0 | 6.3 | 12.081935 | 100 | 0 |
| São Luiz | Vineyards | 1 | Medium | hmSl111B | 2.31 | 10 | 66 | 7.18 | 1.39 | 0.15 | 0.07 | 8.79 | 1.29 | 0 | 6.6 | 10.386766 | 100 | 2 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl112A | 2.88 | 45 | 60 | 8.14 | 1.23 | 0.16 | 0.08 | 9.61 | 1.52 | 0 | 6.7 | 10.985963 | 100 | 1 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl112 | 4.22 | 7 | 70 | 11.23 | 1.36 | 0.16 | 0.15 | 12.91 | 2.61 | 0 | 6.8 | 9.3862222 | 100 | 0 |
| São Luiz | Vineyards | 1 | Medium | hmSl131B | 2.79 | 50 | 104 | 2.19 | 0.69 | 0.28 | 0.05 | 3.22 | 1.31 | 0 | 6.1 | 12.334891 | 100 | 2 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl132 | 2.13 | 49 | 94 | 3.33 | 0.59 | 0.26 | 0.03 | 4.21 | 1.07 | 0 | 6.3 | 11.531788 | 100 | 0 |
| São Luiz | Vineyards | 1 | Medium | hmSl141A | 2.98 | 59 | 129 | 1.78 | 0.24 | 0.32 | 0.03 | 2.37 | 1.54 | 0 | 5.6 | 11.232868 | 100 | 1 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl142A | 3.05 | 52 | 368 | 2.42 | 0.59 | 0.51 | 0.06 | 3.57 | 1.59 | 0 | 6.2 | 11.131185 | 100 | 1 |
| São Luiz | Vineyards | 1 | Gross | hgSl151A | 10.96 | 79 | 127 | 6.98 | 2.45 | 0.47 | 0.08 | 10.93 | 4.38 | 0.95 | 6.1 | 14.519386 | 91 | 1 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl152 | 1.57 | 8 | 74 | 1.71 | 0.77 | 0.20 | 0.07 | 2.76 | 0.96 | 0 | 5.6 | 9.4784083 | 100 | 0 |
| São Luiz | Vineyards | 1 | Gross | hgSl241 | 16.37 | 152 | 767 | 6.12 | 3.73 | 0.88 | 0.13 | 10.87 | 6.22 | 0 | 6.3 | 15.262445 | 100 | 0 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl242 | 5.16 | 51 | 171 | 5.39 | 1.36 | 0.43 | 0.03 | 7.22 | 2.56 | 0 | 6.3 | 11.694377 | 100 | 0 |
| São Luiz | Vineyards | 0 | Medium | nh1mSl26A | 2.17 | 64 | 54 | 7.52 | 2.45 | 0.12 | 0.10 | 10.54 | 1.08 | 0.35 | 7.0 | 11.665733 | 97 | 1 |
| São Luiz | Hedgerows | 0 | Gross | nh2gSlM1A | 9.02 | 4 | 48 | 3.12 | 1.36 | 0.14 | 0.10 | 4.72 | 4.15 | 0 | 5.4 | 12.601401 | 100 | 1 |
| São Luiz | Hedgerows | 0 | Gross | nh2gSlM2D | 12.84 | 5 | 48 | 3.89 | 1.49 | 0.16 | 0.86 | 6.40 | 4.68 | 0 | 6.7 | 15.91753 | 100 | 4 |
| São Luiz | Hedgerows | 0 | Gross | nh2gSlM3B | 8.55 | 2 | 52 | 3.18 | 1.39 | 0.29 | 0.14 | 5.50 | 3.86 | 0.50 | 6.0 | 12.848713 | 91 | 2 |
| São Luiz | Hedgerows | 0 | Gross | nh2gSlM4A | 6.79 | 1 | 46 | 1.82 | 0.83 | 0.16 | 0.08 | 3.27 | 3.43 | 0.38 | 5.9 | 11.485962 | 89 | 1 |
| Granja | Vineyards | 1 | Medium | hmGr21 | 1.41 | 48 | 408 | 3.5 | 1.0 | 0.9 | 0.4 | 5.87 | 0.84 | 0.09 | 5.8 | 9.7611238 | 98 | 0 |
| Granja | Vineyards | 0 | Medium | nh1mGr22E | 1.10 | 24 | 145 | 3.2 | 0.8 | 0.4 | 0.3 | 5.25 | 0.68 | 0.585 | 5.1 | 9.4110118 | 89 | 5 |
| Granja | Vineyards | 1 | Medium | hmGr31 | 1.19 | 6 | 133 | 2.4 | 1.0 | 0.3 | 0.3 | 4.30 | 0.76 | 0.315 | 4.6 | 9.0782211 | 93 | 0 |
| Granja | Vineyards | 0 | Medium | nh1mGr32C | 0.98 | 11 | 60 | 3.6 | 0.8 | 0.2 | 0.3 | 5.28 | 0.69 | 0.338 | 4.6 | 8.2602087 | 94 | 3 |
| Granja | Vineyards | 1 | Medium | hmGr41 | 2.07 | 22 | 207 | 1.7 | 0.7 | 0.7 | 0.4 | 4.29 | 1.28 | 0.72 | 4.7 | 9.37425 | 83 | 0 |
| Granja | Vineyards | 0 | Medium | nh1mGr42C | 2.57 | 32 | 209 | 1.7 | 0.7 | 0.7 | 0.3 | 3.90 | 1.43 | 0.563 | 5.0 | 10.418747 | 86 | 3 |
| Granja | Vineyards | 1 | Medium | hmGr51B | 1.05 | 30 | 175 | 1.6 | 0.5 | 0.4 | 0.3 | 3.48 | 0.71 | 0.63 | 5.0 | 8.590862 | 82 | 2 |
| Granja | Vineyards | 0 | Medium | nh1mGr52D | 1.40 | 25 | 195 | 1.4 | 0.4 | 0.6 | 0.3 | 3.32 | 0.89 | 0.608 | 5.0 | 9.1003955 | 82 | 4 |
| Granja | Vineyards | 1 | Medium | hmGr61 | 1.52 | 17 | 120 | 1.9 | 0.7 | 0.3 | 0.4 | 4.22 | 0.90 | 0.945 | 4.4 | 9.7769956 | 78 | 0 |
| Granja | Vineyards | 0 | Medium | nh1mGr62E | 1.38 | 11 | 149 | 1.4 | 0.4 | 0.4 | 0.3 | 3.83 | 0.86 | 1.35 | 4.4 | 9.3015814 | 65 | 5 |
| Granja | Vineyards | 1 | Medium | hmGr71A | 0.78 | 10 | 100 | 2.2 | 0.9 | 0.3 | 0.3 | 4.31 | 0.54 | 0.675 | 4.7 | 8.3326667 | 84 | 1 |
| Granja | Vineyards | 0 | Medium | nh1mGr72C | 0.55 | 7 | 110 | 1.3 | 0.5 | 0.3 | 0.3 | 3.18 | 0.40 | 0.72 | 4.8 | 7.99936 | 77 | 3 |
| Aciprestes | Vineyards | 1 | Medium | hmAc11 | 1.35 | 100 | 100 | 4.19 | 0.85 | 0.16 | 0.13 | 5.33 | 0.96 | 0 | 7.2 | 8.1347658 | 100 | 2 |
| Aciprestes | Vineyards | 0 | Medium | nh1mAc12 | 2.08 | 96 | 108 | 4.48 | 1.36 | 0.21 | 0.03 | 6.08 | 1.13 | 0 | 7.0 | 10.68941 | 100 | 1 |
| Aciprestes | Vineyards | 1 | Medium | hmAc21 | 2.32 | 90 | 84 | 6.11 | 1.07 | 0.15 | 0.06 | 7.38 | 1.17 | 0 | 7.5 | 11.511899 | 100 | 3 |
| Aciprestes | Vineyards | 0 | Medium | nh1mAc22 | 3.04 | 106 | 72 | 6.54 | 1.71 | 0.13 | 0.15 | 8.53 | 1.54 | 0 | 7.3 | 11.453629 | 100 | 1 |
| Aciprestes | Vineyards | 1 | Medium | hmAc3 | 2.55 | 214 | 104 | 4.67 | 0.96 | 0.20 | 0.04 | 5.87 | 1.36 | 0 | 7.2 | 10.888835 | 100 | 1 |
| Aciprestes | Vineyards | 1 | Medium | hmAc41 | 1.31 | 66 | 66 | 3.90 | 1.20 | 0.11 | 0.09 | 5.31 | 0.68 | 0 | 7.3 | 11.160872 | 100 | 5 |
| Aciprestes | Vineyards | 0 | Medium | nh1mAc42 | 1.24 | 83 | 46 | 6.4 | 1.1 | 0.1 | 0.4 | 7.96 | 0.86 | 0 | 6.9 | 8.3714233 | 100 | 0 |
| Aciprestes | Vineyards | 1 | Medium | hmAc51 | 0.51 | 17 | 127 | 3.94 | 1.55 | 0.09 | 0.11 | 5.68 | 0.33 | 0 | 7.3 | 8.9689794 | 100 | 0 |
| Aciprestes | Vineyards | 0 | Medium | nh1mAc52 | 0.88 | 28 | 48 | 4.8 | 1.3 | 0.2 | 0.3 | 6.67 | 0.55 | 0 | 6.7 | 9.2719855 | 100 | 3 |
| Aciprestes | Vineyards | 1 | Medium | hmAc61 | 0.99 | 91 | 76 | 5.84 | 1.44 | 0.12 | 0.09 | 7.49 | 0.69 | 0 | 7.0 | 8.3036835 | 100 | 0 |
| Aciprestes | Vineyards | 0 | Medium | nh1mAc61 | 2.16 | 100 | 58 | 6.9 | 1.9 | 0.2 | 0.5 | 9.49 | 1.29 | 0 | 6.7 | 9.6891473 | 100 | 1 |
| Aciprestes | Vineyards | 1 | Medium | hmAc71 | 1.46 | 59 | 96 | 4.78 | 1.44 | 0.11 | 0.06 | 6.40 | 0.93 | 0 | 7.2 | 9.0852946 | 100 | 0 |
| Aciprestes | Vineyards | 0 | Medium | nh1mAc72 | 0.62 | 34 | 48 | 4.4 | 1.2 | 0.1 | 0.1 | 5.92 | 0.41 | 0 | 6.9 | 8.7797854 | 100 | 1 |
| Aciprestes | Vineyards | 1 | Medium | hmAc81 | 1.98 | 206 | 159 | 6.18 | 1.33 | 0.13 | 0.07 | 7.71 | 1.27 | 0 | 7.3 | 9.0386469 | 100 | 0 |
| Aciprestes | Vineyards | 0 | Medium | nh1mAc82 | 1.19 | 73 | 80 | 7.9 | 1.5 | 0.2 | 0.4 | 9.95 | 0.77 | 0 | 6.9 | 8.9603221 | 100 | 0 |
| Aciprestes | Hedgerows | 0 | Medium | nh2mAcM1 | 2.62 | 12 | 92 | 4.08 | 1.44 | 0.18 | 0.06 | 5.77 | 1.27 | 0 | 6.8 | 11.9518 | 100 | 0 |
| Aciprestes | Hedgerows | 0 | Gross | nh2gAcM2 | 1.61 | 4 | 66 | 3.07 | 1.87 | 0.14 | 0.04 | 5.12 | 0.93 | 0 | 6.7 | 10.063711 | 100 | 2 |
| Aciprestes | Hedgerows | 0 | Medium | nh2mAcM3 | 1.75 | 20 | 123 | 3.39 | 1.28 | 0.23 | 0.03 | 4.93 | 0.91 | 0 | 6.7 | 11.130978 | 100 | 2 |
| Aciprestes | Hedgerows | 0 | Medium | nh2mAcM4 | 0.56 | 4 | 56 | 3.30 | 1.81 | 0.10 | 0.03 | 5.24 | 0.38 | 0 | 6.6 | 8.6045747 | 100 | 1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, L.; Oliveira, I.; Raimundo, F.; Torres, L.; Marques, G. Soil Chemical Properties Barely Perturb the Abundance of Entomopathogenic Fusarium oxysporum: A Case Study Using a Generalized Linear Mixed Model for Microbial Pathogen Occurrence Count Data. Pathogens 2018, 7, 89. https://doi.org/10.3390/pathogens7040089
Sharma L, Oliveira I, Raimundo F, Torres L, Marques G. Soil Chemical Properties Barely Perturb the Abundance of Entomopathogenic Fusarium oxysporum: A Case Study Using a Generalized Linear Mixed Model for Microbial Pathogen Occurrence Count Data. Pathogens. 2018; 7(4):89. https://doi.org/10.3390/pathogens7040089
Chicago/Turabian StyleSharma, Lav, Irene Oliveira, Fernando Raimundo, Laura Torres, and Guilhermina Marques. 2018. "Soil Chemical Properties Barely Perturb the Abundance of Entomopathogenic Fusarium oxysporum: A Case Study Using a Generalized Linear Mixed Model for Microbial Pathogen Occurrence Count Data" Pathogens 7, no. 4: 89. https://doi.org/10.3390/pathogens7040089
APA StyleSharma, L., Oliveira, I., Raimundo, F., Torres, L., & Marques, G. (2018). Soil Chemical Properties Barely Perturb the Abundance of Entomopathogenic Fusarium oxysporum: A Case Study Using a Generalized Linear Mixed Model for Microbial Pathogen Occurrence Count Data. Pathogens, 7(4), 89. https://doi.org/10.3390/pathogens7040089