ApiAP2 Transcription Factors in Apicomplexan Parasites


Abstract
:1. Introduction
2. Structure of ApiAP2 Transcription Factors
3. DNA Binding Motifs of ApiAP2 Transcription Factors
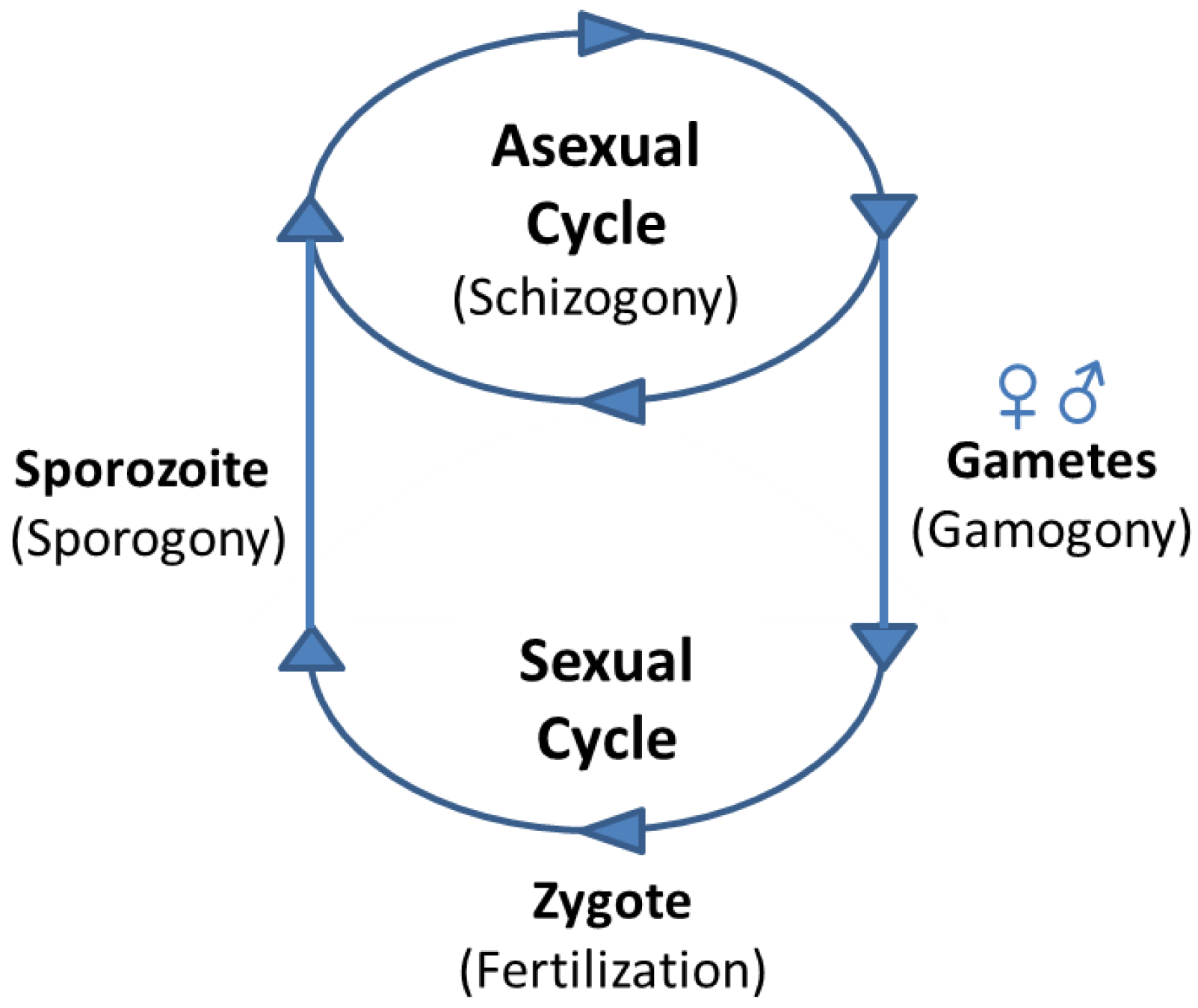
4. Developmental Transitions during Apicomplexan Life Cycles
5. ApiAP2s of Malaria Parasites
5.1. Asexual Stages
5.2. Sexual Differentiation
5.3. Gametogenesis and Sporogony
5.4. Liver Stages
6. ApiAP2 Transcription Factors in Toxoplasma
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. WHO Malaria Report 2017; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Khalil, I.A.; Troeger, C.; Rao, P.C.; Blacker, B.F.; Brown, A.; Brewer, T.G.; Colombara, D.V.; De Hostos, E.L.; Engmann, C.; Guerrant, R.L.; et al. Morbidity, mortality, and long-term consequences associated with diarrhoea from Cryptosporidium infection in children younger than 5 years: A meta-analyses study. Lancet Glob. Health 2018, 6, e758–e768. [Google Scholar] [CrossRef]
- Menard, D.; Dondorp, A. Antimalarial Drug Resistance: A Threat to Malaria Elimination. Cold Spring Harb. Perspect. Med. 2017, 7. [Google Scholar] [CrossRef]
- Montazeri, M.; Mehrzadi, S.; Sharif, M.; Sarvi, S.; Tanzifi, A.; Aghayan, S.A.; Daryani, A. Drug Resistance in Toxoplasma gondii. Front. Microbiol. 2018, 9, 2587. [Google Scholar] [CrossRef] [PubMed]
- Bozdech, Z.; Llinás, M.; Pulliam, B.L.; Wong, E.D.; Zhu, J.; DeRisi, J.L. The Transcriptome of the Intraerythrocytic Developmental Cycle of Plasmodium falciparum. PLoS Biol. 2003, 1, 085–100. [Google Scholar] [CrossRef] [PubMed]
- Le Roch, K.G.; Zhou, Y.; Blair, P.L.; Grainger, M.; Moch, J.K.; Haynes, J.D.; De La Vega, P.; Holder, A.A.; Batalov, S.; Carucci, D.J.; et al. Discovery of gene function by expression profiling of the malaria parasite life cycle. Science 2003, 301, 1503–1508. [Google Scholar] [CrossRef] [PubMed]
- Radke, J.R.; Behnke, M.S.; Mackey, A.J.; Radke, J.B.; Roos, D.S.; White, M.W. The transcriptome of Toxoplasma gondii. BMC Biol. 2005, 3, 26. [Google Scholar] [CrossRef] [PubMed]
- Behnke, M.S.; Radke, J.B.; Smith, A.T.; Sullivan, W.J., Jr.; White, M.W. The transcription of bradyzoite genes in Toxoplasma gondii is controlled by autonomous promoter elements. Mol. Microbiol. 2008, 68, 1502–1518. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Barragan, M.J.; Lemieux, J.; Quinones, M.; Williamson, K.C.; Molina-Cruz, A.; Cui, K.; Barillas-Mury, C.; Zhao, K.; Su, X.Z. Directional gene expression and antisense transcripts in sexual and asexual stages of Plasmodium falciparum. BMC Genom. 2011, 12, 587. [Google Scholar] [CrossRef]
- Duffy, M.F.; Selvarajah, S.A.; Josling, G.A.; Petter, M. Epigenetic regulation of the Plasmodium falciparum genome. Brief. Funct. Genom. 2014, 13, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Latchman, D.S. Transcription factors: An overview. Int. J. Biochem. Cell Biol. 1997, 29, 1305–1312. [Google Scholar] [CrossRef] [Green Version]
- Aravind, L.; Iyer, L.M.; Wellems, T.E.; Miller, L.H. Plasmodium Biology: Genomic Gleanings. Cell 2003, 115, 771–785. [Google Scholar] [CrossRef]
- Templeton, T.J.; Iyer, L.M.; Anantharaman, V.; Enomoto, S.; Abrahante, J.E.; Subramanian, G.M.; Hoffman, S.L.; Abrahamsen, M.S.; Aravind, L. Comparative analysis of apicomplexa and genomic diversity in eukaryotes. Genome Res. 2004, 14, 1686–1695. [Google Scholar] [CrossRef] [PubMed]
- Coulson, R.M.R.; Hall, N.; Ouzounis, C.A. Comparative genomics of transcriptional control in the human malaria parasite Plasmodium falciparum. Genome Res. 2004, 14, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- Boschet, C.; Gissot, M.; Briquet, S.; Hamid, Z.; Claudel-Renard, C.; Vaquero, C. Characterization of PfMyb1 transcription factor during erythrocytic development of 3D7 and F12 Plasmodium falciparum clones. Mol. Biochem. Parasitol. 2004, 138, 159–163. [Google Scholar] [CrossRef]
- Gissot, M.; Ting, L.M.; Daly, T.M.; Bergman, L.W.; Sinnis, P.; Kim, K. High mobility group protein HMGB2 is a critical regulator of plasmodium oocyst development. J. Biol. Chem. 2008, 283, 17030–17038. [Google Scholar] [CrossRef]
- Briquet, S.; Boschet, C.; Gissot, M.; Tissandie, E.; Sevilla, E.; Franetich, J.F.; Thiery, I.; Hamid, Z.; Bourgouin, C.; Vaquero, C. High-mobility-group box nuclear factors of Plasmodium falciparum. Eukaryot. Cell 2006, 5, 672–682. [Google Scholar] [CrossRef] [PubMed]
- Iyer, L.M.; Anantharaman, V.; Wolf, M.Y.; Aravind, L. Comparative genomics of transcription factors and chromatin proteins in parasitic protists and other eukaryotes. Int. J. Parasitol. 2008, 38, 1–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komaki-Yasuda, K.; Okuwaki, M.; Nagata, K.; Kawazu, S.; Kano, S. Identification of a novel and unique transcription factor in the intraerythrocytic stage of Plasmodium falciparum. PLoS ONE 2013, 8, e74701. [Google Scholar] [CrossRef] [PubMed]
- Balaji, S.; Babu, M.M.; Iyer, L.M.; Aravind, L. Discovery of the principal specific transcription factors of Apicomplexa and their implication for the evolution of the AP2-integrase DNA binding domains. Nucleic Acids Res. 2005, 33, 3994–4006. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Kigawa, T.; Seki, M.; Shinozaki, K.; Yokoyama, S. DNA-binding domains of plant-specific transcription factors: Structure, function, and evolution. Trends Plant Sci. 2013, 18, 267–276. [Google Scholar] [CrossRef]
- McFadden, G.I.; Reith, M.E.; Munholland, J.; Lang-Unnasch, N. Plastid in human parasites. Nature 1996, 381, 482. [Google Scholar] [CrossRef]
- Waller, R.F.; McFadden, G.I. The apicoplast: A review of the derived plastid of apicomplexan parasites. Curr. Issues Mol. Biol. 2005, 7, 57–79. [Google Scholar]
- Janouskovec, J.; Horak, A.; Obornik, M.; Lukes, J.; Keeling, P.J. A common red algal origin of the apicomplexan, dinoflagellate, and heterokont plastids. Proc. Natl. Acad. Sci. USA 2010, 107, 10949–10954. [Google Scholar] [CrossRef] [Green Version]
- Foth, B.J.; McFadden, G.I. The apicoplast: A plastid in Plasmodium falciparum and other Apicomplexan parasites. Int. Rev. Cytol. 2003, 224, 57–110. [Google Scholar]
- Aurrecoechea, C.; Barreto, A.; Basenko, E.Y.; Brestelli, J.; Brunk, B.P.; Cade, S.; Crouch, K.; Doherty, R.; Falke, D.; Fischer, S.; et al. EuPathDB: The eukaryotic pathogen genomics database resource. Nucleic Acids Res. 2017, 45, D581–D591. [Google Scholar] [CrossRef]
- Oberstaller, J.; Pumpalova, Y.; Schieler, A.; Llinas, M.; Kissinger, J.C. The Cryptosporidium parvum ApiAP2 gene family: Insights into the evolution of apicomplexan AP2 regulatory systems. Nucleic Acids Res. 2014, 42, 8271–8284. [Google Scholar] [CrossRef]
- Oberstaller, J.; Joseph, S.J.; Kissinger, J.C. Genome-wide upstream motif analysis of Cryptosporidium parvum genes clustered by expression profile. BMC Genom. 2013, 14, 516. [Google Scholar] [CrossRef]
- Oehring, S.C.; Woodcroft, B.J.; Moes, S.; Wetzel, J.; Dietz, O.; Pulfer, A.; Dekiwadia, C.; Maeser, P.; Flueck, C.; Witmer, K.; et al. Organellar proteomics reveals hundreds of novel nuclear proteins in the malaria parasite Plasmodium falciparum. Genome Biol. 2012, 13, R108. [Google Scholar] [CrossRef]
- Lindner, S.E.; De Silva, E.K.; Keck, J.L.; Llinás, M. Structural determinants of DNA binding by a P. falciparum ApiAP2 transcriptional regulator. J. Mol. Biol. 2010, 395, 558–567. [Google Scholar] [CrossRef]
- Aravind, L.; Landsman, D. AT-hook motifs identified in a wide variety of DNA-binding proteins. Nucleic Acids Res. 1998, 26, 4413–4421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, T.L.; De Silva, E.K.; Olszewski, K.L.; Elemento, O.; Llinas, M. Identification and genome-wide prediction of DNA binding specificities for the ApiAP2 family of regulators from the malaria parasite. PLoS Pathog. 2010, 6, e1001165. [Google Scholar] [CrossRef]
- De Silva, E.K.; Gehrke, A.R.; Olszewski, K.; Leon, I.; Chahal, J.S.; Bulyk, M.L.; Llinas, M. Specific DNA-binding by apicomplexan AP2 transcription factors. Proc. Natl. Acad. Sci. USA 2008, 105, 8393–8398. [Google Scholar] [CrossRef]
- Kaneko, I.; Iwanaga, S.; Kato, T.; Kobayashi, I.; Yuda, M. Genome-Wide Identification of the Target Genes of AP2-O, a Plasmodium AP2-Family Transcription Factor. PLoS Pathog. 2015, 11, e1004905. [Google Scholar] [CrossRef] [PubMed]
- Flueck, C.; Bartfai, R.; Niederwieser, I.; Witmer, K.; Alako, B.T.; Moes, S.; Bozdech, Z.; Jenoe, P.; Stunnenberg, H.G.; Voss, T.S. A major role for the Plasmodium falciparum ApiAP2 protein PfSIP2 in chromosome end biology. PLoS Pathog. 2010, 6, e1000784. [Google Scholar] [CrossRef]
- Santos, J.M.; Josling, G.; Ross, P.; Joshi, P.; Orchard, L.; Campbell, T.; Schieler, A.; Cristea, I.M.; Llinas, M. Red Blood Cell Invasion by the Malaria Parasite Is Coordinated by the PfAP2-I Transcription Factor. Cell Host Microbe 2017, 21, 731–741 e710. [Google Scholar] [CrossRef] [PubMed]
- Toenhake, C.G.; Fraschka, S.A.; Vijayabaskar, M.S.; Westhead, D.R.; van Heeringen, S.J.; Bartfai, R. Chromatin Accessibility-Based Characterization of the Gene Regulatory Network Underlying Plasmodium falciparum Blood-Stage Development. Cell Host Microbe 2018, 23, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.L.; Tena, J.J.; Bancells, C.; Cortes, A.; Gomez-Skarmeta, J.L.; Gomez-Diaz, E. Characterization of the accessible genome in the human malaria parasite Plasmodium falciparum. Nucleic Acids Res. 2018, 46, 9414–9431. [Google Scholar] [CrossRef]
- Lippuner, C.; Ramakrishnan, C.; Basso, W.U.; Schmid, M.W.; Okoniewski, M.; Smith, N.C.; Hässig, M.; Deplazes, P.; Hehl, A.B. RNA-Seq analysis during the life cycle of Cryptosporidium parvum reveals significant differential gene expression between proliferating stages in the intestine and infectious sporozoites. Int. J. Parasitol. 2018, 48, 413–422. [Google Scholar] [CrossRef]
- Behnke, M.S.; Wootton, J.C.; Lehmann, M.M.; Radke, J.B.; Lucas, O.; Nawas, J.; Sibley, L.D.; White, M.W. Coordinated progression through two subtranscriptomes underlies the tachyzoite cycle of Toxoplasma gondii. PLoS ONE 2010, 5, e12354. [Google Scholar] [CrossRef]
- Behnke, M.S.; Zhang, T.P.; Dubey, J.P.; Sibley, L.D. Toxoplasma gondii merozoite gene expression analysis with comparison to the life cycle discloses a unique expression state during enteric development. BMC Genom. 2014, 15, 350. [Google Scholar] [CrossRef] [Green Version]
- Gaji, R.Y.; Behnke, M.S.; Lehmann, M.M.; White, M.W.; Carruthers, V.B. Cell cycle-dependent, intercellular transmission of Toxoplasma gondii is accompanied by marked changes in parasite gene expression. Mol. Microbiol. 2011, 79, 192–204. [Google Scholar] [CrossRef]
- Mirhashemi, M.E.; Noubary, F.; Chapman-Bonofiglio, S.; Tzipori, S.; Huggins, G.S.; Widmer, G. Transcriptome analysis of pig intestinal cell monolayers infected with Cryptosporidium parvum asexual stages. Parasites Vectors 2018, 11, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelle, K.G.; Oh, K.; Buchholz, K.; Narasimhan, V.; Joice, R.; Milner, D.A.; Brancucci, N.M.; Ma, S.; Voss, T.S.; Ketman, K.; et al. Transcriptional profiling defines dynamics of parasite tissue sequestration during malaria infection. Genome Med. 2015, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Lasonder, E.; Rijpma, S.R.; van Schaijk, B.C.; Hoeijmakers, W.A.; Kensche, P.R.; Gresnigt, M.S.; Italiaander, A.; Vos, M.W.; Woestenenk, R.; Bousema, T.; et al. Integrated transcriptomic and proteomic analyses of P. falciparum gametocytes: Molecular insight into sex-specific processes and translational repression. Nucleic Acids Res. 2016, 44, 6087–6101. [Google Scholar] [CrossRef] [PubMed]
- Otto, T.D.; Wilinski, D.; Assefa, S.; Keane, T.M.; Sarry, L.R.; Bohme, U.; Lemieux, J.; Barrell, B.; Pain, A.; Berriman, M.; et al. New insights into the blood-stage transcriptome of Plasmodium falciparum using RNA-Seq. Mol. Microbiol. 2010, 76, 12–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartfai, R.; Hoeijmakers, W.A.; Salcedo-Amaya, A.M.; Smits, A.H.; Janssen-Megens, E.; Kaan, A.; Treeck, M.; Gilberger, T.W.; Francoijs, K.J.; Stunnenberg, H.G. H2A.Z demarcates intergenic regions of the plasmodium falciparum epigenome that are dynamically marked by H3K9ac and H3K4me3. PLoS Pathog. 2010, 6, e1001223. [Google Scholar] [CrossRef]
- Zanghi, G.; Vembar, S.S.; Baumgarten, S.; Ding, S.; Guizetti, J.; Bryant, J.M.; Mattei, D.; Jensen, A.T.R.; Renia, L.; Goh, Y.S.; et al. A Specific PfEMP1 Is Expressed in P. falciparum Sporozoites and Plays a Role in Hepatocyte Infection. Cell Rep. 2018, 22, 2951–2963. [Google Scholar] [CrossRef]
- Painter, H.J.; Carrasquilla, M.; Llinás, M. Capturing in vivo RNA transcriptional dynamics from the malaria parasite Plasmodium falciparum. Genome Res. 2017, 27, 1074–1086. [Google Scholar] [CrossRef] [PubMed]
- Painter, H.J.; Chung, N.C.; Sebastian, A.; Albert, I.; Storey, J.D.; Llinás, M. Genome-wide real-time in vivo transcriptional dynamics during Plasmodium falciparum blood-stage development. Nat. Commun. 2018, 9, 2656. [Google Scholar] [CrossRef]
- Bunnik, E.M.; Chung, D.W.; Hamilton, M.; Ponts, N.; Saraf, A.; Prudhomme, J.; Florens, L.; Le Roch, K.G. Polysome profiling reveals translational control of gene expression in the human malaria parasite Plasmodium falciparum. Genome Biol. 2013, 14, R128. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, K.R.; Fritz, H.M.; Chen, X.; Durbin-Johnson, B.; Rocke, D.M.; Ferguson, D.J.; Conrad, P.A.; Boothroyd, J.C. Identification of tissue cyst wall components by transcriptome analysis of in vivo and in vitro Toxoplasma gondii bradyzoites. Eukaryot. Cell 2011, 10, 1637–1647. [Google Scholar] [CrossRef]
- Fritz, H.M.; Buchholz, K.R.; Chen, X.; Durbin-Johnson, B.; Rocke, D.M.; Conrad, P.A.; Boothroyd, J.C. Transcriptomic analysis of toxoplasma development reveals many novel functions and structures specific to sporozoites and oocysts. PLoS ONE 2012, 7, e29998. [Google Scholar] [CrossRef]
- Mota, M.M.; Rodriguez, A. Invasion of mammalian host cells by Plasmodium sporozoites. Bioessays News Rev. Mol. Cell. Dev. Biol. 2002, 24, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Field, J.W.; Shute, P.G. The Microscopic Diagnosis of Human Malaria II; Govt. Pr.: Kuala Lumpur, Malaysia, 1956. [Google Scholar]
- Silvestrini, F.; Alano, P.; Williams, J.L. Commitment to the production of male and female gametocytes in the human malaria parasite Plasmodium falciparum. Parasitology 2000, 121 Pt 5, 465–471. [Google Scholar] [CrossRef]
- Bennink, S.; Kiesow, M.J.; Pradel, G. The development of malaria parasites in the mosquito midgut. Cell. Microbiol. 2016, 18, 905–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matuschewski, K. Getting infectious: Formation and maturation of Plasmodium sporozoites in the Anopheles vector. Cell. Microbiol. 2006, 8, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Baton, L.A.; Ranford-Cartwright, L.C. How do malaria ookinetes cross the mosquito midgut wall? Trends Parasitol. 2005, 21, 22–28. [Google Scholar] [CrossRef]
- Gubbels, M.J.; White, M.; Szatanek, T. The cell cycle and Toxoplasma gondii cell division: Tightly knit or loosely stitched? Int. J. Parasitol. 2008, 38, 1343–1358. [Google Scholar] [CrossRef]
- Bouzid, M.; Hunter, P.R.; Chalmers, R.M.; Tyler, K.M. Cryptosporidium pathogenicity and virulence. Clin. Microbiol. Rev. 2013, 26, 115–134. [Google Scholar] [CrossRef]
- Mauzy, M.J.; Enomoto, S.; Lancto, C.A.; Abrahamsen, M.S.; Rutherford, M.S. The Cryptosporidium parvum transcriptome during in vitro development. PLoS ONE 2012, 7, e31715. [Google Scholar] [CrossRef] [PubMed]
- Bushell, E.; Gomes, A.R.; Sanderson, T.; Anar, B.; Girling, G.; Herd, C.; Metcalf, T.; Modrzynska, K.; Schwach, F.; Martin, R.E.; et al. Functional Profiling of a Plasmodium Genome Reveals an Abundance of Essential Genes. Cell 2017, 170, 260–272. [Google Scholar] [CrossRef]
- Modrzynska, K.; Pfander, C.; Chappell, L.; Yu, L.; Suarez, C.; Dundas, K.; Gomes, A.R.; Goulding, D.; Rayner, J.C.; Choudhary, J.; et al. A Knockout Screen of ApiAP2 Genes Reveals Networks of Interacting Transcriptional Regulators Controlling the Plasmodium Life Cycle. Cell Host Microbe 2017, 21, 11–22. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Z.; Cui, H.; Jiang, Y.; Yang, Z.; Wang, X.; Gao, H.; Liu, C.; Zhang, S.; Su, X.-Z.; et al. Systematic CRISPR-Cas9-Mediated Modifications of Plasmodium yoelii ApiAP2 Genes Reveal Functional Insights into Parasite Development. mBio 2017, 8, e01986-17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, C.; Otto, T.D.; Oberstaller, J.; Liao, X.; Adapa, S.R.; Udenze, K.; Bronner, I.F.; Casandra, D.; Mayho, M.; et al. Uncovering the essential genes of the human malaria parasite Plasmodium falciparum by saturation mutagenesis. Science 2018, 360. [Google Scholar] [CrossRef]
- Sierra-Miranda, M.; Vembar, S.-S.; Delgadillo, D.M.; Ávila-López, P.A.; Herrera-Solorio, A.-M.; Lozano Amado, D.; Vargas, M.; Hernandez-Rivas, R. PfAP2Tel, harbouring a non-canonical DNA-binding AP2 domain, binds to Plasmodium falciparum telomeres. Cell. Microbiol. 2017, 19, e12742. [Google Scholar] [CrossRef]
- Balu, B.; Singh, N.; Maher, S.P.; Adams, J.H. A genetic screen for attenuated growth identifies genes crucial for intraerythrocytic development of Plasmodium falciparum. PLoS ONE 2010, 5, e13282. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, S.; Kaneko, I.; Kato, T.; Yuda, M. Identification of an AP2-family protein that is critical for malaria liver stage development. PLoS ONE 2012, 7, e47557. [Google Scholar] [CrossRef]
- Yuda, M.; Iwanaga, S.; Shigenobu, S.; Mair, G.R.; Janse, C.J.; Waters, A.P.; Kato, T.; Kaneko, I. Identification of a transcription factor in the mosquito-invasive stage of malaria parasites. Mol. Microbiol. 2009, 71, 1402–1414. [Google Scholar] [CrossRef]
- Sinha, A.; Hughes, K.R.; Modrzynska, K.K.; Otto, T.D.; Pfander, C.; Dickens, N.J.; Religa, A.A.; Bushell, E.; Graham, A.L.; Cameron, R.; et al. A cascade of DNA-binding proteins for sexual commitment and development in Plasmodium. Nature 2014, 507, 253–257. [Google Scholar] [CrossRef]
- Kafsack, B.F.; Rovira-Graells, N.; Clark, T.G.; Bancells, C.; Crowley, V.M.; Campino, S.G.; Williams, A.E.; Drought, L.G.; Kwiatkowski, D.P.; Baker, D.A.; et al. A transcriptional switch underlies commitment to sexual development in malaria parasites. Nature 2014, 507, 248–252. [Google Scholar] [CrossRef] [Green Version]
- Ikadai, H.; Shaw Saliba, K.; Kanzok, S.M.; McLean, K.J.; Tanaka, T.Q.; Cao, J.; Williamson, K.C.; Jacobs-Lorena, M. Transposon mutagenesis identifies genes essential for Plasmodium falciparum gametocytogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, E1676–E1684. [Google Scholar] [CrossRef] [Green Version]
- Bischoff, E.; Vaquero, C. In silico and biological survey of transcription-associated proteins implicated in the transcriptional machinery during the erythrocytic development of Plasmodium falciparum. BMC Genom. 2010, 11, 34. [Google Scholar] [CrossRef]
- Yuda, M.; Iwanaga, S.; Kaneko, I.; Kato, T. Global transcriptional repression: An initial and essential step for Plasmodium sexual development. PNAS 2015, 112, 12824–12829. [Google Scholar] [CrossRef] [Green Version]
- Maier, A.G.; Rug, M.; O’Neill, M.T.; Brown, M.; Chakravorty, S.; Szestak, T.; Chesson, J.; Wu, Y.; Hughes, K.; Coppel, R.L.; et al. Exported proteins required for virulence and rigidity of Plasmodium falciparum-infected human erythrocytes. Cell 2008, 134, 48–61. [Google Scholar] [CrossRef]
- Yuda, M.; Iwanaga, S.; Shigenobu, S.; Kato, T.; Kaneko, I. Transcription factor AP2-Sp and its target genes in malarial sporozoites. Mol. Microbiol. 2010, 75, 854–863. [Google Scholar] [CrossRef] [Green Version]
- Martins, R.M.; Macpherson, C.R.; Claes, A.; Scheidig-Benatar, C.; Sakamoto, H.; Yam, X.Y.; Preiser, P.; Goel, S.; Wahlgren, M.; Sismeiro, O.; et al. An ApiAP2 member regulates expression of clonally variant genes of the human malaria parasite Plasmodium falciparum. Sci. Rep. 2017, 7, 14042. [Google Scholar] [CrossRef] [Green Version]
- Cubi, R.; Vembar, S.S.; Biton, A.; Franetich, J.-F.; Bordessoulles, M.; Sossau, D.; Zanghi, G.; Bosson-Vanga, H.; Benard, M.; Moreno, A.; et al. Laser capture microdissection enables transcriptomic analysis of dividing and quiescent liver stages of Plasmodium relapsing species. Cell. Microbiol. 2017, 19, e12735. [Google Scholar] [CrossRef] [Green Version]
- Jex, A.; Mueller, I.; Kappe, S.; Mikolajcjak, S.; Sattabongkot, J.; Patrapuvich, R.; Lindner, S.; Flannery, E.; Koepfli, C.; Ansell, B.; et al. Transcriptome and histone epigenome of Plasmodium vivax salivary-gland sporozoites point to tight regulatory control and potential mechanisms for liver-stage differentiation. Int. J. Parasitol. 2019, in press. [Google Scholar]
- Cobbold, S.A.; Santos, J.M.; Ochoa, A.; Perlman, D.H.; Llinas, M. Proteome-wide analysis reveals widespread lysine acetylation of major protein complexes in the malaria parasite. Sci. Rep. 2016, 6, 19722. [Google Scholar] [CrossRef]
- Young, J.A.; Johnson, J.R.; Benner, C.; Yan, S.F.; Chen, K.; Le Roch, K.G.; Zhou, Y.; Winzeler, E.A. In silico discovery of transcription regulatory elements in Plasmodium falciparum. BMC Genom. 2008, 9, 70. [Google Scholar] [CrossRef] [PubMed]
- Iengar, P.; Joshi, N.V. Identification of putative regulatory motifs in the upstream regions of co-expressed functional groups of genes in Plasmodium falciparum. BMC Genom. 2009, 10, 18. [Google Scholar] [CrossRef]
- Essien, K.; Stoeckert, C.J. Conservation and divergence of known apicomplexan transcriptional regulons. BMC Genom. 2010, 11, 147. [Google Scholar] [CrossRef]
- Josling, G.A.; Petter, M.; Oehring, S.C.; Gupta, A.P.; Dietz, O.; Wilson, D.W.; Schubert, T.; Langst, G.; Gilson, P.R.; Crabb, B.S.; et al. A Plasmodium Falciparum Bromodomain Protein Regulates Invasion Gene Expression. Cell Host Microbe 2015, 17, 741–751. [Google Scholar] [CrossRef] [Green Version]
- Cowell, A.; Winzeler, E. Exploration of the Plasmodium falciparum Resistome and Druggable Genome Reveals New Mechanisms of Drug Resistance and Antimalarial Targets. Microbiol. Insights 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Wendler, J.P.; Okombo, J.; Amato, R.; Miotto, O.; Kiara, S.M.; Mwai, L.; Pole, L.; O’Brien, J.; Manske, M.; Alcock, D.; et al. A genome wide association study of Plasmodium falciparum susceptibility to 22 antimalarial drugs in Kenya. PLoS ONE 2014, 9, e96486. [Google Scholar] [CrossRef]
- van Biljon, R.; Niemand, J.; van Wyk, R.; Clark, K.; Verlinden, B.; Abrie, C.; von Grüning, H.; Smidt, W.; Smit, A.; Reader, J.; et al. Inducing controlled cell cycle arrest and re-entry during asexual proliferation of Plasmodium falciparum malaria parasites. Sci. Rep. 2018, 8, 16581. [Google Scholar] [CrossRef]
- Bankier, A.T.; Spriggs, H.F.; Fartmann, B.; Konfortov, B.A.; Madera, M.; Vogel, C.; Teichmann, S.A.; Ivens, A.; Dear, P.H. Integrated mapping, chromosomal sequencing and sequence analysis of Cryptosporidium parvum. Genome Res. 2003, 13, 1787–1799. [Google Scholar] [CrossRef]
- Ling, K.-H.; Rajandream, M.-A.; Rivailler, P.; Ivens, A.; Yap, S.-J.; Madeira, A.M.B.N.; Mungall, K.; Billington, K.; Yee, W.-Y.; Bankier, A.T.; et al. Sequencing and analysis of chromosome 1 of Eimeria tenella reveals a unique segmental organization. Genome Res. 2007, 17, 311–319. [Google Scholar] [CrossRef] [Green Version]
- Voss, T.S.; Kaestli, M.; Vogel, D.; Bopp, S.; Beck, H.P. Identification of nuclear proteins that interact differentially with Plasmodium falciparum var gene promoters. Mol. Microbiol. 2003, 48, 1593–1607. [Google Scholar] [CrossRef] [Green Version]
- Rono, M.K.; Nyonda, M.A.; Simam, J.J.; Ngoi, J.M.; Mok, S.; Kortok, M.M.; Abdullah, A.S.; Elfaki, M.M.; Waitumbi, J.N.; El-Hassan, I.M.; et al. Adaptation of Plasmodium falciparum to its transmission environment. Nat. Ecol. Evol. 2018, 2, 377–387. [Google Scholar] [CrossRef]
- Honma, H.; Niikura, M.; Kobayashi, F.; Horii, T.; Mita, T.; Endo, H.; Hirai, M. Mutation tendency of mutator Plasmodium berghei with proofreading-deficient DNA polymerase δ. Sci. Rep. 2016, 6, 36971. [Google Scholar] [CrossRef] [Green Version]
- Claessens, A.; Affara, M.; Assefa, S.A.; Kwiatkowski, D.P.; Conway, D.J. Culture adaptation of malaria parasites selects for convergent loss-of-function mutants. Sci. Rep. 2017, 7, 41303. [Google Scholar] [CrossRef] [Green Version]
- Beri, D.; Balan, B.; Chaubey, S.; Subramaniam, S.; Surendra, B.; Tatu, U. A disrupted transsulphuration pathway results in accumulation of redox metabolites and induction of gametocytogenesis in malaria. Sci. Rep. 2017, 7, 40213. [Google Scholar] [CrossRef] [Green Version]
- Brancucci, N.M.B.; Gerdt, J.P.; Wang, C.; De Niz, M.; Philip, N.; Adapa, S.R.; Zhang, M.; Hitz, E.; Niederwieser, I.; Boltryk, S.D.; et al. Lysophosphatidylcholine Regulates Sexual Stage Differentiation in the Human Malaria Parasite Plasmodium falciparum. Cell 2017, 171, 1532–1544. [Google Scholar] [CrossRef]
- Rovira-Graells, N.; Gupta, A.P.; Planet, E.; Crowley, V.M.; Mok, S.; Ribas de Pouplana, L.; Preiser, P.R.; Bozdech, Z.; Cortés, A. Transcriptional variation in the malaria parasite Plasmodium falciparum. Genome Res. 2012, 22, 925–938. [Google Scholar] [CrossRef] [Green Version]
- Eksi, S.; Morahan, B.J.; Haile, Y.; Furuya, T.; Jiang, H.; Ali, O.; Xu, H.; Kiattibutr, K.; Suri, A.; Czesny, B.; et al. Plasmodium falciparum gametocyte development 1 (Pfgdv1) and gametocytogenesis early gene identification and commitment to sexual development. PLoS Pathog. 2012, 8, e1002964. [Google Scholar] [CrossRef]
- Day, K.P.; Karamalis, F.; Thompson, J.; Barnes, D.A.; Peterson, C.; Brown, H.; Brown, G.V.; Kemp, D.J. Genes necessary for expression of a virulence determinant and for transmission of Plasmodium falciparum are located on a 0.3-megabase region of chromosome 9. Proc. Natl. Acad. Sci. USA 1993, 90, 8292–8296. [Google Scholar] [CrossRef]
- Campino, S.; Benavente, E.D.; Assefa, S.; Thompson, E.; Drought, L.G.; Taylor, C.J.; Gorvett, Z.; Carret, C.K.; Flueck, C.; Ivens, A.C.; et al. Genomic variation in two gametocyte non-producing Plasmodium falciparum clonal lines. Malar. J. 2016, 15, 229. [Google Scholar] [CrossRef]
- Kent, R.S.; Modrzynska, K.K.; Cameron, R.; Philip, N.; Billker, O.; Waters, A.P. Inducible developmental reprogramming redefines commitment to sexual development in the malaria parasite Plasmodium berghei. Nat. Microbiol. 2018, 3, 1206–1213. [Google Scholar] [CrossRef]
- Josling, G.A.; Venezia, J.; Orchard, L.; Russell, T.J.; Painter, H.J.; Llinas, M. Regulation of sexual differentiation is linked to invasion in malaria parasites. bioRxiv 2019. [Google Scholar] [CrossRef]
- Tiburcio, M.; Dixon, M.W.; Looker, O.; Younis, S.Y.; Tilley, L.; Alano, P. Specific expression and export of the Plasmodium falciparum Gametocyte EXported Protein-5 marks the gametocyte ring stage. Malar J 2015, 14, 334. [Google Scholar] [CrossRef]
- Bancells, C.; Llorà-Batlle, O.; Poran, A.; Nötzel, C.; Rovira-Graells, N.; Elemento, O.; Kafsack, B.F.C.; Cortés, A. Revisiting the initial steps of sexual development in the malaria parasite Plasmodium falciparum. Nat. Microbiol. 2019, 4, 144–154. [Google Scholar] [CrossRef]
- Cortés, A.; Crowley, V.M.; Vaquero, A.; Voss, T.S. A View on the Role of Epigenetics in the Biology of Malaria Parasites. PLoS Pathog. 2012, 8, e1002943. [Google Scholar] [CrossRef]
- Coleman, B.I.; Skillman, K.M.; Jiang, R.H.Y.; Childs, L.M.; Altenhofen, L.M.; Ganter, M.; Leung, Y.; Goldowitz, I.; Kafsack, B.F.C.; Marti, M.; et al. A Plasmodium falciparum histone deacetylase regulates antigenic variation and gametocyte conversion. Cell Host Microbe 2014, 16, 177–186. [Google Scholar] [CrossRef]
- Brancucci, N.M.B.; Bertschi, N.L.; Zhu, L.; Niederwieser, I.; Chin, W.H.; Wampfler, R.; Freymond, C.; Rottmann, M.; Felger, I.; Bozdech, Z.; et al. Heterochromatin protein 1 secures survival and transmission of malaria parasites. Cell Host Microbe 2014, 16, 165–176. [Google Scholar] [CrossRef]
- Filarsky, M.; Fraschka, S.A.; Niederwieser, I.; Brancucci, N.M.B.; Carrington, E.; Carrió, E.; Moes, S.; Jenoe, P.; Bártfai, R.; Voss, T.S. GDV1 induces sexual commitment of malaria parasites by antagonizing HP1-dependent gene silencing. Science 2018, 359, 1259–1263. [Google Scholar] [CrossRef]
- Bunnik, E.M.; Cook, K.B.; Varoquaux, N.; Batugedara, G.; Prudhomme, J.; Cort, A.; Shi, L.; Andolina, C.; Ross, L.S.; Brady, D.; et al. Changes in genome organization of parasite-specific gene families during the Plasmodium transmission stages. Nat. Commun. 2018, 9, 1910. [Google Scholar] [CrossRef]
- Poran, A.; Notzel, C.; Aly, O.; Mencia-Trinchant, N.; Harris, C.T.; Guzman, M.L.; Hassane, D.C.; Elemento, O.; Kafsack, B.F.C. Single-cell RNA sequencing reveals a signature of sexual commitment in malaria parasites. Nature 2017, 551, 95–99. [Google Scholar] [CrossRef]
- Lesage, K.M.; Huot, L.; Mouveaux, T.; Courjol, F.; Saliou, J.-M.; Gissot, M. Cooperative binding of ApiAP2 transcription factors is crucial for the expression of virulence genes in Toxoplasma gondii. Nucleic Acids Res. 2018, 46, 6057–6068. [Google Scholar] [CrossRef]
- Pieszko, M.; Weir, W.; Goodhead, I.; Kinnaird, J.; Shiels, B. ApiAP2 Factors as Candidate Regulators of Stochastic Commitment to Merozoite Production in Theileria annulata. PLoS Negl. Trop. Dis. 2015, 9, e0003933. [Google Scholar] [CrossRef]
- Parobek, C.M.; Lin, J.T.; Saunders, D.L.; Barnett, E.J.; Lon, C.; Lanteri, C.A.; Balasubramanian, S.; Brazeau, N.; DeConti, D.K.; Garba, D.L.; et al. Selective sweep suggests transcriptional regulation may underlie Plasmodium vivax resilience to malaria control measures in Cambodia. Proc. Natl. Acad. Sci. USA 2016, 113, E8096–E8105. [Google Scholar] [CrossRef]
- Dharia, N.V.; Bright, A.T.; Westenberger, S.J.; Barnes, S.W.; Batalov, S.; Kuhen, K.; Borboa, R.; Federe, G.C.; McClean, C.M.; Vinetz, J.M.; et al. Whole-genome sequencing and microarray analysis of ex vivo Plasmodium vivax reveal selective pressure on putative drug resistance genes. Proc. Natl. Acad. Sci. USA 2010, 107, 20045–20050. [Google Scholar] [CrossRef] [Green Version]
- Chaubey, S.; Grover, M.; Tatu, U. Endoplasmic reticulum stress triggers gametocytogenesis in the malaria parasite. J. Biol. Chem. 2014, 289, 16662–16674. [Google Scholar] [CrossRef]
- Mair, G.R.; Lasonder, E.; Garver, L.S.; Franke-Fayard, B.M.D.; Carret, C.K.; Wiegant, J.C.A.G.; Dirks, R.W.; Dimopoulos, G.; Janse, C.J.; Waters, A.P. Universal features of post-transcriptional gene regulation are critical for Plasmodium zygote development. PLoS Pathog. 2010, 6, e1000767. [Google Scholar] [CrossRef]
- Mair, G.R.; Braks, J.A.M.; Garver, L.S.; Dimopoulos, G.; Hall, N.; Wiegant, J.C.A.G.; Dirks, R.W.; Khan, S.M.; Janse, C.J.; Waters, A.P. Translational Repression is essential for Plasmodium sexual development and mediated by a DDX6-type RNA helicase. Science 2006, 313, 667–669. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Lindner, S.; Miao, J. Translational regulation during stage transitions in malaria parasites. Ann. N. Y. Acad. Sci. 2015, 1342, 1–9. [Google Scholar] [CrossRef]
- Gomez-Diaz, E.; Yerbanga, R.S.; Lefevre, T.; Cohuet, A.; Rowley, M.J.; Ouedraogo, J.B.; Corces, V.G. Epigenetic regulation of Plasmodium falciparum clonally variant gene expression during development in Anopheles gambiae. Sci. Rep. 2017, 7, 40655. [Google Scholar] [CrossRef]
- Frevert, U. Sneaking in through the back entrance: The biology of malaria liver stages. Trends Parasitol. 2004, 20, 417–424. [Google Scholar] [CrossRef]
- Silvie, O.; Goetz, K.; Matuschewski, K. A sporozoite asparagine-rich protein controls initiation of Plasmodium liver stage development. PLoS Pathog. 2008, 4, e1000086. [Google Scholar] [CrossRef]
- Aly, A.S.; Lindner, S.E.; MacKellar, D.C.; Peng, X.; Kappe, S.H. SAP1 is a critical post-transcriptional regulator of infectivity in malaria parasite sporozoite stages. Mol. Microbiol. 2011, 79, 929–939. [Google Scholar] [CrossRef]
- Aly, A.S.; Mikolajczak, S.A.; Rivera, H.S.; Camargo, N.; Jacobs-Lorena, V.; Labaied, M.; Coppens, I.; Kappe, S.H. Targeted deletion of SAP1 abolishes the expression of infectivity factors necessary for successful malaria parasite liver infection. Mol. Microbiol. 2008, 69, 152–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helm, S.; Lehmann, C.; Nagel, A.; Stanway, R.R.; Horstmann, S.; Llinas, M.; Heussler, V.T. Identification and characterization of a liver stage-specific promoter region of the malaria parasite Plasmodium. PLoS ONE 2010, 5, e13653. [Google Scholar] [CrossRef] [PubMed]
- Cogswell, F.B. The hypnozoite and relapse in primate malaria. Clin. Microbiol. Rev. 1992, 5, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Voorberg-van der Wel, A.; Roma, G.; Gupta, D.K.; Schuierer, S.; Nigsch, F.; Carbone, W.; Zeeman, A.M.; Lee, B.H.; Hofman, S.O.; Faber, B.W.; et al. A comparative transcriptomic analysis of replicating and dormant liver stages of the relapsing malaria parasite Plasmodium cynomolgi. eLife 2017, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gural, N.; Mancio-Silva, L.; Miller, A.B.; Galstian, A.; Butty, V.L.; Levine, S.S.; Patrapuvich, R.; Desai, S.P.; Mikolajczak, S.A.; Kappe, S.H.I.; et al. In Vitro Culture, Drug Sensitivity, and Transcriptome of Plasmodium Vivax Hypnozoites. Cell Host Microbe 2018, 23, 395–406.e394. [Google Scholar] [CrossRef] [PubMed]
- White, M.W.; Radke, J.R.; Radke, J.B. Toxoplasma development—Turn the switch on or off? Cell. Microbiol. 2014, 16, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Szatanek, T.; Anderson-White, B.R.; Faugno-Fusci, D.M.; White, M.; Saeij, J.P.J.; Gubbels, M.-J. Cactin is essential for G1 progression in Toxoplasma gondii. Mol. Microbiol. 2012, 84, 566–577. [Google Scholar] [CrossRef]
- Walker, R.; Gissot, M.; Croken, M.M.; Huot, L.; Hot, D.; Kim, K.; Tomavo, S. The Toxoplasma nuclear factor TgAP2XI-4 controls bradyzoite gene expression and cyst formation. Mol. Microbiol. 2013, 87, 641–655. [Google Scholar] [CrossRef]
- Wang, J.; Dixon, S.E.; Ting, L.-M.; Liu, T.-K.; Jeffers, V.; Croken, M.M.; Calloway, M.; Cannella, D.; Hakimi, M.A.; Kim, K.; et al. Lysine acetyltransferase GCN5b interacts with AP2 factors and is required for Toxoplasma gondii proliferation. PLoS Pathog. 2014, 10, e1003830. [Google Scholar] [CrossRef]
- Hong, D.-P.; Radke, J.B.; White, M.W. Opposing Transcriptional Mechanisms Regulate Toxoplasma Development. mSphere 2017, 2, e00347-16. [Google Scholar] [CrossRef] [PubMed]
- Radke, J.B.; Lucas, O.; De Silva, E.K.; Ma, Y.; Sullivan, W.J., Jr.; Weiss, L.M.; Llinas, M.; White, M.W. ApiAP2 transcription factor restricts development of the Toxoplasma tissue cyst. Proc. Natl. Acad. Sci. USA 2013, 110, 6871–6876. [Google Scholar] [CrossRef] [Green Version]
- Croken, M.M.; Ma, Y.; Markillie, L.M.; Taylor, R.C.; Orr, G.; Weiss, L.M.; Kim, K. Distinct strains of Toxoplasma gondii feature divergent transcriptomes regardless of developmental stage. PLoS ONE 2014, 9, e111297. [Google Scholar] [CrossRef] [PubMed]
- Radke, J.B.; Worth, D.; Hong, D.; Huang, S.; Sullivan, W.J., Jr.; Wilson, E.H.; White, M.W. Transcriptional repression by ApiAP2 factors is central to chronic toxoplasmosis. PLoS Pathog. 2018, 14, e1007035. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Holmes, M.J.; Radke, J.B.; Hong, D.-P.; Liu, T.-K.; White, M.W.; Sullivan, W.J., Jr. Toxoplasma gondii AP2IX-4 Regulates Gene Expression during Bradyzoite Development. mSphere 2017, 2, e00054-17. [Google Scholar] [CrossRef] [PubMed]
- Klug, D.; Kehrer, J.; Frischknecht, F.; Singer, M. A synthetic promoter for multi-stage expression to probe complementary functions of Plasmodium adhesins. J. Cell Sci. 2018, 131, jcs210971. [Google Scholar] [CrossRef] [PubMed]
- Pino, P.; Sebastian, S.; Kim, E.A.; Bush, E.; Brochet, M.; Volkmann, K.; Kozlowski, E.; Llinás, M.; Billker, O.; Soldati-Favre, D. A tetracycline-repressible transactivator system to study essential genes in malaria parasites. Cell Host Microbe 2012, 12, 824–834. [Google Scholar] [CrossRef]
- Lai, B.-S.; Witola, W.H.; El Bissati, K.; Zhou, Y.; Mui, E.; Fomovska, A.; McLeod, R. Molecular target validation, antimicrobial delivery, and potential treatment of Toxoplasma gondii infections. Proc. Natl. Acad. Sci. USA 2012, 109, 14182–14187. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| P. falciparum ApiAP2 TF | Name | Ortholog in P. vivax | Ortholog in P. berghei | Ortholog in P. yoelii | Mutated in P. falciparum [67] | Disrupted in P. berghei | Disrupted in P. yoelii [66] | Phenotype (-| Block) | Motif Recognized [33,34] | Further Ref | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | PF3D7_0404100 | AP2-SP2 | PVX_001040 | PBANKA_1001800 | PY17X_1003200 | yes | yes [65] | yes | oocyst -| sporoblast | ||
| 2 | PF3D7_0420300 | PVX_090110 | PBANKA_0521700 | PY17X_0523100 | refractory | refractory [64,65] | yes | n.a. | CACACAC, GTGTTACAC | ||
| 3 | PF3D7_0516800 | AP2-O2 | PVX_080410 | PBANKA_1231600 | PY17X_1235000 | yes | yes [64,65] | yes | zygote -| ookinete | TGACATCA | |
| 4 | PF3D7_0604100 | SIP2 | PVX_113370 | PBANKA_0102900 | PY17X_0104500 | yes | refractory [64,65] | refractory | n.a. | GGTGCAC | [36] |
| 5 | PF3D7_0611200 | PVX_113695 | PBANKA_0109500 | PY17X_0111100 | yes | yes [64] | refractory | slow growth | |||
| 6 | PF3D7_0613800 | PVX_113825 | PBANKA_0112100 | PY17X_0113700 | refractory | refractory [64,65] | refractory | n.a. | ATAAGCCCA, CTCTAGAG | ||
| 7 | PF3D7_0622900 | AP2-Tel/SP3 | PVX_114260 | PBANKA_1121800 | PY17X_1123200 | yes | yes [65] | yes | sporozoites -| liver stage | [68,69] | |
| 8 | PF3D7_0730300 | AP2-L | PVX_081810 | PBANKA_0214400 | PY17X_0215800 | yes | yes [64,65,70] | refractory | sporozoites -| liver stage | AATTTCC | [70] |
| 9 | PF3D7_0802100 | n.a. | PBANKA_1228100 | PY17X_1231600 | refractory | refractory [64,65] | refractory | n.a. | CACACACA | ||
| 10 | PF3D7_0934400 | PVX_086995 | PBANKA_0835200 | PY17X_0838600 | refractory | refractory [64,65] | refractory | n.a. | |||
| 11 | PF3D7_1007700 | AP2-I | PVX_094580 | PBANKA_1205900 | PY17X_1209100 | refractory | refractory [65] | refractory | n.a. | GGGTCGACCC TCTTGCC GTGCACTA | [37] |
| 12 | PF3D7_1107800 | PVX_091065 | PBANKA_0939100 | PY17X_0941600 | refractory | refractory [64,65] | refractory | n.a. | AGCATAC | ||
| 13 | PF3D7_1115500 | PVX_091420 | PBANKA_0932300 | PY17X_0934300 | refractory | yes [64,65] | refractory | n.a. | |||
| 14 | PF3D7_1139300 | PVX_092570 | PBANKA_0909600 | PY17X_0911000 | yes | refractory [64,65] | refractory | n.a. | TAGAACAA | ||
| 15 | PF3D7_1143100 | AP2-O | PVX_092760 | PBANKA_0905900 | PY17X_0907300 | refractory | yes [64,65,71] | yes | ookinete -| oocyst | TAGCTA [71] | [71] |
| 16 | PF3D7_1222400 | PVX_123750 | n.a. | n.a. | yes | n.a. | n.a. | n.a. | TATATATA | ||
| 17 | PF3D7_1222600 | AP2-G | PVX_123760 | PBANKA_1437500 | PY17X_1440000 | yes | yes [64,65,72] | yes | asexual -| gametocyte | GxGTACxC [72] | [72,73] |
| 18 | PF3D7_1239200 | PVX_100910 | PBANKA_1453700 | PY17X_1456200 | refractory | refractory [65] | refractory | n.a. | TCTA(C/T)AA | ||
| 19 | PF3D7_1305200 | PVX_122095 | PBANKA_1403700 | PY17X_1405400 | yes | refractory [64,65] | refractory | n.a. | TGCACACAC | ||
| 20 | PF3D7_1317200 | AP2-G3 | PVX_122680 | PBANKA_1415700 | PY17X_1417400 | yes | refractory [65] | yes | asexual -| gametocyte | TAGCTCA(G/A)A | [74] |
| 21 | PF3D7_1342900 | PVX_083040 | PBANKA_1356000 | PY17X_1361700 | yes | refractory [65] | refractory | n.a. | GCGGGGC | [69] | |
| 22 | PF3D7_1350900 | AP2-O4 | PVX_083440 | PBANKA_1363700 | PY17X_1369400 | refractory | yes [64,65] | yes | ookinete -| oocyst | ATTCTAGAA | [75] |
| 23 | PF3D7_1408200 | AP2-G2 | PVX_086035 | PBANKA_1034300 | PY17X_1036700 | yes | yes [64,65,72,76] | yes | asexual -| gametocyte | GTTG(T/C) [76] | [72,76] |
| 24 | PF3D7_1429200 | AP2-O3 | PVX_085085 | PBANKA_1015500 | PY17X_1017000 | yes | yes [65] | yes | zygote -| ookinete | [77] | |
| 25 | PF3D7_1449500 | AP2-O5 | PVX_118015 | PBANKA_1313200 | PY17X_1317000 | refractory | refractory [64] | yes | zygote -| ookinete | ||
| 26 | PF3D7_1456000 | PVX_117665 | PBANKA_1319700 | PY17X_1323500 | yes | yes [64,65] | yes | dispensable | CACACACAC | ||
| 27 | PF3D7_1466400 | AP2-SP/EXP | PVX_117145 | PBANKA_1329800 | PY17X_1334500 | yes | yes [64,65,78] | refractory | oocyst -| sporoblast | TGCATGCA | [31,79] |
| 28 | n.a. | AP2-Q | PVX_080355 | n.a. | n.a. | n.a. | n.a. | n.a. | n.a. | [80,81] |
| AP2 | Name | Function/Phenotype | Interactions | |
|---|---|---|---|---|
| 1 | TGME49_208020 | AP2Ib-1 | ||
| 2 | TGME49_252370 | AP2III-1 | ||
| 3 | TGME49_253380 | AP2III-2 | ||
| 4 | TGME49_299150 | AP2III-3 | ||
| 5 | TGME49_299020 | AP2III-4 | ||
| 6 | TGME49_320700 | AP2IV-1 | ||
| 7 | TGME49_320680 | AP2IV-2 | ||
| 8 | TGME49_318610 | AP2IV-3 | Control of bradyzoite genes [133,134] | |
| 9 | TGME49_318470 | AP2IV-4 | KO leads to expression of bradyzoite specific genes in tachyzoites [136] | |
| 10 | TGME49_211720 | AP2IV-5 | ||
| 11 | TGME49_267460 | AP2IX-1 | ||
| 12 | TGME49_264485 | AP2IX-3 | ||
| 13 | TGME49_288950 | AP2IX-4 | KO leads to reduces tissue cyst formation [137] | |
| 14 | TGME49_289710 | AP2IX-5 | ||
| 15 | TGME49_290180 | AP2IX-6 | ||
| 16 | TGME49_290630 | AP2IX-7 | Implicated in cell cycle progression to S-phase [130] | GCN5b [132] |
| 17 | TGME49_306000 | AP2IX-8 | ||
| 18 | TGME49_306620 | AP2IX-9 | KO enhances cyst formation, alkaline stress response [133,134] | |
| 19 | TGME49_220530 | AP2V-1 | ||
| 20 | TGME49_285895 | AP2V-2 | ||
| 21 | TGME49_240460 | AP2VI-1 | ||
| 22 | TGME49_240900 | AP2VI-2 | ||
| 23 | TGME49_244510 | AP2VI-3 | ||
| 24 | TGME49_280470 | AP2VIIa-1 | ||
| 25 | TGME49_280460 | AP2VIIa-2 | ||
| 26 | TGME49_205650 | AP2VIIa-3 | ||
| 27 | TGME49_203710 | AP2VIIa-4 | ||
| 28 | TGME49_203690 | AP2VIIa-5 | ||
| 29 | TGME49_203050 | AP2VIIa-6 | Implicated in cell cycle progression to S-phase [130] | |
| 30 | TGME49_202490 | AP2VIIa-7 | ||
| 31 | TGME49_282210 | AP2VIIa-8 | ||
| 32 | TGME49_282220 | AP2VIIa-9 | ||
| 33 | TGME49_262420 | APVIIb-1/ADA2-B | ||
| 34 | TGME49_262000 | AP2VIIb-2 | ||
| 35 | TGME49_255220 | AP2VIIb-3 | ||
| 36 | TGME49_229370 | AP2VIII-1 | ||
| 37 | TGME49_233120 | AP2VIII-2 | ||
| 38 | TGME49_273660 | AP2VIII-3 | ||
| 39 | TGME49_272710 | AP2VIII-4 | Implicated in cell cycle progression to S-phase [130] | |
| 40 | TGME49_271200 | AP2VIII-5 | ||
| 41 | TGME49_271030 | AP2VIII-6 | ||
| 42 | TGME49_269010 | AP2VIII-7 | ||
| 43 | TGME49_227900 | AP2X-1 | ||
| 44 | TGME49_225110 | AP2X-2 | ||
| 45 | TGME49_224230 | AP2X-3 | ||
| 46 | TGME49_224050 | AP2X-4 | ||
| 47 | TGME49_237090 | AP2X-5 | Implicated in virulence gene regulation [112] | AP2XI-5 [112] |
| 48 | TGME49_237425 | AP2X-6 | ||
| 49 | TGME49_214840 | AP2X-7 | Implicated in cell cycle progression to S-phase [130] | |
| 50 | TGME49_214960 | AP2X-8 | GCN5b [132] | |
| 51 | TGME49_215150 | AP2X-9 | ||
| 52 | TGME49_215340 | AP2X-10 | ||
| 53 | TGME49_215570 | AP2X-11 | ||
| 54 | TGME49_309410 | AP2XI-1 | ||
| 55 | TGME49_310900 | AP2XI-2 | GCN5b [132] | |
| 56 | TGME49_310950 | AP2XI-3 | ||
| 57 | TGME49_315760 | AP2XI-4 | KO inhibits cyst formation [131,134] | |
| 58 | TGME49_216220 | AP2XI-5 | Implicated in virulence gene regulation [112] | AP2X-5 [112] |
| 59 | TGME49_215895 | AP2 domain-containing protein | ||
| 60 | TGME49_218960 | AP2XII-1 | ||
| 61 | TGME49_217700 | AP2XII-2 | ||
| 62 | TGME49_246660 | AP2XII-3 | ||
| 63 | TGME49_247700 | AP2XII-4 | Implicated in cell cycle progression to S-phase [130] | GCN5b [132] |
| 64 | TGME49_247730 | AP2XII-5 | ||
| 65 | TGME49_249190 | AP2XII-6 | ||
| 66 | TGME49_250800 | AP2XII-8 | ||
| 67 | TGME49_251740 | AP2XII-9 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeninga, M.D.; Quinn, J.E.; Petter, M. ApiAP2 Transcription Factors in Apicomplexan Parasites. Pathogens 2019, 8, 47. https://doi.org/10.3390/pathogens8020047
Jeninga MD, Quinn JE, Petter M. ApiAP2 Transcription Factors in Apicomplexan Parasites. Pathogens. 2019; 8(2):47. https://doi.org/10.3390/pathogens8020047
Chicago/Turabian StyleJeninga, Myriam D., Jennifer E. Quinn, and Michaela Petter. 2019. "ApiAP2 Transcription Factors in Apicomplexan Parasites" Pathogens 8, no. 2: 47. https://doi.org/10.3390/pathogens8020047
APA StyleJeninga, M. D., Quinn, J. E., & Petter, M. (2019). ApiAP2 Transcription Factors in Apicomplexan Parasites. Pathogens, 8(2), 47. https://doi.org/10.3390/pathogens8020047