A Review on Viruses Infecting Taro (Colocasia esculenta (L.) Schott)


 and
and
Abstract
:1. Introduction
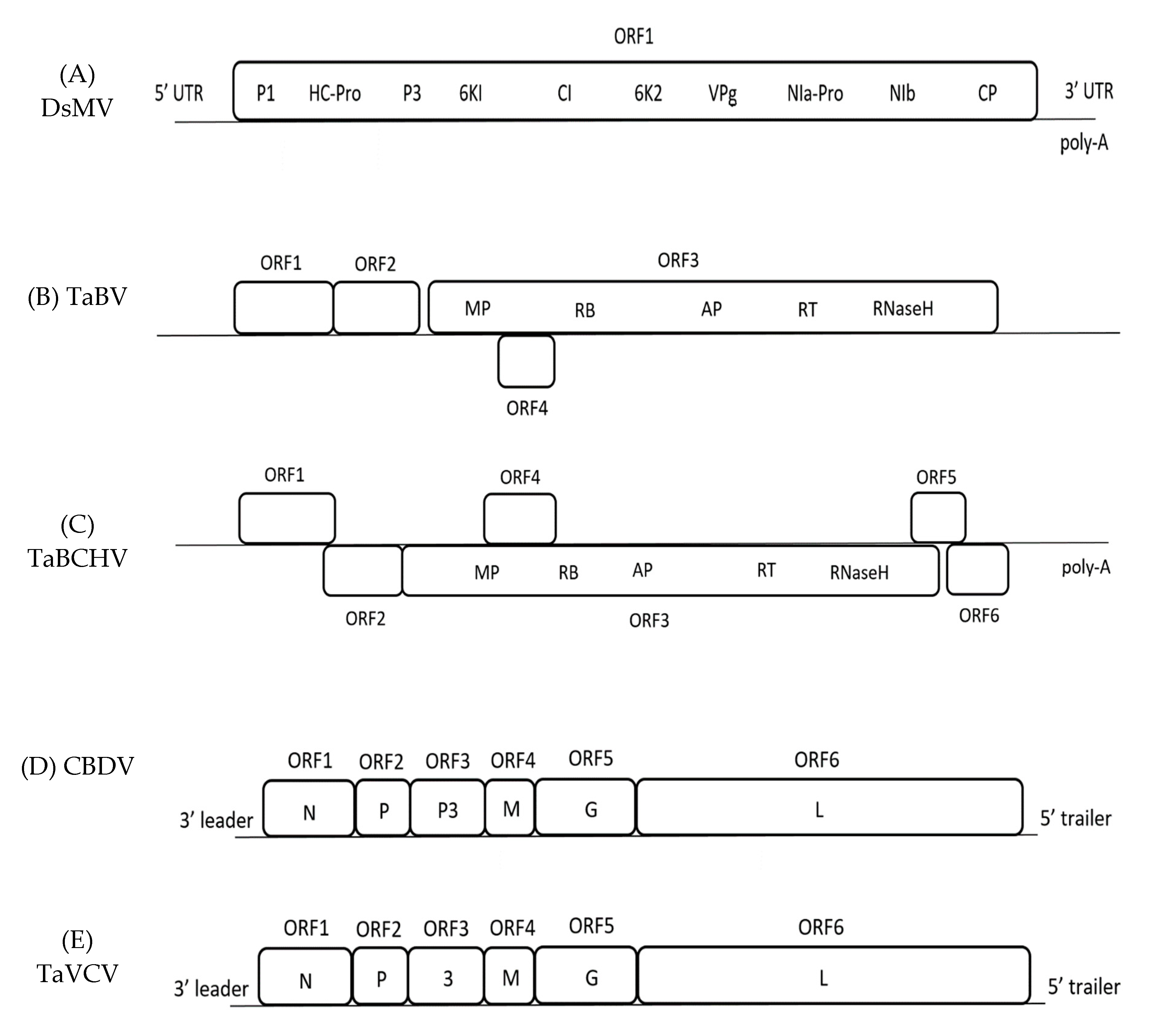
2. Dasheen Mosaic Virus
3. Taro Bacilliform Virus
4. Colocasia Bobone Disease Virus
5. Taro Vein Chlorosis Virus
6. Taro as Host for Plant Viruses
6.1. Diagnostics
6.2. Taro-Virus Interaction
6.3. Taro-Virus Genome Integration
6.4. Taro Diseases
6.5. Taro Viral Disease Management and Challenges
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kreike, C.M.; Van Eck, H.J.; Lebot, V. Genetic diversity of taro, Colocasia esculenta (L.) Schott, in Southeast Asia and the Pacific. Theor. Appl. Genet. 2004, 109, 761–768. [Google Scholar] [CrossRef]
- Lebot, V.; Prana, M.S.; Kreike, N.; Van Heck, H.; Pardales, J.; Okpul, T.; Gendua, T.; Thongjiem, M.; Hue, H.; Viet, N.; et al. Characterisation of taro (Colocasia esculenta (L.) Schott) genetic resources in Southeast Asia and Oceania. Genet. Resour. Crop. Evol. 2004, 51, 381–392. [Google Scholar] [CrossRef]
- Chaïr, H.; Traore, R.E.; Duval, M.F.; Rivallan, R.; Mukherjee, A.; Aboagye, L.M.; Van Rensburg, W.J.; Andrianavalona, V.; Pinheiro de Carvalho, M.A.A.; Saborio, F.; et al. Genetic Diversification and Dispersal of Taro (Colocasia esculenta (L.) Schott). PLoS ONE 2016, 11, 1–19. [Google Scholar] [CrossRef]
- Yang, I.C.; Hafner, G.J.; Revill, P.A.; Dale, J.L.; Harding, R.M. Sequence diversity of South Pacific isolates of Taro bacilliform virus and the development of a PCR-based diagnostic test. Arch. Virol. 2003, 148, 1957–1968. [Google Scholar] [CrossRef]
- Yang, I.C.; Hafner, G.J.; Dale, J.L.; Harding, R.M. Genomic characterisation of taro bacilliform virus. Arch. Virol. 2003, 148, 937–949. [Google Scholar] [CrossRef]
- Pearson, M.N.; Bussell, W.T.; Scheffer, J.J.C. New plant disease record in New Zealand: Dasheen mosaic potyvirus infecting taro (Colocasia esculenta Schott). N. Z. J. Crop. Hortic. Sci. 1998, 26, 69–70. [Google Scholar] [CrossRef]
- Jackson, G.V.H.; Vilsoni, F.; Kumar, J.; Pearson, M.N.; Morton, J.R. Comparison of the growth of in vitro produced pathogen-tested Colocasia taro and field-collected planting material. N. Z. J. Crop. Hortic. Sci. 2001, 29, 171–176. [Google Scholar] [CrossRef]
- Long, M.H.; Ayin, C.; Li, R.; Hu, J.S.; Melzer, M.J. First Report of Taro vein chlorosis virus Infecting Taro (Colocasia esculenta) in the United States. Plant Dis. 2014, 98, 1160. [Google Scholar] [CrossRef]
- Wang, Y.N.; Hu, J.S.; Borth, W.B.; Hamim, I.; Green, J.C.; Melzer, M.J. First report of Taro bacilliform CH Virus (TaBCHV) on Taro (Colocasia esculenta) in Hawaii, USA. Plant Dis. 2017, 101, 1334. [Google Scholar] [CrossRef]
- Tong, P.S. Colocasia esculenta (taro, yam, keladi) as a small farm crop in the Kinta Valley of Malaysia. UTAR Agric. Sci. J. 2016, 2, 49–56. [Google Scholar]
- Sar, S.A. Review of research in Papua New Guinea for sustainable production of taro (Colocasia esculenta). Trop. Agric. 1998, 75, 134–138. [Google Scholar]
- Kidanemariam, D.B.; Macharia, M.W.; Harvey, J.; Holton, T.; Sukal, A.; James, A.P.; Harding, R.M.; Abraham, A.D. First report of Dasheen mosaic virus infecting taro (Colocasia esculenta L.) from Ethiopia. Plant Dis. 2018, 102, 1470. [Google Scholar] [CrossRef]
- Wang, D.; Hamim, I.; Borth, W.B.; Melzer, M.J.; Sun, G.F.; Hu, J.S. First report of Dasheen mosaic virus infecting taro (Colocasia esculenta L.) in Bangladesh. Plant Dis. 2018, 102, 2668. [Google Scholar] [CrossRef]
- Atibalentja, N.; Ilaoa, E.M.; Fiafia, S.T.; Gurr, I.B. Survey of virus diseases of taro, Colocasia esculenta, in American Samoa. In Proceedings of the 2015 APS Annual Meeting, Pasadena, CA, USA, 1–5 August 2015. [Google Scholar]
- Meli, D.; Atibalentja, N. Survey of Virus Diseases of Taro, Colocasia esculenta, on the Aunu’u Island of American Samoa. J. Health Dispar. Res. Pract. 2015, 9, 71. [Google Scholar]
- Jackson, G.V.H.; Gollifer, D.E. Disease and pest problems of Taro (Colocasia esculenta L. Schott) in the British Solomon Islands disease and pest problems of Taro (Coloccasia esculenta L. Schott) in the British Solomon Islands. Pest Artic. News Summ. 1975, 21, 45–53. [Google Scholar] [CrossRef]
- Wang, Y.; Borth, W.B.; Green, J.C.; Hamim, I.; Cao, K.; Hu, J.S.; Melzer, M.J. Genome characterization and distribution of Taro bacilliform CH virus on taro in Hawaii, USA. Eur. J. Plant Pathol. 2018, 150, 1107–1111. [Google Scholar] [CrossRef]
- Sivaprasad, Y.; Bhaskara Reddy, B.V.; Naresh Kumar, C.V.M.; Raja Reddy, K.; Sai Gopal, D.V.R. First report of groundnut bud necrosis Virus infecting Taro (Colocasia esculenta). Australas. Plant Pathol. 2011, 6, 30–32. [Google Scholar] [CrossRef]
- Kazmi, S.A.; Yang, Z.; Hong, N.; Wang, G.; Wang, Y. Characterization by Small RNA Sequencing of Taro Bacilliform CH Virus (TaBCHV), a Novel Badnavirus. PLoS ONE 2015, 10, 1–13. [Google Scholar] [CrossRef]
- Zettler, F.W.; Hartman, R.D. Dasheen Mosaic Virus as a pathogen of cultivated aroids and control of the Virus by tissue culture. Plant Dis. 1987, 71, 958–963. [Google Scholar] [CrossRef]
- Yeturu, S.; Reddy, B.V.B.; Asadhi, S.; Gopal, D.V.R.S. Sequence diversity of the nucleoprotein gene of peanut bud necrosis virus isolates from South India. J. Phytopathol. 2013, 162, 542–547. [Google Scholar] [CrossRef]
- Pearson, M.N.; Jackson, G.V.H.; Saelea, J.; Morar, S.G. Evidence for two rhabdoviruses in taro (Colocasia esculenta) in the Pacific region. Australas. Plant Pathol. 1999, 28, 248–253. [Google Scholar] [CrossRef]
- Sukal, A.C.; Taylor, M.; Tuia, V.S. Viruses and their impact on the utilization of plant genetic resources in the Pacific. Acta Hortic. 2015, 1101, 127–132. [Google Scholar] [CrossRef]
- Pan, S.; Ghosh, S.K. Global status of bacterial, viral and physiogenic diseases of taro Colocasia esculenta L. Environ. Ecol. 1990, 8, 387–392. [Google Scholar]
- Matthews, C.G.; Milne, K.S.; Forster, R.L.S.; Neilson, H.F. Comparison of four potyvirus isolates infecting aroid species. Acta Hortic. 1996, 432, 354–363. [Google Scholar] [CrossRef]
- Wang, Y.F.; Wang, G.P.; Wang, P.; Hong, N. First report of cucumber mosaic virus in Taro plants in China. Am. Phytopathol. Soc. 2014, 98, 574. [Google Scholar]
- Kidanemariam, D.B.; Sukal, A.C.; Abraham, A.D.; Stomeo, F.; Dale, J.L.; James, A.P.; Harding, R.M. Identification and molecular characterisation of Taro bacilliform virus and Taro bacilliform CH virus from East Africa. Plant Pathol. 2018, 67, 1977–1986. [Google Scholar] [CrossRef]
- Kenten, R.H.; Woods, R.D. Viruses of Colocasia esculenta and Xanthosoma saggitifolium. Pest Artic. News Summ. 1973, 19, 38–41. [Google Scholar] [CrossRef]
- Gollifer, D.E.; Jackson, G.V.H.; Dabek, A.J.; Plumb, R.T. Incidence, and effects on yield, of virus diseases of taro (Colocasia esculenta) in the Solomon Islands. Ann. Appl. Biol. 1978, 88, 131–135. [Google Scholar] [CrossRef]
- Li, R.H.; Zettler, F.W.; Hiebert, E.; Purcifull, D.E.; Morales, F.J. Differentiation of Dasheen Mosaic Potyvirus Isolates Based on Variability in the Apparent Size of the Capsid Protein in Western Blots. J. Phytopathol. 1999, 147, 359–364. [Google Scholar] [CrossRef]
- Li, Y.; Xu, C.; Chen, J. Establishment of virus-free Taro (Colocasia esulenta cv. Fenghuayunaitou) by meristem-tip culture combined with thermotherapy. Pakistan J. Plant Pathol. 2002, 1, 40–43. [Google Scholar]
- Abo El-Nil, M.M.; Zettler, F.W.; Hiebert, E. Purification, serology, and some physical properties of dasheen mosaic virus. Phytopathology 1977, 67, 1445–1450. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, B.; Borth, W.B.; Hamim, I.; Green, J.C.; Melzer, M.J.; Hu, J.S. Molecular characterization and distribution of two strains of Dasheen mosaic virus on taro in Hawaii, USA. Plant Dis. 2017, 101, 1980–1989. [Google Scholar] [CrossRef]
- Kamala, S.; Makeshkumar, T.; Sreekumar, J.; Chakrabarti, S.K. Whole transcriptome sequencing of diseased elephant foot yam reveals complete genome sequence of Dasheen mosaic virus. Virol. Rep. 2014, 5, 1–9. [Google Scholar] [CrossRef]
- Chen, J.; Chen, J.; Chen, J.; Adams, M.J. Molecular characterisation of an isolate of Dasheen mosaic virus from Zantedeschia aethiopica in China and comparisons in the genus Potyvirus. Arch. Virol. 2001, 146, 1821–1829. [Google Scholar] [CrossRef]
- Kim, M.K.; Kwon, S.B.; Yoon, J.Y.; Ryu, K.H.; Heo, S.J.; Hong, J.K.; Kim, K.H.; Choi, J.K. Characterization of a Korean Isolate of Dasheen mosaic virus Isolated from Taro in Korea. Plant Pathol. J. 2004, 20, 135–141. [Google Scholar] [CrossRef]
- Hu, J.S.; Meleisea, S.; Wang, M.; Shaarawy, M.A.; Zettler, F.W. Dasheen mosaic potyvirus in Hawaiian taro. Australas. Plant Pathol. 1995, 24, 112–117. [Google Scholar] [CrossRef]
- Babu, B.; Hegde, V.; Makeshkumar, T.; Jeeva, M.L. Detection and Identification of Dasheen mosaic virus Infecting Colocasia esculenta in India. Indian J. Virol. 2011, 22, 59–62. [Google Scholar] [CrossRef]
- Watanabe, S.; Greenwell, A.M.; Bressan, A. Localization, concentration, and transmission efficiency of banana bunchy top virus in four asexual lineages of pentalonia aphids. Viruses 2013, 5, 758–775. [Google Scholar] [CrossRef]
- Revill, P.A.; Jackson, G.V.H.; Hafner, G.J.; Yang, I.; Maino, M.K.; Dowling, M.L.; Devitt, L.C.; Dale, J.L.; Harding, R.M. Incidence and distribution of viruses of Taro (Colocasia esculenta) in Pacific Island countries. Australas. Plant Pathol. 2005, 34, 327–331. [Google Scholar] [CrossRef]
- Gollifer, D.E.; Jackson, G.V.H.; Dabek, A.J.; Plumb, R.T.; May, Y.Y. The occurrence and transmission of Viruses of edible aroids in the Solomon Islands and the Southwest Pacific. Pest Artic. News Summ. 1977, 23, 171–177. [Google Scholar] [CrossRef]
- Dabek, A.J.; Plumb, R.T. Viruslike Particles in the Taro Planthopper, Tarophagus proserpina. J. Invertebr. Pathol. 1975, 26, 271–272. [Google Scholar] [CrossRef]
- Borah, B.K.; Sharma, S.; Kant, R.; Johnson, A.M.A.; Venkata, D.; Saigopal, R.; Dasgupta, I. Bacilliform DNA-containing plant viruses in the tropics: Commonalities within a genetically diverse group. Mol. Plant Pathol. 2013, 14, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.C.; Iommarini, J.P.; Becker, D.K.; Hafner, G.J.; Dale, J.L.; Harding, R.M. A promoter derived from taro bacilliform badnavirus drives strong expression in transgenic banana and tobacco plants. Plant Cell Rep. 2003, 21, 1199–1206. [Google Scholar] [CrossRef] [PubMed]
- Kidanemariam, D.B.; Sukal, A.C.; Crew, K.; Jackson, G.V.H.; Abraham, A.D.; Dale, J.L.; Harding, R.M.; James, A.P. Characterization of an Australian isolate of Taro bacilliform virus and development of an infectious clone. Arch. Virol. 2018, 163, 1677–1681. [Google Scholar] [CrossRef] [PubMed]
- Macanawai, A.R.A.; Ebenebe, A.A.A.; Hunter, D.B.; Devitt, L.C.C.; Hafner, G.J.C.; Harding, R.M.C. Investigations into the seed and mealybug transmission of Taro bacilliform virus. Australas. Plant Pathol. 2005, 34, 73–76. [Google Scholar] [CrossRef]
- Higgins, C.M.; Bejerman, N.; Li, M.; James, A.P.; Dietzgen, R.G.; Pearson, M.N.; Revill, P.A.; Harding, R.M. Complete genome sequence of Colocasia bobone disease-associated virus, a putative cytorhabdovirus infecting taro. Arch. Virol. 2016, 161, 745–748. [Google Scholar] [CrossRef]
- Revill, P.; Trinh, X.; Dale, J.; Harding, R. Taro vein chlorosis virus: Characterization and variability of a new nucleorhabdovirus. J. Gen. Virol. 2005, 86, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Lamprecht, R.L.; Pietersen, G.; Kasdorf, G.G.F.; Nel, L.H. Characterisation of a proposed Nucleorhabdovirus new to South Africa. Eur. J. Plant Pathol. 2009, 123, 105–110. [Google Scholar] [CrossRef]
- Liebrecht, M.; Winter, S. Complete Genome of Dasheen Mosaic Virus Isolate CTCRI-II-14 Isolated from Taro (Colocasia esculenta L. (Schott). 2015. Available online: https://www.ncbi.nlm.nih.gov/nuccore/KT026108.1 (accessed on 19 December 2018).
- Liu, Z.Q.; Li, X.D. Determination of the Complete Genomic Sequences of Dasheen Mosaic Virus. 2012. Available online: https://www.ncbi.nlm.nih.gov/nuccore/JX083210.1 (accessed on 19 December 2018).
- Kamala, S.; Makeshkumar, T.; Sreekumar, J.; Chakrabarti, S.K. Dasheen Mosaic Virus RNA for Polyprotein Gene, Isolate T10. 2014. Available online: https://www.ncbi.nlm.nih.gov/nuccore/KJ786965.1 (accessed on 19 December 2018).
- Manikonda, P.; Puthankalam, S.K.; Hema, M.; Pothur, S. Development of duplex RT-PCR for detection of Konjac mosaic virus and Spathiphyllum chlorotic vein banding virus in taro and peace lily. Indian J. Biotechnol. 2015, 14, 120–122. [Google Scholar]
- Harding, R.M.; Revill, P.A.; Hafner, G.J.; Yang, I.; Maino, M.K.; Devitt, L.C.; Dowling, M.L.; Dale, J.L. Characterisation of Taro Viruses and the Development of Diagnostic Tests. In Proceedings of the Third Taro Symposium, Tanoa International Hotel, Nadi, Fiji Islands, 21–23 May 2003. [Google Scholar]
- Atibalentja, N.; Fiafia, S.T.; Gosai, R.C.; Melzer, M.J. First Report of Taro vein chlorosis virus on Taro (Colocasia esculenta) in the U.S. Territory of American Samoa. Plant Dis. 2018, 102, 828. [Google Scholar] [CrossRef]
- Qiao, W.; Medina, V.; Kuo, Y.-W.; Falk, B.W. A Distinct, non-virion plant virus movement protein encoded by a Crinivirus essential for systemic infection. mBio 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Xiong, Y.; Eickbush, T.H. Origin and evolution of retroelements based upon their reverse transcriptase sequences. EMBO J. 1990, 9, 3353–3362. [Google Scholar] [CrossRef]
- Ballut, L.; Drucker, M.; Pugnière, M.; Cambon, F.; Blanc, S.; Roquet, F.; Candresse, T.; Schmid, H.P.; Nicolas, P.; Le Gall, O.; et al. HcPro, a multifunctional protein encoded by a plant RNA virus, targets the 20S proteasome and affects its enzymic activities. J. Gen. Virol. 2005, 86, 2595–2603. [Google Scholar] [CrossRef] [Green Version]
- Bhat, A.I.; Hohn, T.; Selvarajan, R. Badnaviruses: The current global scenario. Viruses 2016, 8, 177. [Google Scholar] [CrossRef]
- Geering, A.D.W.; Olszewski, N.E.; Dahal, G.; Thomas, J.E.; Lockhart, B.E.L. Analysis of the distribution and structure of integrated Banana streak virus DNA in a range of Musa cultivars. Mol. Plant Pathol. 2001, 2, 207–213. [Google Scholar] [CrossRef]
- Ivancic, A.; Liloqula, R.; Levelaz, H.; Saelea, J. Effect of alomae-bobone virus complex on young Taro seedlings and other aroid species in controlled conditions. In Proceedings of the Sustainable Taro Culture for the Pacific Conference, Honolulu, HI, USA, 24–25 September 1992; University of Hawaii: Honolulu, HI, USA, 1993; pp. 25–31. [Google Scholar]
- Ivancic, A.; Liwqula, R.; Levela, H.; Saelea, J.; Wagatora, D. Genetic resistance to alomae-bobone virus complex the lethal disease of Taro (Colocasia esculenta (L.) Schott). In Proceedings of the Sustainable Taro Culture for the Pacific Conference, Honolulu, HI, USA, 24–25 September 1992; University of Hawaii: Honolulu, HI, USA, 1993; pp. 48–54. [Google Scholar]
- Zettler, F.W.; Jackson, G.V.H.; Frison, E.A. Fao/Ibpgr Technical Guidelines For The Safe Movement of Edible Aroid Germplasm; Bioversity International: Rome, Italy, 1989. [Google Scholar]
- He, X.; Miyasaka, S.C.; Fitch, M.M.M.; Zhu, Y.J. Taro (Colocasia esculenta (L.) Schott). In Methods in Molecular Biology; Wang, K., Ed.; Springer Science+Business Media: New York, NY, USA, 2015; Volume 1224, pp. 97–108. ISBN 9781493916580. [Google Scholar]
- Li, R.H.; Zettler, F.W.; Purcifull, D.E.; Hiebert, E. The nucleotide sequence of the 3 -terminal region of Dasheen mosaic virus (Caladium isolate) and expression of its coat protein in Escherichia coli for antiserum production. Arch. Virol. 1998, 143, 2461–2469. [Google Scholar] [CrossRef]
- Zhao, J.-P.; Jiang, X.-M.; Bai, X.-F.; Zhang, P.; Yong, J.-P. Several Physiological Factors Influencing Virus-free and Rapid Propagation in Shoot-tip Culture of Colocasia esculenta L. Schott. Plant Phisiol. J. 2009, 45, 133–136. [Google Scholar]
- Banjaw, D.T. Review of Taro (Colocasia esculenta) Genetics and Breeding. J. Hortic. 2017, 04, 1–4. [Google Scholar] [CrossRef]
- Davis, R.I.; Lomavatu-Fong, M.F.; Mcmichael, L.A.; Ruabete, T.K.; Kumar, S.; Turaganivalu, U. Cucumber mosaic virus infection of kava (Piper methysticum) and implications for cultural control of kava dieback disease. Australas. Plant Pathol. 2005, 34, 377–384. [Google Scholar] [CrossRef]
- Matsumoto, B.M.; Nishida, T. Predator-Prey Investigations on the Taro Leafhopper and Its Egg Predator; Technical bulletin No. 64; Hawaii Agricultural Experiment Station: Honolulu, HI, USA, December 1966. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Name (Taxon) | Isolate (Length) | Source | Accession Number | Reference |
|---|---|---|---|---|
| DsMV (Potyvirus) (+)ssRNA virus | DsMV (10038 bp) | China | NC_003537.1 | [35] |
| DsMV strain I (10002 bp) | Hawaii, USA | KY242358.1 | [33] | |
| DsMV strain II (10019 bp) | Hawaii, USA | KY242359.1 | [33] | |
| DsMV CTCRI-II-14 (10004 bp) | India | KT026108.1 | [50] | |
| DsMV SdP (10030 bp) | China | JX083210.1 | [51] | |
| DsMV T10 (10024 bp) | India | KJ786965.1 | [52] | |
| TaBV (Badnavirus) ssDNA virus | TaBV (7458 bp) | Papua New Guinea | NC_004450.1 AF357836.1 | [5] |
| TaBV Aus7 (7494 bp) | Australia | MG017318.1 | [45] | |
| TaBV Ke52 (7805 bp) | East Africa (Kenya) | MG017321.1 | [27] | |
| TaBV Tz17 (7803 bp) | East Africa (Tanzania) | MG017322.1 | [27] | |
| TaBV Tz24 (7798 bp) | East Africa (Tanzania) | MG833013.1 | [27] | |
| TaBV Ug75 (7796 bp) | East Africa (Uganda) | MG017323.1 | [27] | |
| TaBCHV (Badnavirus) ssDNA virus | TaBCHV-1 (7641 bp) | China | NC_026819.1 | [19] |
| TaBCHV-2 (7641 bp) | China | KP710177.1 | [19] | |
| TaBCHV Et17 (7610 bp) | East Africa (Ethiopia) | MG017324.1 | [27] | |
| TaBCHV Ke43 (7647 bp) | East Africa (Kenya) | MG017325.1 | [27] | |
| TaBCHV Tz27 (7389 bp) | East Africa (Tanzania) | MG833014.1 | [27] | |
| TaBCHV Tz36 (7654 bp) | East Africa (Tanzania) | MG017326.1 | [27] | |
| TaBCHV Ug10 (7643 bp) | East Africa (Uganda) | MG017327.1 | [17] | |
| TaBCHV isolate Hawaii (7634 bp) | Hawaii, USA | KY359389.1 | [17] | |
| CBDV (Cytorhabdovirus) (-)ssRNA virus | CBDV strain SI (12193 bp) | Solomon Islands | NC_034551.1 KT381973.1 | [47] |
| TaVCV (Nucleorhabdovirus) (-)ssRNA virus | TaVCV (12020 bp) | Fiji | NC_006942.1 AY674964.1 | [48] |
| Virus | Detection Method | Remarks | Reference |
|---|---|---|---|
| DsMV | SDS-PAGE | Estimation on the relative size of CP of DsMV compared to other viruses could be done. | [32] |
| Western blot | Characterising the CP of DsMV could be done for comparative studies among isolates and potyviruses. | [30] | |
| ELISA | Relatively sensitive & easy to use for routine virus detection. | [37] | |
| RT-PCR | This method was developed after nucleic acid sequence was completed. This allow the detection of wide range of isolates. | [31,33,35,40,54] | |
| TaBV | PCR | Primers designed are usually based on the putative coding regions of TaBV, with the most widely mentioned in literatures being the primers BadnaFP & BadnaRP responsible for RT & RNaseH-coding regions. | [4,5,27,45] |
| TaBCHV | RT-PCR | Developed to examine the occurrence & distribution of TaBCHV on taro. | [9,17,19] |
| Dot-blot hybridisation assay | Used in confirmation test alongside RT-PCR in [9]. | [9] | |
| PCR | In [27], the method is described alongside rolling circle amplification (RCA) to identify the virus. | [27] | |
| CBDV | Immuno-osphoresis | A part of serological study of rhabdoviruses detection in multiple taro samples in the Pacific Islands. | [22] |
| RT-PCR | Method used for detection, completing genome sequence, and recording distribution of the viruses. | [40,47] | |
| TaVCV | ELISA | A part of serological study of rhabdoviruses detection in multiple taro samples in the Pacific Islands. | [22] |
| RT-PCR | For examining the distribution of viruses in the Pacific Islands. | [54,55] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yusop, M.S.M.; Saad, M.F.M.; Talip, N.; Baharum, S.N.; Bunawan, H. A Review on Viruses Infecting Taro (Colocasia esculenta (L.) Schott). Pathogens 2019, 8, 56. https://doi.org/10.3390/pathogens8020056
Yusop MSM, Saad MFM, Talip N, Baharum SN, Bunawan H. A Review on Viruses Infecting Taro (Colocasia esculenta (L.) Schott). Pathogens. 2019; 8(2):56. https://doi.org/10.3390/pathogens8020056
Chicago/Turabian StyleYusop, Mohd Shakir Mohamad, Mohd Faiz Mat Saad, Noraini Talip, Syarul Nataqain Baharum, and Hamidun Bunawan. 2019. "A Review on Viruses Infecting Taro (Colocasia esculenta (L.) Schott)" Pathogens 8, no. 2: 56. https://doi.org/10.3390/pathogens8020056
APA StyleYusop, M. S. M., Saad, M. F. M., Talip, N., Baharum, S. N., & Bunawan, H. (2019). A Review on Viruses Infecting Taro (Colocasia esculenta (L.) Schott). Pathogens, 8(2), 56. https://doi.org/10.3390/pathogens8020056