Withanolides from Withania aristata as Antikinetoplastid Agents through Induction of Programmed Cell Death


 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
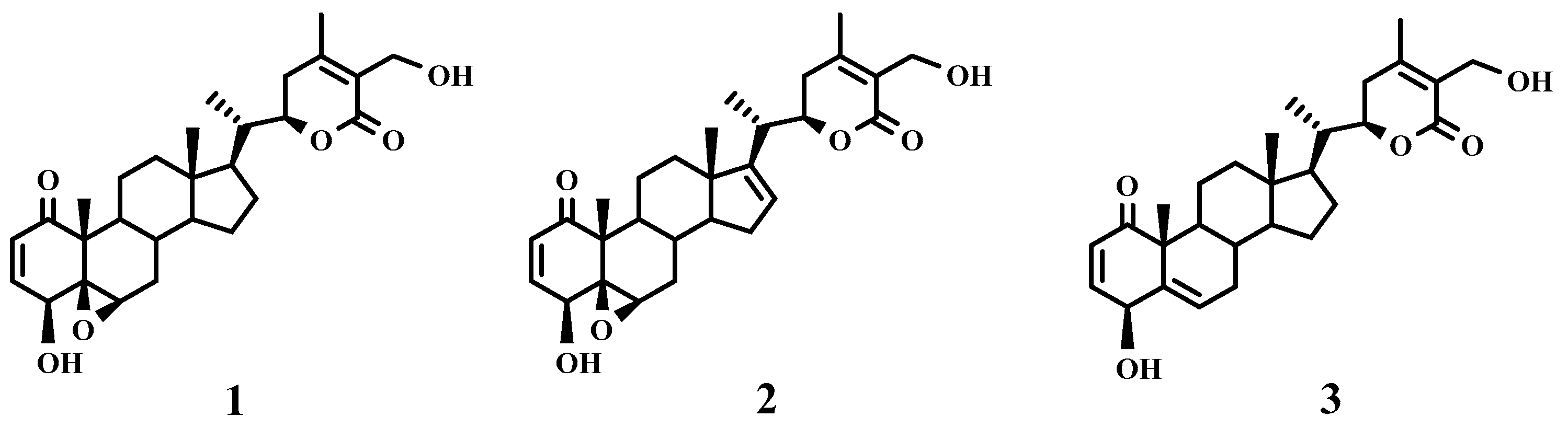
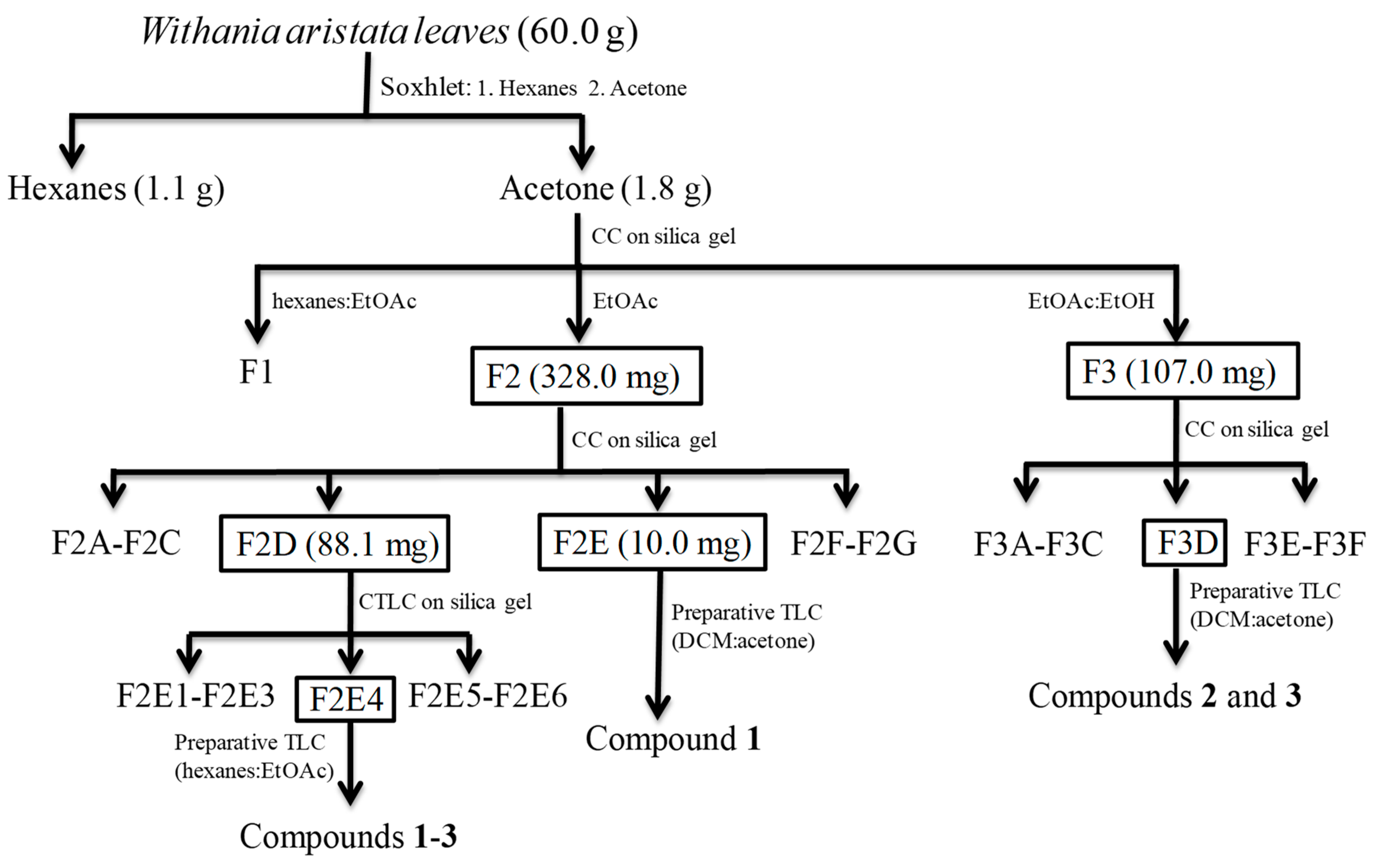
2.1. Bioassay-Guided Fractionation
2.2. Mechanisms of Cell Death
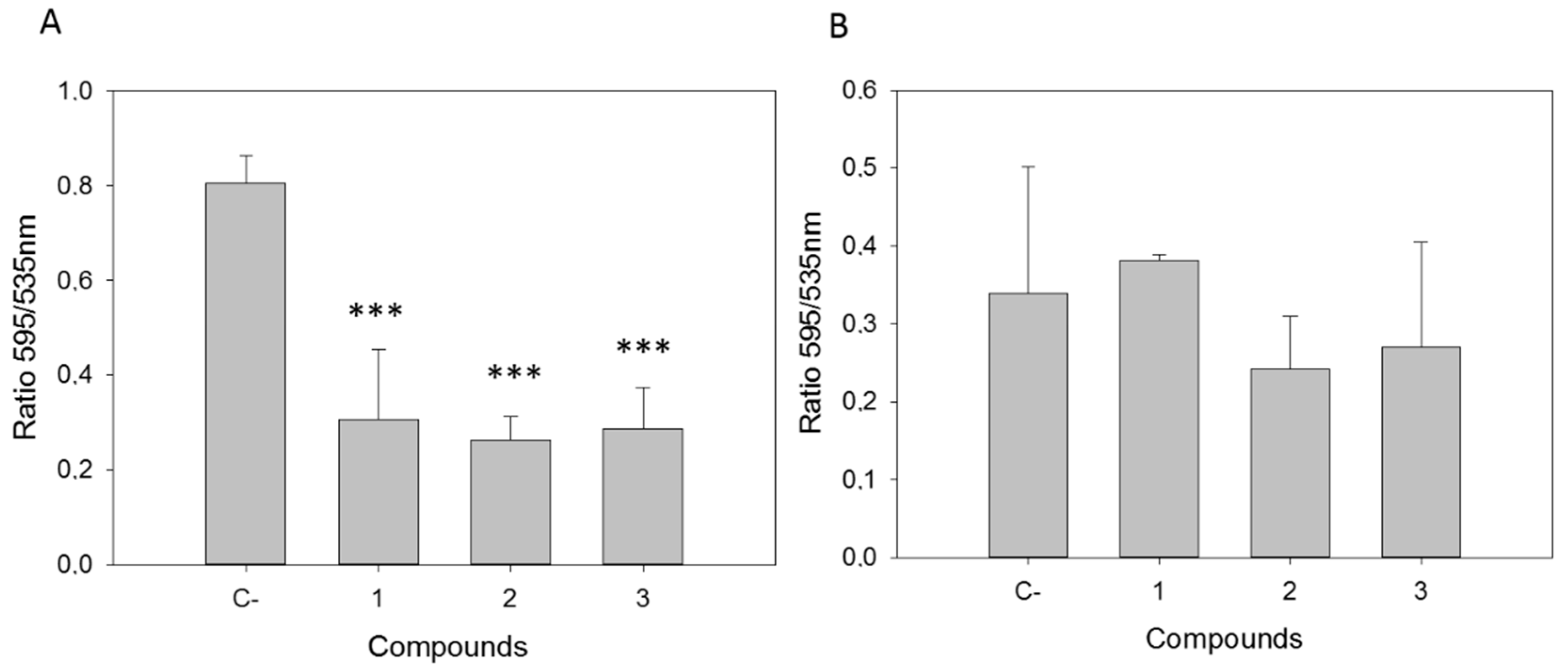
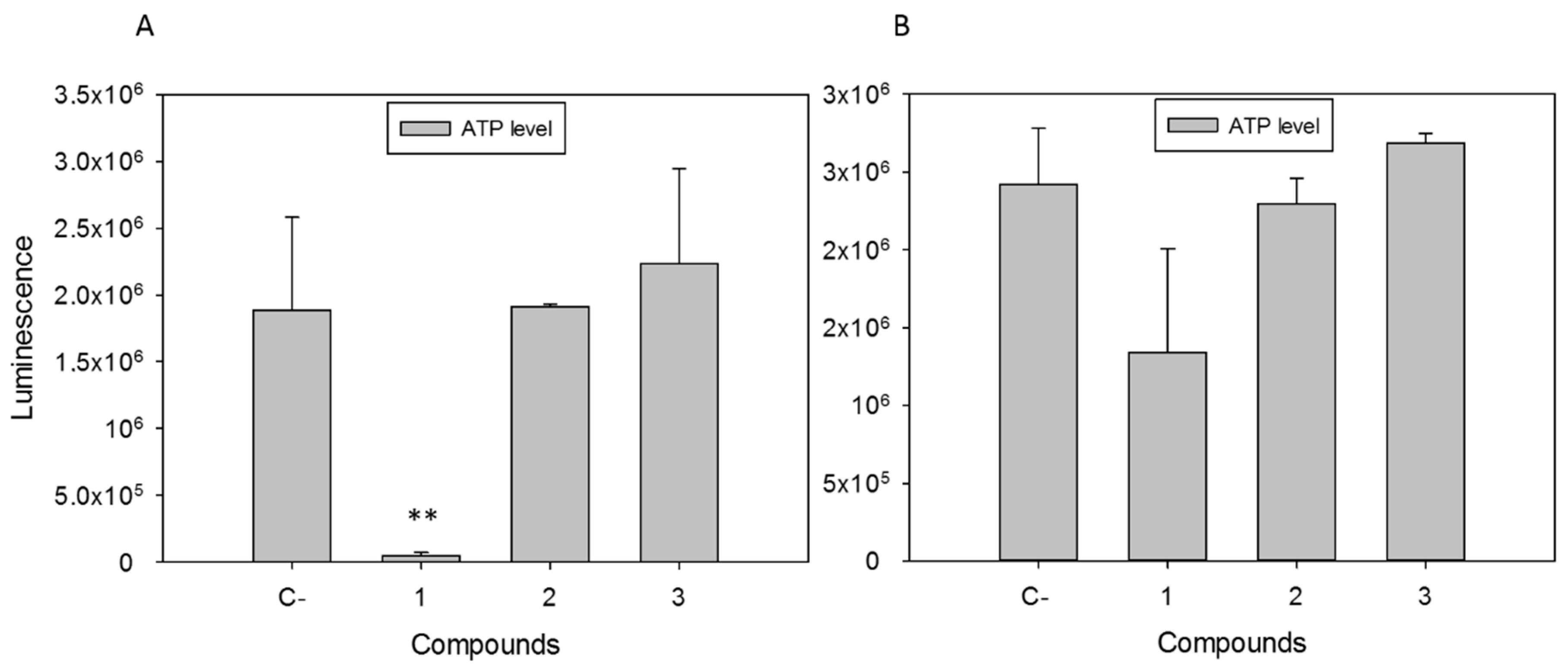
2.2.1. Withanolides induced Mitochondrial Damage in L. amazonensis
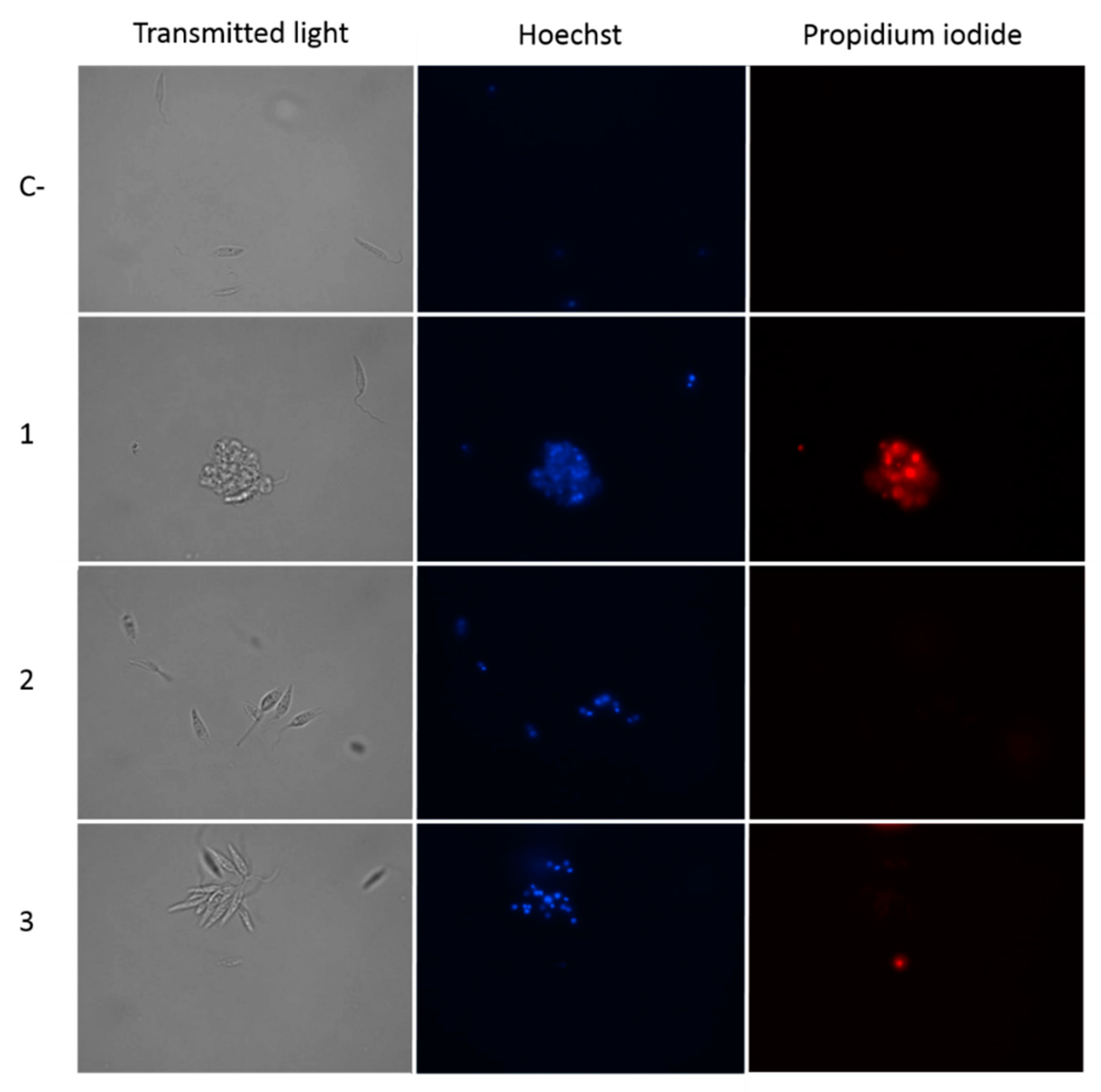
2.2.2. Withanolides Caused Chromatin Condensation in Treated Cells
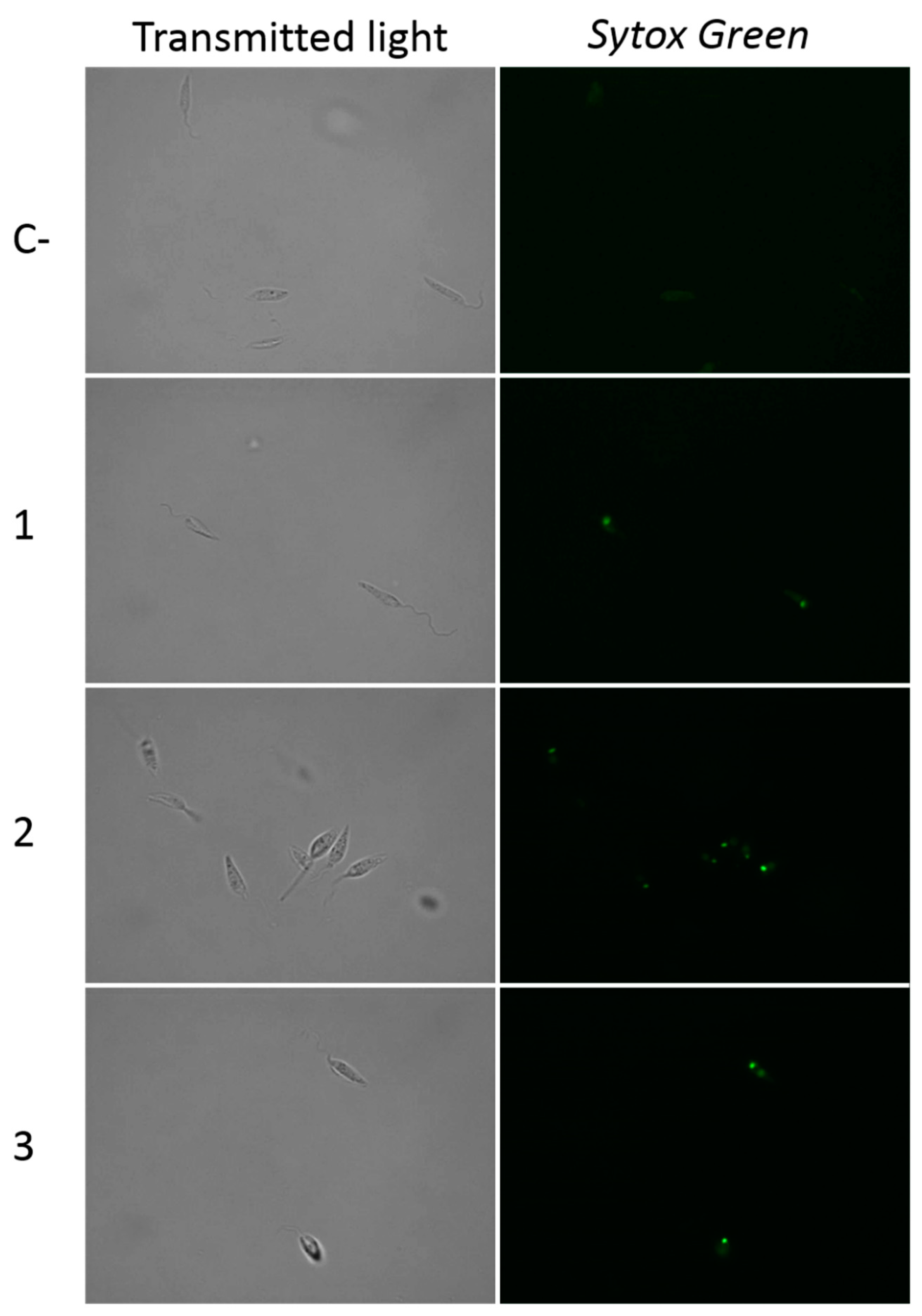
2.2.3. Withanolides Induce Cytoplasmic Membrane Permeability in Treated Cells.
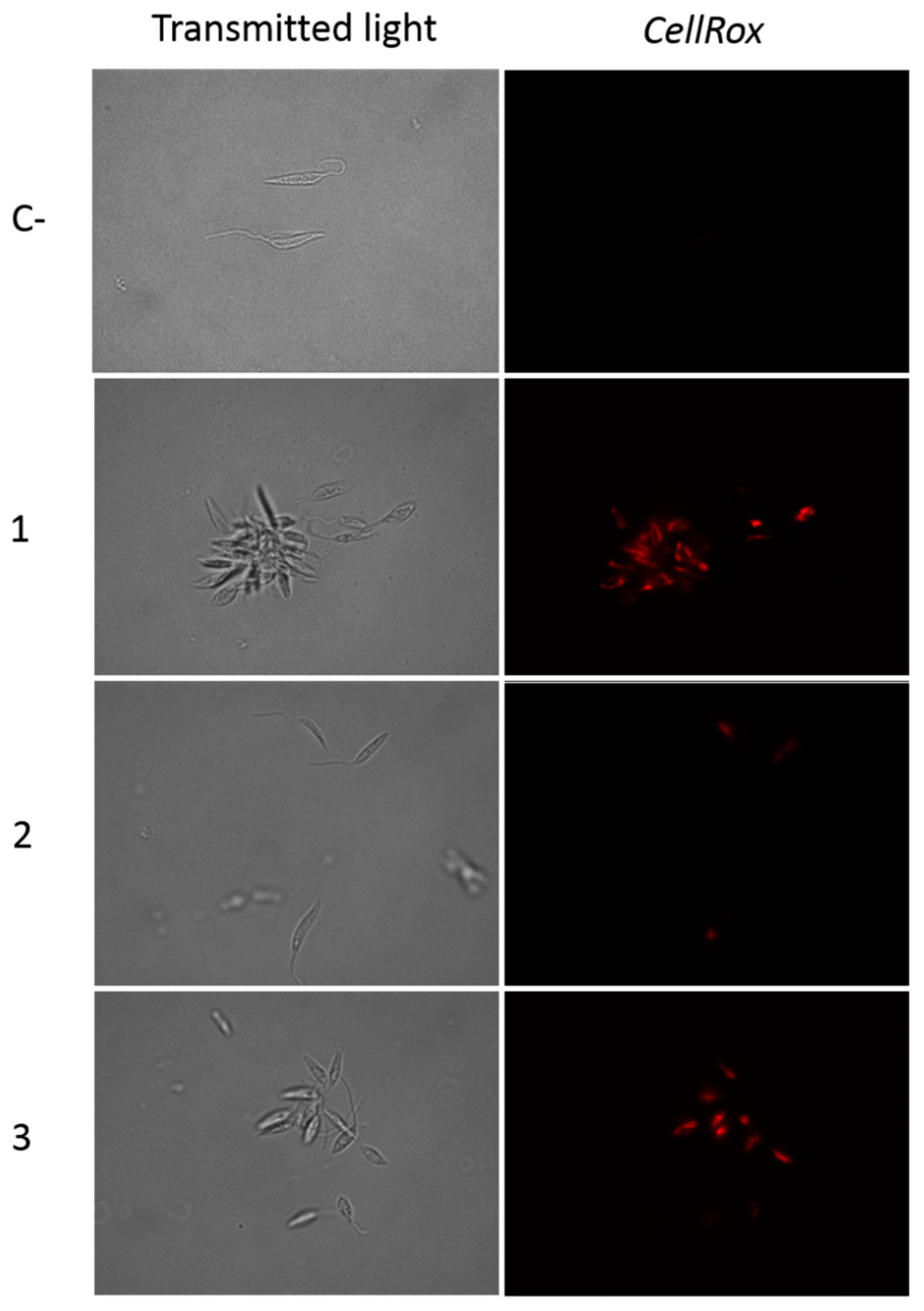
2.2.4. Withanolides Induce Oxidative Stress in L. amazonensis
3. Material and Methods
3.1. General Procedures
3.2. Plant Material
3.3. Extraction, Bioassay-Guided Fractionation and Isolation
3.4. Cultures
3.5. Leishmanicidal and Trypanocidal Assays Cytotoxic Effect Evaluation
3.5.1. Leishmanicidal Activity Assay
3.5.2. Trypanocidal Capacity Assay
3.5.3. Cytotoxicity Assay
3.6. Mechanisms of Cell Death
3.6.1. Mitochondrial Membrane Potential evaluation
3.6.2. Measurement of ATP
3.6.3. Chromatin Condensation Determination
3.6.4. Plasma Membrane Permeability
3.6.5. Oxidative Stress
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- WHO. Neglected Tropical Diseases. 2019. Available online: http://www.who.int/neglected_diseases/diseases/en/ (accessed on 19 April 2019).
- Rao, S.P.S.; Barrett, M.P.; Dranoff, G.; Faraday, C.J.; Gimpelewicz, C.R.; Hailu, A.; Jones, C.L.; Kelly, J.M.; Lazdins-Helds, J.K.; Mäser, P.; et al. Drug discovery for kinetoplastid diseases: Future directions. ACS Infect. Dis. 2019, 5, 152–157. [Google Scholar] [CrossRef]
- Bustos, P.L.; Milduberger, N.; Volta, B.J.; Perrone, A.E.; Laucella, S.A.; Bua, J. Trypanosoma cruzi infection at the maternal-fetal interface: Implications of parasite load in the congenital transmission and challenges in the diagnosis of infected newborns. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Nagle, A.S.; Khare, S.; Kumar, A.B.; Supek, F.; Buchynskyy, A.; Mathison, C.J.N.; Chennamaneni, N.K.; Pendem, N.; Buckner, F.S.; Gelb, M.H.; et al. Recent developments in drug discovery for leishmaniasis and human african trypanosomiasis. Chem. Rev. 2014, 114, 11305–11347. [Google Scholar] [CrossRef] [PubMed]
- Cheuka, P.M.; Mayoka, G.; Mutai, P.; Chibale, K. The role of natural products in drug discovery and development against neglected tropical diseases. Molecules 2017, 22, 58. [Google Scholar] [CrossRef] [PubMed]
- Eich, E. Solanaceae and Convolvulaceae: Secondary Metabolites: Biosynthesis, Chemotaxonomy, Biological and Economic Significance; Springer: Berlin/Heidelberg, Germany, 2008; pp. 16–21. [Google Scholar]
- Chen, L.X.; He, H.; Qiu, F. Natural withanolides: An overview. Nat. Prod. Rep. 2011, 28, 705–740. [Google Scholar] [CrossRef] [PubMed]
- Cruz, S.J. Más de 100 Plantas Medicinales. Medicina Popular Canaria Monografías; Obra Social de La Caja de Canarias; Imprenta Pérez Galdós: Las Palmas de Gran Canaria, España, 2007. [Google Scholar]
- Llanos, G.G.; Araujo, L.M.; Jiménez, I.A.; Moujir, L.M.; Vázquez, J.T.; Bazzocchi, I.L. Withanolides from Withania aristata and their cytotoxic activity. Steroids 2010, 7, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Llanos, G.G.; Araujo, L.M.; Jiménez, I.A.; Moujir, L.M.; Bazzocchi, I.L. Withaferin A-related steroids from Withania aristata exhibit potent antiproliferative activity by inducing apoptosis in human tumor cells. Eur. J. Med. Chem. 2012, 54, 499–511. [Google Scholar] [CrossRef]
- Benjumea, D.; Martín-Herrera, D.; Abdala, S.; Gutiérrez-Luis, J.; Quiñones, W.; Cardona, D.; Torres, F.; Echeverri, F. Withanolides from Whitania aristata and their diuretic activity. J. Ethnopharmacol. 2009, 123, 351–355. [Google Scholar] [CrossRef]
- Chirumamilla, C.S.; Perez-Novo, C.; Van, O.X.; Berghe, W.V. Molecular insights into cancer therapeutic effects of the dietary medicinal phytochemical withaferin A. Proc. Nutr. Soc. 2017, 76, 96–105. [Google Scholar] [CrossRef]
- Jiménez-Ruiz, A.; Alzate, J.F.; MacLeod, E.T.; Kurt, L.C.G.; Fasel, N.; Hurd, H. Apoptotic markers in protozoan parasites. Parasites Vectors 2010, 3, 104. [Google Scholar] [CrossRef]
- Basmaciyan, L.; Azas, N.; Casanova, M. A potential acetyltransferase involved in Leishmania major metacaspase-dependent cell death. Parasites Vectors 2019, 12, 266. [Google Scholar] [CrossRef] [PubMed]
- López-Arencibia, A.; García-Velázquez, D.; Martín-Navarro, C.M.; Sifaoui, I.; Reyes-Batlle, M.; Lorenzo-Morales, J.; Piñero, J. In Vitro Activities of Hexaazatrinaphthylenes against Leishmania spp. Antimicrob. Agents Chemother. 2015, 59, 2867–2874. [Google Scholar] [CrossRef] [PubMed]
- Vandana, D.R.; Tiwari, R.; Katyal, A.; Pandey, K.C. Metacaspases: Potential Drug Target Against Protozoan Parasites. Front Pharmacol. 2019, 10, 790. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, S.; Dayakar, A.; Veronica, J.; Sundar, S.; Maurya, R. An in vitro study of apoptotic like death in Leishmania donovani promastigotes by withanolides. Parasitol. Int. 2013, 62, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, S.; Veronica, J.; Sundar, S.; Maurya, R. Alcoholic Fractions F5 and F6 from Withania somnifera Leaves Show a Potent Antileishmanial and Immunomodulatory Activities to Control Experimental Visceral Leishmaniasis. Front. Med. (Lausanne) 2017, 4, 55. [Google Scholar] [CrossRef]
- Kuroyanagi, M.; Murata, M.; Nakane, T.; Shirota, O.; Sekita, S.; Fuchino, H.; Shinwari, Z.K. Leishmanicidal active withanolides from a Pakistani medicinal plant, Withania coagulans. Chem. Pharm. Bull. (Tokyo) 2012, 60, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Nasser, A.; Al-Sokari, S.; Mothana, R.; Hamed, M.; Wagih, M.; Cos, P.; Maes, L. In vitro antiprotozoal activity of five plant extracts from Albaha region. World J. Pharm. Res. 2016, 5, 338–346. [Google Scholar]
- Lee, I.; Choi, B.Y. Withaferin-A a natural anticancer agent with pleitropic mechanisms of action. Int. J. Mol. Sci. 2016, 17, 290–312. [Google Scholar] [CrossRef]
- Llanos, G.G.; Araujo, L.M.; Jiménez, I.A.; Moujir, L.M.; Rodríguez, J.; Jiménez, C.; Bazzocchi, I.L. Structure-based design, synthesis, and biological evaluation of withaferin A-analogues as potent apoptotic inducers. Eur. J. Med. Chem. 2017, 140, 52–64. [Google Scholar] [CrossRef]
- Perestelo, N.R.; Llanos, G.G.; Reyes, C.P.; Amesty, A.; Sooda, K.; Afshinjavid, S.; Jiménez, I.A.; Javid, F.; Bazzocchi, I.L. Expanding the Chemical Space of Withaferin A by Incorporating Silicon to Improve Its Clinical Potential on Human Ovarian Carcinoma Cells. J. Med. Chem. 2019, 62, 4571–4585. [Google Scholar] [CrossRef]
- Sen, N.; Banerjee, B.; Das, B.B.; Ganguly, A.; Sen, T.; Pramanik, S.; Mukhopadhyay, S.; Majumder, H.K. Apoptosis is induced in leishmanial cells by a novel protein kinase inhibitor withaferin A and is facilitated by apoptotic topoisomerase I-DNA complex. Cell Death Differ. 2007, 14, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Salmerón, M.L.; Quintana-Aguiar, J.; De La Rosa, J.V.; López-Blanco, F.; Castrillo, A.; Gallardo, G.; Tabraue, C. Phenalenone-photodynamic therapy induces apoptosis on human tumor cells mediated by caspase-8 and p38-MAPK activation. Mol. Carcinog. 2018, 57, 1525–1539. [Google Scholar] [CrossRef] [PubMed]
- Sifaoui, I.; López-Arencibia, A.; Martín-Navarro, C.M.; Chammem, N.; Reyes-Batlle, M.; Mejri, M.; Lorenzo-Morales, J.; Abderabba, M.; Piñero, J.E. Activity of olive leaf extracts against the promastigote stage of Leishmania species and their correlation with the antioxidant activity. Exp. Parasitol. 2014, 141, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.K.; Sahu, R.; Walker, L.A.; Tekwani, B.L. A parasite rescue and transformation assay for antileishmanial screening against intracellular Leishmania donovani amastigotes in THP1 human acute monocytic leukemia cell line. J. Vis. Exp. 2012, 30, 4054. [Google Scholar] [CrossRef] [PubMed]
- Fadel, H.; Sifaoui, I.; López-Arencibia, A.; Reyes-Batlle, M.; Hajaji, S.; Chiboub, O.; Jiménez, I.A.; Bazzocchi, I.L.; Lorenzo-Morales, J.; Benayache, S.; et al. Assessment of the antiprotozoal activity of Pulicaria inuloides extracts, an Algerian medicinal plant: Leishmanicidal bioguided fractionation. Parasitol. Res. 2018, 117, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Codonho, B.S.; Costa, S.S.; Peloso, E.F.; Joazeiro, P.P.; Gadelha, F.R.; Giorgio, S. HSP70 of Leishmania amazonensis alters resistance to different stresses and mitochondrial bioenergetics. Memórias do Instituto Oswaldo Cruz 2016, 111, 460–468. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract/Fractions | L. amazonensis IC50 b (µg/mL) | L. donovani IC50 (µg/mL) | T. cruzi IC50 (µg/mL) | Murine Macrophages CC50 c (µg/mL) |
|---|---|---|---|---|
| Acetone extract | 2.87 | 20.25 | 12.78 | 42.63 |
| Fraction 1 | 12.27 | 19.12 | 28.91 | 81.39 |
| Fraction 2 | 1.02 | 12.73 | 1.12 | 11.22 |
| F2A | 1.37 | >50 | 11.24 | 13.79 |
| F2B | 1.05 | 3.06 | 1.93 | 8.02 |
| F2C | 1.40 | 4.21 | 3.14 | 10.23 |
| F2D | 0.36 | 1.85 | 1.73 | 7.65 |
| F2E | 0.19 | 2.70 | 2.02 | 13.90 |
| F2F | 2.11 | >50 | 5.74 | 21.12 |
| Fraction 3 | 4.53 | 22.28 | 3.63 | 46.11 |
| F3A | 2.51 | 9.03 | 8.42 | 22.90 |
| F3B | 2.48 | 5.96 | 6.04 | 23.78 |
| F3C | 2.35 | >50 | 5.56 | 16.08 |
| F3D | 1.01 | 3.67 | 3.59 | 9.56 |
| F3E | 3.42 | >50 | 7.05 | 21.57 |
| F3F | 19.52 | >50 | >50 | 43.98 |
| Miltefosine d | 2.64 | 1.35 | 29.42 | |
| Benznidazole d | 1.81 | 104.1 |
| Cp | L. amazonensis IC50 (µM ± SD) a | SI | L. donovani IC50 (µM ± SD) | SI | T. cruzi IC50 (µM ± SD) | SI c | Murine Macrophages CC50 (µM) b |
|---|---|---|---|---|---|---|---|
| 1 | 0.83 ± 0.07 | 14.36 | 13.22 ± 1.16 | 0.91 | 1.02 ± 0.10 | 11.69 | 11.92 ± 1.08 |
| 2 | 27.37 ± 1.03 | 3.12 | >50 | 14.68 ± 0.43 | 5.82 | 85.47 ± 6.00 | |
| 3 | 2.82 ± 0.07 | 4.35 | 20.88 ± 1.33 | 0.59 | 2.41 ± 0.11 | 5.09 | 12.26 ± 1.16 |
| M d | 6.48 ± 0.10 | 11.14 | 3.31 ± 0.11 | 21.81 | 72.18 ± 1.25 | ||
| B d | 6.95 ± 0.50 | 57.54 | 399.91 ± 1.04 |
| Compound | L. amazonensis IC50 (µM ± SD) | SI a |
|---|---|---|
| 1 | 0.055 ± 0.009 | 216.73 |
| 2 | 0.663 ± 0.075 | 128.91 |
| 3 | 0.209 ± 0.028 | 58.66 |
| Miltefosine | 3.12 ± 0.12 | 23.13 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Arencibia, A.; San Nicolás-Hernández, D.; Bethencourt-Estrella, C.J.; Sifaoui, I.; Reyes-Batlle, M.; Rodríguez-Expósito, R.L.; Rizo-Liendo, A.; Lorenzo-Morales, J.; Bazzocchi, I.L.; Piñero, J.E.; et al. Withanolides from Withania aristata as Antikinetoplastid Agents through Induction of Programmed Cell Death. Pathogens 2019, 8, 172. https://doi.org/10.3390/pathogens8040172
López-Arencibia A, San Nicolás-Hernández D, Bethencourt-Estrella CJ, Sifaoui I, Reyes-Batlle M, Rodríguez-Expósito RL, Rizo-Liendo A, Lorenzo-Morales J, Bazzocchi IL, Piñero JE, et al. Withanolides from Withania aristata as Antikinetoplastid Agents through Induction of Programmed Cell Death. Pathogens. 2019; 8(4):172. https://doi.org/10.3390/pathogens8040172
Chicago/Turabian StyleLópez-Arencibia, Atteneri, Desirée San Nicolás-Hernández, Carlos J. Bethencourt-Estrella, Ines Sifaoui, María Reyes-Batlle, Rubén L. Rodríguez-Expósito, Aitor Rizo-Liendo, Jacob Lorenzo-Morales, Isabel L. Bazzocchi, José E. Piñero, and et al. 2019. "Withanolides from Withania aristata as Antikinetoplastid Agents through Induction of Programmed Cell Death" Pathogens 8, no. 4: 172. https://doi.org/10.3390/pathogens8040172
APA StyleLópez-Arencibia, A., San Nicolás-Hernández, D., Bethencourt-Estrella, C. J., Sifaoui, I., Reyes-Batlle, M., Rodríguez-Expósito, R. L., Rizo-Liendo, A., Lorenzo-Morales, J., Bazzocchi, I. L., Piñero, J. E., & Jiménez, I. A. (2019). Withanolides from Withania aristata as Antikinetoplastid Agents through Induction of Programmed Cell Death. Pathogens, 8(4), 172. https://doi.org/10.3390/pathogens8040172