Molecular Identification of Zoonotic Parasites of the Genus Anisakis (Nematoda: Anisakidae) from Fish of the Southeastern Pacific Ocean (Off Peru Coast)

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Results
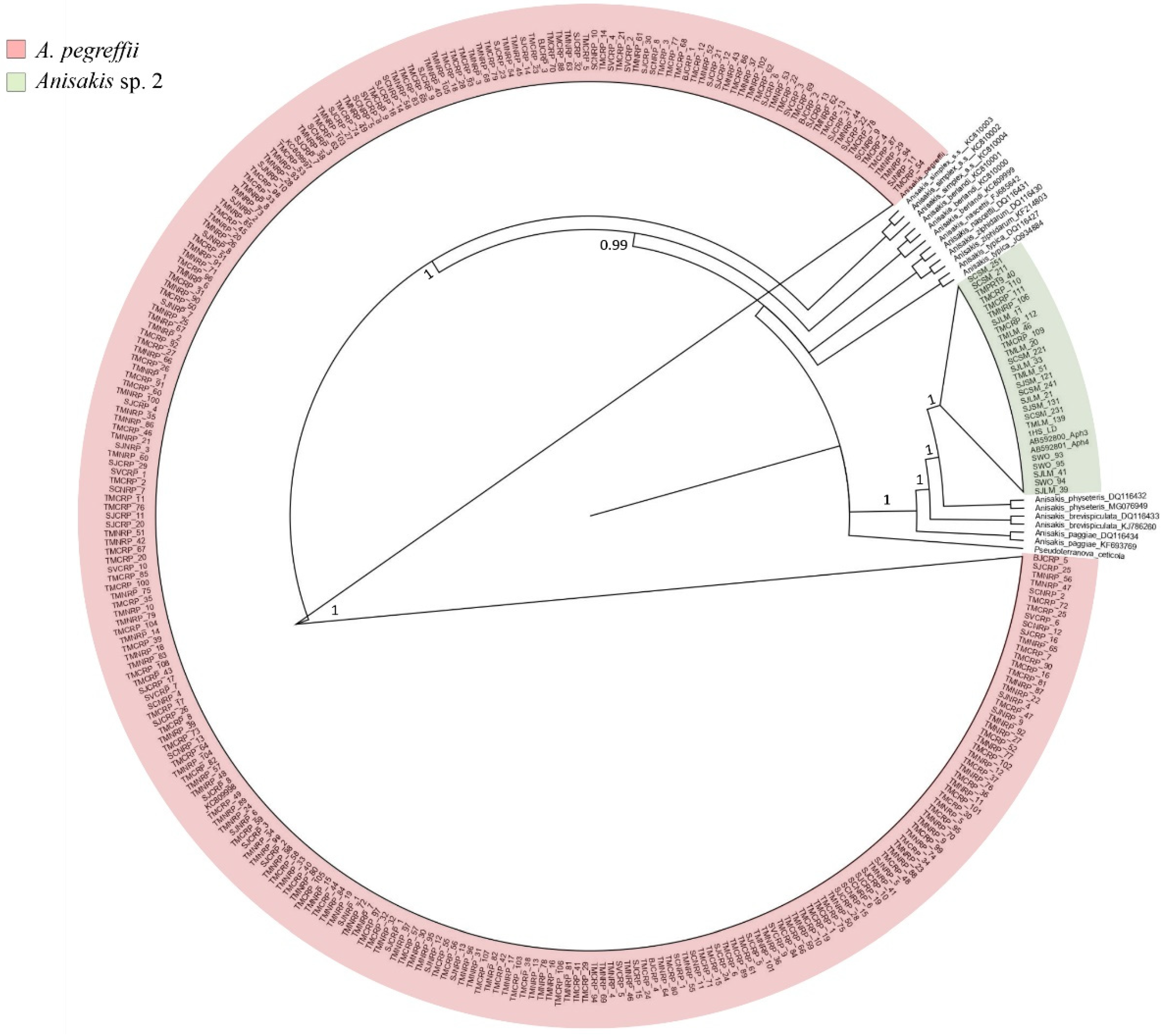
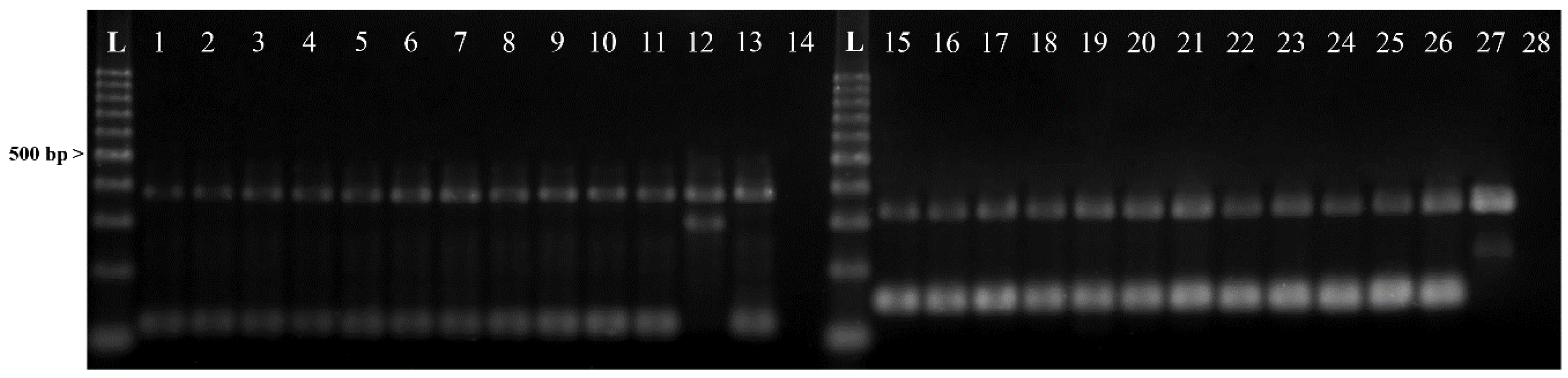
2.1. Identification of Anisakis spp.
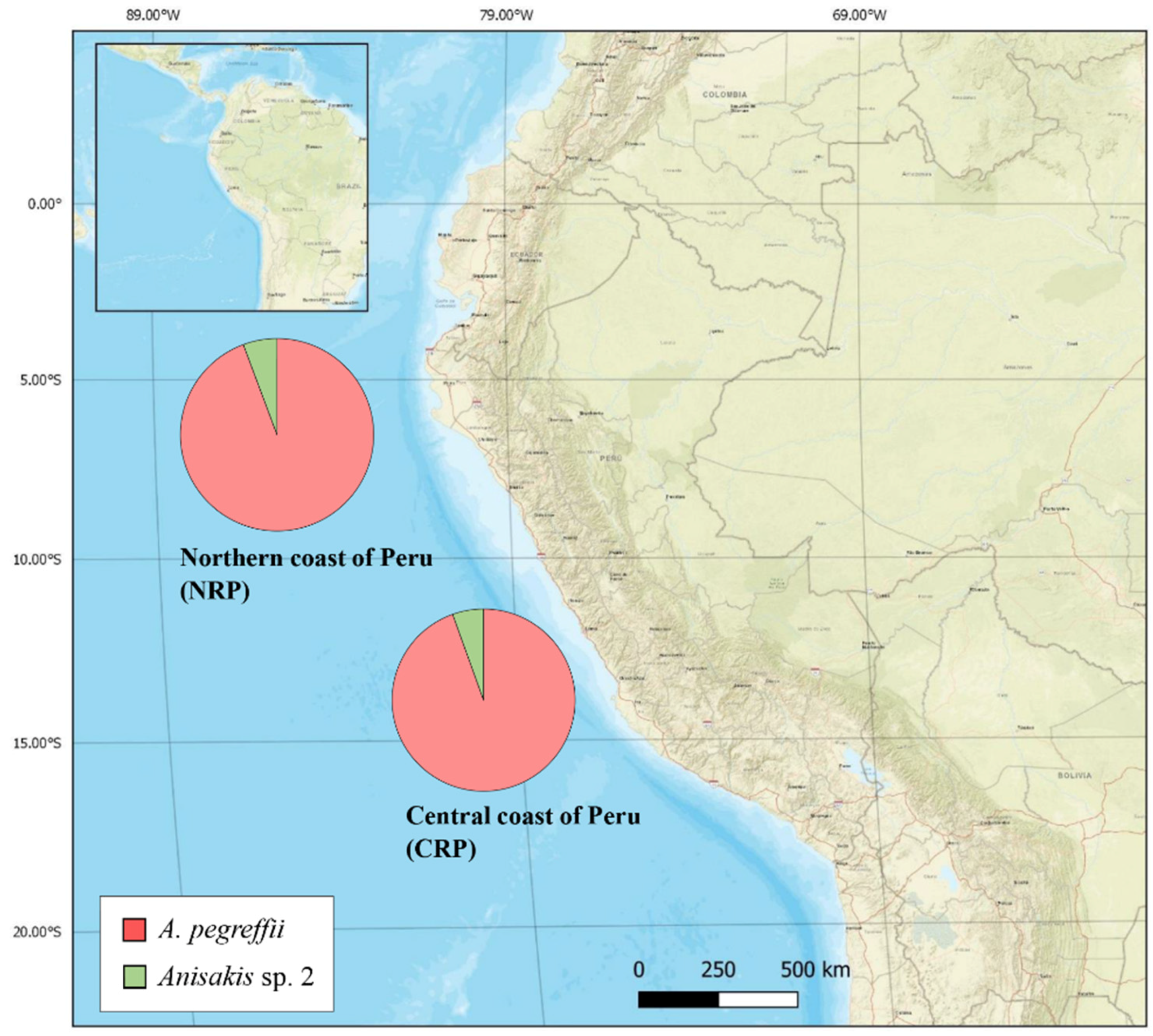
2.2. Distribution of Anisakis spp. by Fishing Ground, Fish Species, and Site of Infection
3. Discussion
4. Materials and Methods
4.1. Fish Samplings
4.2. Parasitological Analysis
4.3. Genetic Identification of Larval Nematodes
4.4. Statistical Analysis of the Parasitic Infection Parameters
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAO). The State of World Fisheries and Aquaculture 2016: Contributing to Food Security and Nutrition for All; Food & Agriculture Org: Rome, Italy, 2016; pp. 80–95. ISBN 978-925-109-185-2. [Google Scholar]
- Sanchez, D.N.; Gallo-Seminario, M. Status of and trends in the use of small pelagic fish species for reduction fisheries and for human consumption in Peru. In Fish as Feed Inputs for Aquaculture: Practices, Sustainability and Implications; Hasan, M.R., Halwart, M., Eds.; FAO: Rome, Italy, 2009; pp. 325–369. ISBN 978-92-5-106419-1. [Google Scholar]
- Fayer, R. Introduction and Public Health Importance of Foodborne Parasites. In Biology of Foodborne Parasites, 1st ed.; Xiao, L., Ryan, U., Feng, Y., Eds.; CRC Press: Boca Raton, FL, USA, 2015; Volume 1, pp. 2–20. ISBN 978-146-656-885-3. [Google Scholar]
- Mattiucci, S.; Cipriani, P.; Levsen, A.; Paoletti, M.; Nascetti, G. Molecular Epidemiology of Anisakis and Anisakiasis: An Ecological and Evolutionary Road Map. Adv. Parasitol. 2018, 99, 93–263. [Google Scholar] [CrossRef]
- Levsen, A.; Svanevik, C.S.; Cipriani, P.; Mattiucci, S.; Gay, M.; Hastie, L.C.; Bušelić, I.; Mladineo, I.; Horst, K.; Ostermeyer, U.; et al. A survey of zoonotic nematodes of commercial key fish species from major European fishing grounds—Introducing the FP7 PARASITE exposure assessment study. Fish. Res. 2018, 202, 4–21. [Google Scholar] [CrossRef]
- Mattiucci, S.; Nascetti, G. Advances and trends in the molecular systematics of anisakid nematodes, with implications for their evolutionary ecology and host-parasite co-evolutionary processes. Adv. Parasitol. 2008, 66, 47–148. [Google Scholar] [CrossRef] [PubMed]
- Chai, J.Y.; Murrell, K.D.; Lymbery, A.J. Fish-borne parasitic zoonoses: Status and issues. Int. J. Parasitol. 2005, 35, 1233–1254. [Google Scholar] [CrossRef] [PubMed]
- Audicana, M.T.; Kennedy, M.W. Anisakis simplex: From Obscure Infectious Worm to Inducer of Immune Hypersensitivity. Clin. Microbiol. Rev. 2008, 21, 360–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera, R.; Suárez-Ognio, L. Probable emergencia de anisakiosis por Anisakis physeteris durante el fenómeno El Niño 1997–1998 en la costa peruana. Parasitol. Latinoam. 2002, 57, 166–170. [Google Scholar] [CrossRef]
- Cabrera, R.; Trillo-Altamirano, M.D.P. Anisakidosis: Una zoonosis parasitaria marina desconocida o emergente en el Perú? Rev. Gastroenterol. Peru 2004, 24, 335–342. [Google Scholar]
- Eiras, J.C.; Pavanelli, G.C.; Takemoto, R.M.; Nawa, Y. Fish-borne nematodiases in South America: Neglected emerging diseases. J. Helminthol. 2018, 92, 649–654. [Google Scholar] [CrossRef]
- Barriga, J.; Salazar, F.; Barriga, E. Anisakiasis: Presentación de un caso y revisión de la literatura. Rev. Gastroenterol Peru 1999, 19, 317–323. [Google Scholar] [PubMed]
- Mercado, R.; Torres, P.; Gil, L.C.; Goldin, L. Anisakiasis en una paciente portadora de una pequeña hernia hiatal. Caso clinico. Rev. Med. Chil. 2006, 134, 1562–1564. [Google Scholar] [CrossRef] [Green Version]
- Weitzel, T.; Sugiyama, H.; Yamasaki, H.; Ramirez, C.; Rosas, R.; Mercado, R. Human infections with Pseudoterranova cattani, Chile. Emerging Infect. Dis. 2015, 21, 1874–1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menghi, C.I.; Gatta, C.L.; Arias, L.E.; Santoni, G.; Nicola, F.; Smayevsky, J.; Degese, M.F.; Krivokapich, S.J. Human infection with Pseudoterranova cattani by ingestion of “ceviche” in Buenos Aires. Argentina. Rev. Argent. Microbiol. 2020, 52, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Pérez, I.; Chávez, A.; Casas, E. Presencia de formas parasitarias en peces comerciales del mar peruano. Rev. Investig. Vet. Peru 1999, 10, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Llerena, C.; Chávez, A.; Casas, E. Frecuencia de larvas Diphyllobothriidae y larvas Anisakidae en peces marinos comerciales del terminal pesquero de ventanilla-callao. Rev. Investig. Vet. Peru 2001, 12, 58–62. [Google Scholar] [CrossRef]
- Van Waerebeek, K.; Reyes, J.C.; Alfaro, J. Helminth parasites and phoronts of dusky dolphins Lagenorhynchus obscurus (Gray, 1828) from Peru. Aquat. Mamm. 1993, 19, 159–169. [Google Scholar]
- Reyes, J.C.; Wan Waerebeek, K. Aspects of the biology of Burmeister’s porpoise from Peru. In Scientific Committee Special Meeting Report: Biology of the Phocoenids No. 16 (Report of the International Whaling Commission, Special Issue); Bjorge, A., Donovan, G.P., Eds.; International Whaling Commission: Cambridge, UK, 1995; pp. 349–364. ISBN 978-090-697-529-9. [Google Scholar]
- Oliva, M.E. Metazoan parasites of the jack mackerel Trachurus murphyi (Teleostei, Carangidae) in a latitudinal gradient from South America (Chile and Perú). Parasite 1999, 6, 223–230. [Google Scholar] [CrossRef] [Green Version]
- George-Nascimento, M. Geographical variations in the jack mackerel Trachurus symmetricus murphyi populations in the southeastern pacific ocean as evidenced from the associated parasite communities. J. Parasitol. 2000, 86, 929–932. [Google Scholar] [CrossRef]
- Muñoz, G.; Olmos, V. Revisión bibliográfica de especies endoparásitas y hospedadoras de sistemas acuáticos de Chile. Rev. Biol. Mar. Oceanogr. 2008, 43, 173–245. [Google Scholar] [CrossRef]
- Gerlotto, F.; Gutiérrez, M.; Bertrand, A. Insight on population structure of the Chilean jack mackerel (Trachurus murphyi). Aquat. Living Resour. 2012, 25, 341–355. [Google Scholar] [CrossRef] [Green Version]
- Oliva, M.E. Is Anisakis simplex s.l. a biological marker for stock identification of Strangomera bentincki from Chile? J. Fish. Biol. 2013, 83, 412–416. [Google Scholar] [CrossRef]
- George-Nascimento, M.; Oliva, M. Fish population studies using parasites from the Southeastern Pacific Ocean: Considering host population changes and species body size as sources of variability of parasite communities. Parasitology 2015, 142, 25–35. [Google Scholar] [CrossRef]
- Valentini, A.; Mattiucci, S.; Bondanelli, P.; Webb, S.C.; Mignucci-Giannone, A.A.; Colom-Llavina, M.M.; Nascetti, G. Genetic relationships among Anisakis species (Nematoda: Anisakidae) inferred from mitochondrial cox-2 sequences, and comparison with allozyme data. J. Parasitol. 2006, 92, 156–166. [Google Scholar] [CrossRef]
- Mattiucci, S.; Cipriani, P.; Webb, S.C.; Paoletti, M.; Marcer, F.; Bellisario, B.; Gibson, D.I.; Nascetti, G. Genetic and morphological approaches distinguish the three sibling species of the Anisakis simplex species complex, with a species designation as Anisakis berlandi sp. for A. simplex sp. C (Nematoda: Anisakidae). J. Parasitol. 2014, 100, 199–214. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. Bayesian Analysis of Molecular Evolution Using MrBayes. In Statistical Methods in Molecular Evolution. Statistics for Biology and Health, 1st ed.; Nielsen, R., Ed.; Springer: New York, NY, USA, 2005; pp. 183–226. ISBN 978-0-387-27733-2. [Google Scholar]
- Darriba, D.; Taboada, G.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D.; Buckley, T.R. Model Selection and Model Averaging in Phylogenetics: Advantages of Akaike Information Criterion and Bayesian Approaches over Likelihood Ratio Tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef]
- Palomba, M.; Paoletti, M.; Webb, S.; Nascetti, G.; Mattiucci, S. A novel nuclear marker and development of an ARMS-PCR assay targeting the metallopeptidase 10 (nas 10) locus to identify the species of the Anisakis simplex (s.l.) complex (Nematoda, Anisakidae). Parasite 2020, 27, 39:1–39:9. [Google Scholar] [CrossRef]
- Murata, R.; Suzuki, J.; Sadamasu, K.; Kai, A. Morphological and molecular characterization of Anisakis larvae (Nematoda: Anisakidae) in Beryx splendens from Japanese waters. Parasitol. Int. 2011, 60, 193–198. [Google Scholar] [CrossRef]
- Garcia, A.; Mattiucci, S.; Damiano, S.; Santos, M.N.; Nascetti, G. Metazoan parasites of swordfish, Xiphias gladius (Pisces: Xiphiidae) from the Atlantic Ocean: Implications for host stock identification. ICES J. Mar. Sci. 2011, 68, 175–182. [Google Scholar] [CrossRef] [Green Version]
- Mattiucci, S.; Garcia, A.; Cipriani, P.; Santos, M.N.; Nascetti, G.; Cimmaruta, R. Metazoan parasite infection in the swordfish, Xiphias gladius, from the Mediterranean Sea and comparison with Atlantic populations: Implications for its stock characterization. Parasite 2014, 21, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakun, A.; Weeks, S.J. The marine ecosystem off Peru: What are the secrets of its fishery productivity and what might its future hold? Prog. Oceanogr. 2008, 79, 290–299. [Google Scholar] [CrossRef]
- Oerder, V.; Colas, F.; Echevin, V.; Codron, F.; Tam, J.; Belmadani, A. Peru-Chile upwelling dynamics under climate change. J. Geophys. Res. Oceans 2015, 120, 1152–1172. [Google Scholar] [CrossRef] [Green Version]
- Jahncke, J.; Checkley, D.M., Jr.; Hunt, G.L., Jr. Trends in carbon flux to seabirds in the Peruvian upwelling system: Effects of wind and fisheries on population regulation. Fish. Oceanogr. 2004, 13, 208–223. [Google Scholar] [CrossRef] [Green Version]
- Passuni, G.; Barbraud, C.; Chaigneau, A.; Demarcq, H.; Ledesma, J.; Bertrand, A.; Castillo, R.; Perea, A.; Mori, J.; Viblanc, V.A.; et al. Seasonality in marine ecosystems: Peruvian seabirds, anchovy, and oceanographic conditions. Ecology 2016, 97, 182–193. [Google Scholar] [CrossRef]
- Barber, I.; Hoare, D.; Krause, J. Effects of parasites on fish behaviour: A review and evolutionary perspective. Rev. Fish. Biol. Fish. 2000, 10, 131–165. [Google Scholar] [CrossRef]
- Farjallah, S.; Slimane, B.B.; Busi, M.; Paggi, L.; Amor, N.; Blel, H.; Said, K.; D’Amelio, S. Occurrence and molecular identification of Anisakis spp. from the North African coasts of Mediterranean Sea. Parasitol. Res. 2008, 102, 371–379. [Google Scholar] [CrossRef]
- Meloni, M.; Angelucci, G.; Merella, P.; Siddi, R.; Deiana, C.; Orru, G.; Salati, F. Molecular characterization of Anisakis larvae from fish caught off Sardinia. J. Parasitol. 2011, 97, 908–914. [Google Scholar] [CrossRef]
- Mattiucci, S.; Cimmaruta, R.; Cipriani, P.; Abaunza, P.; Bellisario, B.; Nascetti, G. Integrating parasite data and host genetic structure in the frame of an holistic approach for stock identification in Mediterranean Sea fish species. Parasitology 2014, 142, 90–108. [Google Scholar] [CrossRef]
- Chou, Y.Y.; Wang, C.S.; Chen, H.G.; Chen, H.Y.; Chen, S.N.; Shih, H.H. Parasitism between Anisakis simplex (Nematoda: Anisakidae) third-stage larvae and the spotted mackerel Scomber australasicus with regard to the application of stock identification. Vet. Parasitol. 2011, 177, 324–331. [Google Scholar] [CrossRef]
- Setyobudi, E.; Jeon, C.H.; Lee, C.H.; Seong, K.B.; Kim, J.H. Occurrence and identification of Anisakis spp. (Nematoda: Anisakidae) isolated from chum salmon (Oncorhynchus keta) in Korea. Parasitol. Res. 2011, 108, 585–592. [Google Scholar] [CrossRef]
- Bak, T.J.; Jeon, C.H.; Kim, J.H. Occurrence of anisakid nematode larvae in chub mackerel (Scomber japonicus) caught off Korea. Int. J. Food Microbiol. 2014, 191, 149–156. [Google Scholar] [CrossRef]
- Chen, H.Y.; Shih, H.H. Occurrence and prevalence of fish-borne Anisakis larvae in the spotted mackerel Scomber australasicus from Taiwanese waters. Acta Trop. 2015, 145, 61–67. [Google Scholar] [CrossRef]
- Kong, Q.; Fan, L.; Zhang, J.; Akao, N.; Dong, K.; Lou, D.; Ding, J.; Tong, Q.; Zheng, B.; Chen, R.; et al. Molecular identification of Anisakis and Hysterothylacium larvae in marine fishes from the East China Sea and the Pacific coast of central Japan. Int. J. Food Microbiol. 2015, 199, 1–7. [Google Scholar] [CrossRef]
- Reyes, J.C. Ballenas, Delfines y otros Cetáceos del Perú. Una Fuente de Información, 1st ed.; Squema Ediciones: Lima, Perú, 2009; pp. 70–150. [Google Scholar]
- Tantalean, M.; Cabrera, R. Algunos helmintos de la marsopa espinosa, Phocoena spinipinnis de la Reserva Nacional de Paracas, Perú. Parasitología 1999, 23, 57–58. [Google Scholar] [CrossRef]
- Van Bressem, M.F.; Van Waerebeek, K.; Montes, D.; Kennedy, S.; Reyes, J.C.; García–Godos, I.; Ontón, K.; Alfaro, J. Diseases, lesions and malformations in the long-beaked common dolphin Delphinus capensis from the Southeast Pacific. Dis. Aquat. Org. 2006, 68, 149–165. [Google Scholar] [CrossRef] [Green Version]
- García-Godos, I.; Van Waerebeek, K.; Reyes, J.C.; Alfaro, J.; Arias-Schreiber, M. Prey occurrence in the stomach of four small cetacean species in Peru. LAJAM 2007, 6, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Mattiucci, S.; Paoletti, M.; Borrini, F.; Palumbo, M.; Palmieri, R.M.; Gomes, V.; Casati, A.; Nascetti, G. First molecular identification of the zoonotic parasite Anisakis pegreffii (Nematoda: Anisakidae) in a paraffin-embedded granuloma taken from a case of human intestinal anisakiasis in Italy. BMC Infect. Dis. 2011, 11, 82. [Google Scholar] [CrossRef]
- Santoro, M.; Di Nocera, F.; Iaccarino, D.; Cipriani, P.; Guadano Procesi, I.; Maffucci, F.; Hochscheid, S.; Blanco, C.; Cerrone, A.; Galiero, G.; et al. Helminth parasites of the dwarf sperm whale Kogia sima (Cetacea: Kogiidae) from the Mediterranean Sea, with implications on host ecology. Dis. Aquat. Org. 2018, 129, 175–182. [Google Scholar] [CrossRef]
- Céspedes, R.E.; Iannacone, J.; Salas, A. Helmintos parásitos de Dosidicus gigas “Pota” eviscerada en Arequipa, Perú. Ecol. Apl. 2011, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Mattiucci, S.; Abaunza, P.; Ramadori, L.; Nascetti, G. Genetic identification of Anisakis larvae in European hake from Atlantic and Mediterranean waters for stock recognition. J. Fish. Biol. 2004, 65, 495–510. [Google Scholar] [CrossRef]
- Cipriani, P.; Sbaraglia, G.; Palomba, L.; Giulietti, L.; Bellisario, B.; Bušelić, I.; Mladineo, I.; Cheleschi, R.; Nascetti, G.; Mattiucci, S. Anisakis pegreffii (Nematoda: Anisakidae) in European anchovy Engraulis encrasicolus from the Mediterranean Sea: Considerations in relation to fishing ground as a driver for parasite distribution. Fish. Res. 2018, 202, 59–68. [Google Scholar] [CrossRef]
- Cipriani, P.; Acerra, V.; Bellisario, B.; Sbaraglia, G.L.; Cheleschi, R.; Nascetti, G.; Mattiucci, S. Larval migration of the zoonotic parasite Anisakis pegreffii (Nematoda: Anisakidae) in European anchovy, Engraulis encrasicolus: Implications to seafood safety. Food Control. 2016, 59, 148–157. [Google Scholar] [CrossRef] [Green Version]
- Valero, A.; Martín-Sánchez, J.; Reyes-Muelas, E.; Adroher, F.J. Larval anisakids parasitizing the blue whiting, Micromesistius poutassou, from Motril Bay in the Mediterranean region of southern Spain. J. Helminthol. 2000, 74, 361–364. [Google Scholar] [CrossRef] [Green Version]
- Valero, A.; del Mar López-Cuello, M.; Benítez, R.; Adroher, F.J. Anisakis spp. in European hake, Merluccius merluccius (L.) from the Atlantic off north-west Africa and the Mediterranean off southern Spain. Acta Parasitol. 2006, 51, 209–212. [Google Scholar] [CrossRef]
- Suzuki, J.; Murata, R.; Hosaka, M.; Araki, J. Risk factors for human Anisakis infection and association between the geographic origins of Scomber japonicus and anisakid nematodes. Int. J. Food Microbiol. 2010, 137, 88–93. [Google Scholar] [CrossRef]
- Mattiucci, S.; Fazii, P.; De Rosa, A.; Paoletti, M.; Megna, A.S.; Glielmo, A.; De Angelis, M.; Costa, A.; Meucci, C.; Calvaruso, V.; et al. Anisakiasis and gastroallergic reactions associated with Anisakis pegreffii infection, Italy. Emerg. Infect. Dis. 2013, 19, 496–499. [Google Scholar] [CrossRef]
- Karl, H.; Leinemann, M. A fast and quantitative detection method for nematodes in fish fillets and fishery products. Arch. Lebensmittelhyg. 1993, 44, 105–128. [Google Scholar]
- Berland, B. Nematodes from some Norwegian marine fishes. Sarsia. 1961, 2, 1–50. [Google Scholar] [CrossRef]
- Reiczigel, J.; Rozsa, L. Quantitative Parasitology 3.0. Budapest. Distributed by the authors. Available online: http://www.zoologia.hu/qp/ (accessed on 24 April 2020).
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Rozsa, L.; Reiczigel, J.; Majoros, G. Quantifying parasites in samples of hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef]
- Reiczigel, J. Confidence intervals for the binomial parameter: Some new considerations. Stat. Med. 2003, 22, 611–621. [Google Scholar] [CrossRef]
- Reiczigel, J.; Abonyi-Tóth, Z.; Singer, J. An exact confidence set for two binomial proportions and exact unconditional confidence intervals for the difference and ratio of proportions. Comput. Stat. Data Anal. 2008, 52, 5046–5053. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | Mean total body length ± SD (Min-Max) (mm) | Mean Weight ± SD (Min-Max) (g) | Ncoll | Nid | |
|---|---|---|---|---|---|
| NRP (4°54’ S, 81°21’ W) | |||||
| Trachurus murphyi | 21 | 337.71 ± 22.85 (282–375) | 309.29 ± 30.91 (267–350) | 117 | 106 |
| Scomber japonicus | 47 | 275.27 ± 22.9 (236–345) | 253.3 ± 38.56 (192–350) | 15 | 15 |
| Sarda chiliensis | 45 | 419.33 ± 20.52 (376–473) | 1240.3 ± 226.9 (310–1700) | 20 | 20 |
| Total | 152 | 141 | |||
| CRP (12°09′ S, 77°28′ W) | |||||
| Trachurus murphyi | 100 | 257.82 ± 33.6 (243–383) | 298.17 ± 35.72 (220–460) | 212 | 112 |
| Scomber japonicus | 70 | 294.07 ± 30.88 (236–358) | 281.67 ± 52.04 (192–460) | 42 | 37 |
| Seriolella violacea | 35 | 282.67 ± 31.24 (241–379) | 275.23 ± 31.7 (189–346) | 10 | 10 |
| Brama japonica | 30 | 329.25 ± 29.34 (283–381) | 310.62 ± 21.89 (285–382) | 5 | 5 |
| Total | 269 | 164 |
| A. pegreffii | Anisakis sp. 2 | ||
|---|---|---|---|
| mtDNA cox2 | nas10 nDNA | mtDNA cox2 | |
| Trachurus murphyii | |||
| NRP | 105 | 105 | 1 |
| CRP | 108 | 108 | 4 |
| Scomber japonicus | |||
| NRP | 13 | 13 | 2 |
| CRP | 32 | 32 | 5 |
| Sarda chiliensis | |||
| NRP | 15 | 15 | 5 |
| Seriolella violacea | |||
| CRP | 10 | 10 | - |
| Brama japonica | |||
| CRP | 5 | 5 | - |
| Total | 288 | 288 | 17 |
| Fishing Area/Fish Species | Anisakis pegreffii | Anisakis sp. 2 | ||||||
|---|---|---|---|---|---|---|---|---|
| Overall | Viscera | Musculature | Overall/Viscera | |||||
| P (%) | A | P (%) | A | P (%) | A | P (%) | A | |
| NRP (4°54’ S, 81°21’ W) | ||||||||
| Trachurus murphyi | 60.0 (0.14–0.41) | 5.25 ± 5.44 | 60.0 (0.36–0.81) | 4.75 ± 4.91 | 30.0 (0.12–0.54) | 0.50 ± 0.89 | 5.0 (0.01–0.25) | 0.05 ± 0.22 |
| Scomber japonicus | 26.1 (0.36–0.81) | 0.28 ± 0.50 | 21.7 (0.11–0.36) | 0.24 ± 0.48 | 4.3 (0.01–0.15) | 0.04 ± 0.21 | 4.3 (0.01–0.15) | 0.04 ± 0.21 |
| Sarda chiliensis | 22.7 (0.12–0.38) | 0.32 ± 0.67 | 22.7 (0.12–0.38) | 0.32 ± 0.67 | - | - | 11.4 (0.04–0.25) | 0.11 ± 0.32 |
| CRP (12°09’ S, 77°28’ W) | ||||||||
| Trachurus murphyi | 39.4 (0.30–0.50) | 1.09 ± 2.39 | 38.4 (0.29–0.49) | 0.94 ± 2.13 | 14.1 (0.02–0.14) | 0.15 ± 0.39 | 4.0 (0.01–0.10) | 0.04 ± 0.20 |
| Scomber japonicus | 24.6 (0.15–0.37) | 0.46 ± 0.98 | 24.6 (0.15–0.37) | 0.39 ± 0.77 | 5.8 (0.02–0.14) | 0.07 ± 0.31 | 7.2 (0.02–0.16) | 0.07 ± 0.26 |
| Seriolella violacea | 17.6 (0.07–0.35) | 0.32 ± 0.81 | 17.6 (0.07–0.35) | 0.32 ± 0.81 | - | - | - | - |
| Brama japonica | 17.2 (0.06–0.36) | 0.17 ± 0.38 | 17.2 (0.06–0.36) | 0.17 ± 0.38 | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aco Alburqueque, R.; Palomba, M.; Santoro, M.; Mattiucci, S. Molecular Identification of Zoonotic Parasites of the Genus Anisakis (Nematoda: Anisakidae) from Fish of the Southeastern Pacific Ocean (Off Peru Coast). Pathogens 2020, 9, 910. https://doi.org/10.3390/pathogens9110910
Aco Alburqueque R, Palomba M, Santoro M, Mattiucci S. Molecular Identification of Zoonotic Parasites of the Genus Anisakis (Nematoda: Anisakidae) from Fish of the Southeastern Pacific Ocean (Off Peru Coast). Pathogens. 2020; 9(11):910. https://doi.org/10.3390/pathogens9110910
Chicago/Turabian StyleAco Alburqueque, Renato, Marialetizia Palomba, Mario Santoro, and Simonetta Mattiucci. 2020. "Molecular Identification of Zoonotic Parasites of the Genus Anisakis (Nematoda: Anisakidae) from Fish of the Southeastern Pacific Ocean (Off Peru Coast)" Pathogens 9, no. 11: 910. https://doi.org/10.3390/pathogens9110910
APA StyleAco Alburqueque, R., Palomba, M., Santoro, M., & Mattiucci, S. (2020). Molecular Identification of Zoonotic Parasites of the Genus Anisakis (Nematoda: Anisakidae) from Fish of the Southeastern Pacific Ocean (Off Peru Coast). Pathogens, 9(11), 910. https://doi.org/10.3390/pathogens9110910