Transformation of Helicobacter pylori into Coccoid Forms as a Challenge for Research Determining Activity of Antimicrobial Substances



Abstract
:
1. Introduction
2. Review Strategy and Literature Included
3. Results
3.1. Antibiotics and Proton Pump Inhibitors
3.2. Plant Extracts and Plant-derived Compounds
3.3. Synthetic Compounds
3.4. Fatty Acids and Fatty Acid Derivatives
3.5. Peptides
4. Discussion
4.1. Culture Methods in the Viability Assessing
4.2. Extending Techniques to Determine the Viability
4.2.1. Amount of ATP in the Viability Assessment
4.2.2. Morphostructure Determined by TEM in the Viability Assessment
4.2.3. Degradation of Genetic Material and eDNA Amount in the Viability Assessment
4.2.4. Fluorescence Determination in the Viability Assessment
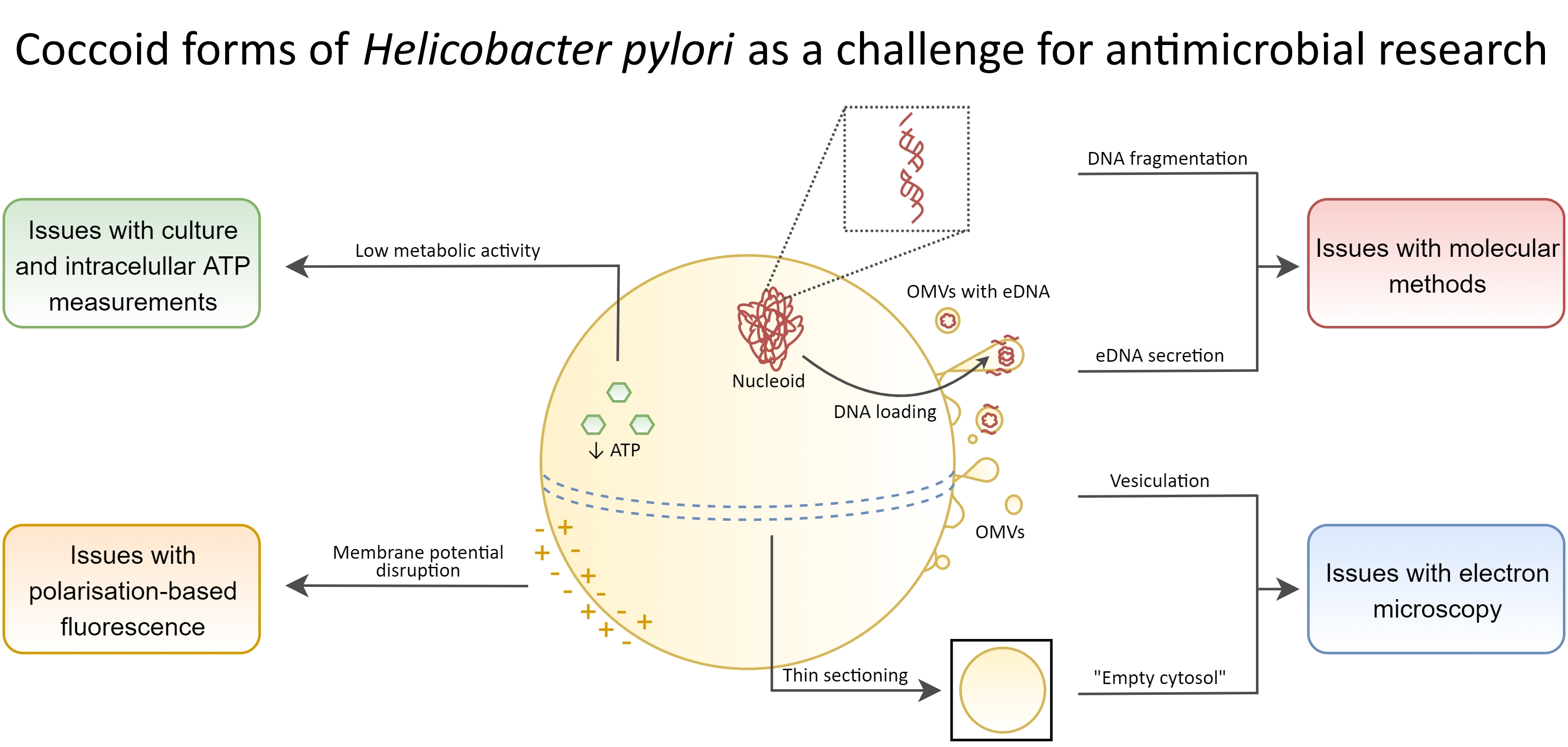
4.3. Difficulties in Determining the Viability of H. pylori in the Context of Applied Research Techniques
4.4. Future Research Determining the Viability of H. pylori
4.5. Implications for Scientists and Clinicians
5. Conclusions
- Apply several methods determining the viability of H. pylori.
- Search for new, more accurate techniques determining the viability of H. pylori.
- Test the activity of substances against preformed coccoid H. pylori forms (regardless of studies on spiral forms).
- Use coccoid H. pylori forms, not treated with antimicrobial substances, as a control of the activity of substances against these morphological forms (without reference to spiral forms characterized by different physiology).
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global Prevalence of Helicobacter pylori Infection: Systematic Review and Meta-Analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [Green Version]
- Graham, D.Y. History of Helicobacter pylori, Duodenal Ulcer, Gastric Ulcer and Gastric Cancer. World J. Gastroenterol. 2014, 20, 5191–5204. [Google Scholar] [CrossRef]
- Šterbenc, A.; Jarc, E.; Poljak, M.; Homan, M. Helicobacter pylori Virulence Genes. World J. Gastroenterol. 2019, 25, 4870–4884. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Megraud, F.; O’Morain, C.A.; Gisbert, J.P.; Kuipers, E.J.; Axon, A.T.; Bazzoli, F.; Gasbarrini, A.; Atherton, J.; Graham, D.Y.; et al. Management of Helicobacter pylori Infection—The Maastricht V/Florence Consensus Report. Gut 2017, 66, 6–30. [Google Scholar] [CrossRef] [Green Version]
- Leja, M.; Grinberga-Derica, I.; Bilgilier, C.; Steininger, C. Review: Epidemiology of Helicobacter pylori Infection. Helicobacter 2019, 24, e12635. [Google Scholar] [CrossRef] [Green Version]
- Makristathis, A.; Hirschl, A.M.; Mégraud, F.; Bessède, E. Review: Diagnosis of Helicobacter pylori Infection. Helicobacter 2019, 24, e12641. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, A.; Liou, J.; Gisbert, J.P.; O’Morain, C. Review: Treatment of Helicobacter pylori Infection 2019. Helicobacter 2019, 24, e12640. [Google Scholar] [CrossRef] [Green Version]
- Fallone, C.A.; Moss, S.F.; Malfertheiner, P. Reconciliation of Recent Helicobacter pylori Treatment Guidelines in a Time of Increasing Resistance to Antibiotics. Gastroenterology 2019, 157, 44–53. [Google Scholar] [CrossRef]
- Savoldi, A.; Carrara, E.; Graham, D.Y.; Conti, M.; Tacconelli, E. Prevalence of Antibiotic Resistance in Helicobacter pylori: A Systematic Review and Meta-analysis in World Health Organization Regions. Gastroenterology 2018, 155, 1372–1382.e17. [Google Scholar] [CrossRef] [Green Version]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, Research, and Development of New Antibiotics: The WHO Priority List of Antibiotic-Resistant Bacteria and Tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Mégraud, F. Antibiotic Resistance Is the Key Element in Treatment of Helicobacter pylori Infection. Gastroenterology 2018, 155, 1300–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellicano, R.; Ribaldone, D.G.; Fagoonee, S.; Astegiano, M.; Saracco, G.M.; Mégraud, F. A 2016 Panorama of Helicobacter pylori Infection: Key Messages for Clinicians. Panminerva Med. 2016, 58, 304–317. [Google Scholar] [PubMed]
- Krzyżek, P.; Gościniak, G. Morphology of Helicobacter pylori as a Result of Peptidoglycan and Cytoskeleton Rearrangements. Prz. Gastroenterol. 2018, 13, 182–195. [Google Scholar] [CrossRef] [Green Version]
- Catrenich, C.E.; Makin, K.M. Characterization of the Morphologic Conversion of Helicobacter pylori from Bacillary to Coccoid Forms. Scand. J. Gastroenterol. 1991, 26, 58–64. [Google Scholar] [CrossRef]
- Kusters, J.G.; Gerrits, M.M.; Van Strijp, J.A.; Vandenbroucke-Grauls, C.M. Coccoid Forms of Helicobacter pylori are the Morphologic Manifestation of Cell Death. Infect. Immun. 1997, 65, 3679. [Google Scholar] [CrossRef] [Green Version]
- Eaton, K.A.; Catrenich, C.E.; Makin, K.M.; Krakowka, S. Virulence of Coccoid and Bacillary Forms of Helicobacter pylori in Gnotobiotic Piglets. J. Infect. Dis. 1995, 171, 459–462. [Google Scholar] [CrossRef]
- Mizoguchi, H.; Fujioka, T.; Nasu, M. Evidence for Viability of Coccoid Forms of Helicobacter pylori. J. Gastroenterol. 1999, 34, 32–36. [Google Scholar]
- Zheng, P.Y.; Hua, J.; Ng, H.C.; Ho, B. Unchanged Characteristics of Helicobacter pylori during Its Morphological Conversion. Microbios 1999, 98, 51–64. [Google Scholar]
- Bode, G.; Mauch, F.; Malfertheiner, P. The Coccoid Forms of Helicobacter pylori. Criteria for Their Viability. Epidemiol. Infect. 1993, 111, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Sörberg, M.; Nilsson, M.; Hanberger, H.; Nilsson, L.E. Morphologic Conversion of Helicobacter pylori from Bacillary to Coccoid Form. Eur. J. Clin. Microbiol. Infect. Dis. 1996, 15, 216–219. [Google Scholar] [CrossRef]
- Narikawa, S.; Kawai, S.; Aoshima, H.; Kawamata, O.; Kawaguchi, R.; Hikiji, K.; Kato, M.; Iino, S.; Mizushima, Y. Comparison of the Nucleic Acids of Helical and Coccoid Forms of Helicobacter pylori. Clin. Diagn. Lab. Immunol. 1997, 4, 290. [Google Scholar] [CrossRef] [Green Version]
- Mizoguchi, H.; Fujioka, T.; Kishi, K.; Nishizono, A.; Kodama, R.; Nasu, M. Diversity in Protein Synthesis and Viability of Helicobacter pylori Coccoid Forms in Response to Various Stimuli. Infect. Immun. 1998, 66, 5555–5560. [Google Scholar] [CrossRef] [Green Version]
- Shirai, M.; Kakada, J.; Shibata, K.; Morshed, M.G.; Matsushita, T.; Nakazawa, T. Accumulation of Polyphosphate Granules in Helicobacter pylori Cells under Anaerobic Conditions. J. Med. Microbiol. 2000, 49, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Willén, R.; Carlen, B.; Wang, X.; Papadogiannakis, N.; Odselius, R.; Wadström, T. Morphologic Conversion of Helicobacter pylori from Spiral to Coccoid Form: Scanning (SEM) and Transmission Electron Microscopy (TEM) Suggest Viability. Ups. J. Med. Sci. 2000, 105, 31–40. [Google Scholar] [CrossRef]
- Saito, N.; Konishi, K.; Sato, F.; Kato, M.; Takeda, H.; Sugiyama, T.; Asaka, M. Plural Transformation-processes from Spiral to Coccoid Helicobacter pylori and Its Viability. J. Infect. 2003, 46, 49–55. [Google Scholar] [CrossRef]
- Azevedo, N.F.; Almeida, C.; Cerqueira, L.; Dias, S.; Keevil, C.W.; Vieira, M.J. Coccoid Form of Helicobacter pylori as a Morphological Manifestation of Cell Adaptation to the Environment. Appl. Environ. Microbiol. 2007, 73, 3423–3427. [Google Scholar] [CrossRef] [Green Version]
- Ng, B.L.; Quak, S.H.; Aw, M.; Goh, K.T.; Ho, B. Immune Responses to Differentiated Forms of Helicobacter pylori in Children with Epigastric Pain. Clin. Diagn. Lab. Immunol. 2003, 10, 866–869. [Google Scholar] [CrossRef] [Green Version]
- Balakrishna, J.P.; Filatov, A. Coccoid Forms of Helicobacter pylori Causing Active Gastritis. Am. J. Clin. Pathol. 2013, 140, A101. [Google Scholar] [CrossRef]
- Wang, X.; Sturegard, E.; Rupar, R.; Nilsson, H.-O.; Aleljung, P.A.; Carlen, B.; Willen, R.; Wadstrom, T. Infection of BALB/c A Mice by Spiral and Coccoid Forms of Helicobacter pylori. J. Med. Microbiol. 1997, 46, 657–663. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, N.S. Chronic Inactive Gastritis and Coccoid Helicobacter pylori in Patients Treated for Gastroesophageal Reflux Disease or with H pylori Eradication Therapy. Am. J. Clin. Pathol. 2002, 118, 719–726. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.Y.; Hui, P.K.; Leung, K.M.; Chow, J.; Kwok, F.; Ng, C.S. Coccoid Forms of Helicobacter pylori in the Human Stomach. Am. J. Clin. Pathol. 1994, 102, 503–507. [Google Scholar] [CrossRef]
- Figura, N.; Moretti, E.; Vaglio, L.; Langone, F.; Vernillo, R.; Vindigni, C.; Giordano, N. Factors Modulating the Outcome of Treatment for the Eradication of Helicobacter pylori Infection. New Microbiol. 2012, 35, 335–340. [Google Scholar]
- Chaput, C.; Ecobichon, C.; Cayet, N.; Girardin, S.E.; Werts, C.; Guadagnini, S.; Prévost, M.-C.; Mengin-Lecreulx, D.; Labigne, A.; Boneca, I.G. Role of AmiA in the Morphological Transition of Helicobacter pylori and in Immune Escape. PLoS Pathog. 2006, 2, e97. [Google Scholar] [CrossRef] [Green Version]
- Shahamat, M.; Alem, N.; Asalkhou, M.; Hamedi, N.; Alem, N.; Morshedizadeh, K.; Alem, M. IgM Antibody Response to Antigens Prepared from Vegetative and Coccoid Forms of Helicobacter pylori. Exp. Mol. Pathol. 2006, 81, 171–175. [Google Scholar] [CrossRef]
- Loke, M.F.; Ng, C.G.; Vilashni, Y.; Lim, J.; Ho, B. Understanding the Dimorphic Lifestyles of Human Gastric Pathogen Helicobacter pylori Using the SWATH-Based Proteomics Approach. Sci. Rep. 2016, 6, 26784. [Google Scholar] [CrossRef]
- Kadkhodaei, S.; Siavoshi, F.; Akbari Noghabi, K. Mucoid and Coccoid Helicobacter pylori with Fast Growth and Antibiotic Resistance. Helicobacter 2019, e12678. [Google Scholar] [CrossRef]
- Horii, T.; Mase, K.; Suzuki, Y.; Kimura, T.; Ohta, M.; Maekawa, M.; Kanno, T.; Kobayashi, M. Antibacterial Activities of β-lactamase Inhibitors Associated with Morphological Changes of Cell Wall in Helicobacter pylori. Helicobacter 2002, 7, 39–45. [Google Scholar] [CrossRef]
- Narayana, J.L.; Huang, H.N.; Wu, C.J.; Chen, J.Y. Epinecidin-1 Antimicrobial Activity: In vitro Membrane Lysis and In vivo Efficacy against Helicobacter pylori Infection in a Mouse Model. Biomaterials 2015, 61, 41–51. [Google Scholar] [CrossRef]
- Narayana, J.L.; Huang, H.N.; Wu, C.J.; Chen, J.Y. Efficacy of the Antimicrobial Peptide TP4 against Helicobacter pylori Infection: In vitro Membrane Perturbation via Micellization and In vivo Suppression of Host Immune Responses in a Mouse Model. Oncotarget 2015, 6, 12936–12954. [Google Scholar] [CrossRef] [Green Version]
- Krzyżek, P.; Franiczek, R.; Krzyżanowska, B.; Łaczmański, Ł.; Migdał, P.; Gościniak, G. In vitro Activity of 3-Bromopyruvate, an Anti-Cancer Compound, Against Antibiotic-Susceptible and Antibiotic-Resistant Helicobacter pylori Strains. Cancers 2019, 11, 229. [Google Scholar] [CrossRef] [Green Version]
- Krzyżek, P.; Franiczek, R.; Krzyżanowska, B.; Łaczmański, Ł.; Migdał, P.; Gościniak, G. In vitro Activity of Sertraline, an Antidepressant, Against Antibiotic-Susceptible and Antibiotic-Resistant Helicobacter pylori Strains. Pathogens 2019, 8, 228. [Google Scholar] [CrossRef] [Green Version]
- Faghri, J.; Poursina, F.; Moghim, S.; Zarkesh Esfahani, H.; Nasr Esfahani, B.; Fazeli, H.; Mirzaei, N.; Jamshidian, A.; Ghasemian Safaei, H. Morphological and Bactericidal Effects of Different Antibiotics on Helicobacter pylori. Jundishapur J. Microbiol. 2014, 7, e8704. [Google Scholar] [CrossRef] [Green Version]
- Obonyo, M.; Zhang, L.; Thamphiwatana, S.; Pornpattananangkul, D.; Fu, V.; Zhang, L. Antibacterial Activities of Liposomal Linolenic Acids against Antibiotic-Resistant Helicobacter pylori. Mol. Pharm. 2012, 9, 2677–2685. [Google Scholar] [CrossRef] [Green Version]
- Saniee, P.; Shahreza, S.; Siavoshi, F. Negative Effect of Proton-Pump Inhibitors (PPIs) on Helicobacter pylori Growth, Morphology, and Urease Test and Recovery after PPI Removal—An In vitro Study. Helicobacter 2016, 21, 143–152. [Google Scholar] [CrossRef]
- Matsushima, M.; Suzuki, T.; Masui, A.; Kasai, K.; Kouchi, T.; Takagi, A.; Shirai, T.; Mine, T. Growth Inhibitory Action of Cranberry on Helicobacter pylori. J. Gastroenterol. Hepatol. 2008, 23, S175–S180. [Google Scholar] [CrossRef]
- Ali, S.M.; Khan, A.A.; Ahmed, I.; Musaddiq, M.; Ahmed, K.S.; Polasa, H.; Rao, L.V.; Habibullah, C.M.; Sechi, L.A.; Ahmed, N. Antimicrobial Activities of Eugenol and Cinnamaldehyde against the Human Gastric Pathogen Helicobacter pylori. Ann. Clin. Microbiol. Antimicrob. 2005, 4, 20. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, C.R.; Michel, V.; Genovese, S.; Prévost, M.-C.; Epifano, F.; Touati, E. Anti-Helicobacter pylori Activities of Natural Isopentenyloxycinnamyl Derivatives from Boronia pinnata. Nat. Prod. Commun. 2012, 7, 1350. [Google Scholar] [CrossRef]
- Moon, S.H.; Lee, J.H.; Kim, K.T.; Park, Y.S.; Nah, S.Y.; Ahn, D.U.; Paik, H.D. Antimicrobial Effect of 7-O-butylnaringenin, a Novel Flavonoid, and Various Natural Flavonoids against Helicobacter pylori Strains. Int. J. Environ. Res. Public Health 2013, 10, 5459–5469. [Google Scholar] [CrossRef]
- Goswami, S.; Bhakuni, R.S.; Chinniah, A.; Pal, A.; Kar, S.K.; Das, P.K. Anti-Helicobacter pylori Potential of Artemisinin and Its Derivatives. Antimicrob. Agents Chemother. 2012, 56, 4594–4607. [Google Scholar] [CrossRef] [Green Version]
- Ngan, L.T.M.; Moon, J.-K.; Shibamoto, T.; Ahn, Y.-J. Growth-Inhibiting, Bactericidal, and Urease Inhibitory Effects of Paeonia lactiflora Root Constituents and Related Compounds on Antibiotic-Susceptible and -Resistant Strains of Helicobacter pylori. J. Agric. Food Chem. 2012, 60, 9062–9073. [Google Scholar] [CrossRef]
- Lee, H.-K.; Song, H.E.; Lee, H.-B.; Kim, C.-S.; Koketsu, M.; Thi My Ngan, L.; Ahn, Y.-J. Growth Inhibitory, Bactericidal, and Morphostructural Effects of Dehydrocostus Lactone from Magnolia sieboldii Leaves on Antibiotic-Susceptible and -Resistant Strains of Helicobacter pylori. PLoS ONE 2014, 9, e95530. [Google Scholar] [CrossRef]
- Damasceno, J.P.L.; Rodrigues, R.P.; Gonçalvesa, R.D.C.R.; Kitagawa, R.R. Anti-Helicobacter pylori Activity of Isocoumarin Paepalantine: Morphological and Molecular Docking Analysis. Molecules 2017, 22, 786. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.F.; Lian, D.W.; Chen, Y.Q.; Cai, Y.F.; Zheng, Y.F.; Fan, P.L.; Ren, W.K.; Fu, L.J.; Li, Y.C.; Xie, J.H.; et al. In vitro and In vivo Antibacterial Activities of Patchouli Alcohol, a Naturally Occurring Tricyclic Sesquiterpene, against Helicobacter pylori Infection. Antimicrob. Agents Chemother. 2017, 61, e00122-17. [Google Scholar] [CrossRef] [Green Version]
- Romero, M.; Freire, J.; Pastene, E.; García, A.; Aranda, M.; González, C. Propolis Polyphenolic Compounds Affect the Viability and Structure of Helicobacter pylori In vitro. Brazilian J. Pharmacogn. 2019, 29, 325–332. [Google Scholar] [CrossRef]
- Wang, G.; Pang, J.; Hu, X.; Nie, T.; Lu, X.; Li, X.; Wang, X.; Lu, Y.; Yang, X.; Jiang, J.; et al. Daphnetin: A Novel Anti-Helicobacter pylori Agent. Int. J. Mol. Sci. 2019, 20, 850. [Google Scholar] [CrossRef] [Green Version]
- Barry, D.P.; Asim, M.; Leiman, D.A.; de Sablet, T.; Singh, K.; Casero, R.A.; Chaturvedi, R.; Wilson, K.T. Difluoromethylornithine is a Novel Inhibitor of Helicobacter pylori Growth, cagA Translocation, and Interleukin-8 Induction. PLoS ONE 2011, 6, e17510. [Google Scholar] [CrossRef] [Green Version]
- Tharmalingam, N.; Port, J.; Castillo, D.; Mylonakis, E. Repurposing the Anthelmintic Drug Niclosamide to Combat Helicobacter pylori. Sci. Rep. 2018, 8, 3701. [Google Scholar] [CrossRef] [Green Version]
- Chakraborti, S.; Bhattacharya, S.; Chowdhury, R.; Chakrabarti, P. The Molecular Basis of Inactivation of Metronidazole-Resistant Helicobacter pylori Using Polyethyleneimine Functionalized Zinc Oxide Nanoparticles. PLoS ONE 2013, 8, e70776. [Google Scholar] [CrossRef] [Green Version]
- Inoue, M.; Segawa, K.; Matsunaga, S.; Matsumoto, N.; Oda, M.; Yamase, T. Antibacterial Activity of Highly Negative Charged Polyoxotungstates, K 27[KAs4W40O140] and K 18[KSb9W21O86], and Keggin-structural Polyoxotungstates against Helicobacter pylori. J. Inorg. Biochem. 2005, 99, 1023–1031. [Google Scholar] [CrossRef]
- Dai, G.; Cheng, N.; Dong, L.; Muramatsu, M.; Xiao, S.; Wang, M.-W.; Zhu, D.-X. Bactericidal and Morphological Effects of NE-2001, a Novel Synthetic Agent Directed against Helicobacter pylori. Antimicrob. Agents Chemother. 2005, 49, 3468–3473. [Google Scholar] [CrossRef] [Green Version]
- Kamoda, O.; Anzai, K.; Mizoguchi, J.I.; Shiojiri, M.; Yanagi, T.; Nishino, T.; Kamiya, S. In vitro Activity of a Novel Antimicrobial Agent, TG44, for Treatment of Helicobacter pylori Infection. Antimicrob. Agents Chemother. 2006, 50, 3062–3069. [Google Scholar] [CrossRef] [Green Version]
- Makobongo, M.O.; Einck, L.; Peek, R.M.; Merrell, D.S. In vitro Characterization of the Anti-Bacterial Activity of SQ109 against Helicobacter pylori. PLoS ONE 2013, 8, e68917. [Google Scholar] [CrossRef]
- Correia, M.; Michel, V.; Matos, A.A.; Carvalho, P.; Oliveira, M.J.; Ferreira, R.M.; Dillies, M.-A.; Huerre, M.; Seruca, R.; Figueiredo, C.; et al. Docosahexaenoic Acid Inhibits Helicobacter pylori Growth In vitro and Mice Gastric Mucosa Colonization. PLoS ONE 2012, 7, e35072. [Google Scholar] [CrossRef]
- Yonezawa, H.; Osaki, T.; Hanawa, T.; Kurata, S.; Zaman, C.; Woo, T.D.H.; Takahashi, M.; Matsubara, S.; Kawakami, H.; Ochiai, K.; et al. Destructive Effects of Butyrate on the Cell Envelope of Helicobacter pylori. J. Med. Microbiol. 2012, 61, 582–589. [Google Scholar] [CrossRef]
- Huang, Y.; Hang, X.; Jiang, X.; Zeng, L.; Jia, J.; Xie, Y.; Li, F.; Bi, H. In vitro and In vivo Activities of Zinc Linolenate, a Selective Antibacterial Agent against Helicobacter pylori. Antimicrob. Agents Chemother. 2019, 63, e00004-19. [Google Scholar] [CrossRef] [Green Version]
- Makobongo, M.O.; Gancz, H.; Carpenter, B.M.; McDaniel, D.P.; Merrell, D.S. The Oligo-acyl Lysyl Antimicrobial Peptide C12K- 2β12 Exhibits a Dual Mechanism of Action and Demonstrates Strong In Vivo Efficacy against Helicobacter pylori. Antimicrob. Agents Chemother. 2012, 56, 378–390. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Ma, L.; Huang, Y.; Wu, H.; Dou, J.; Zhou, C. Antimicrobial Activities of Peptide Cbf-K16 against Drug-resistant Helicobacter pylori Infection In vitro and In vivo. Microb. Pathog. 2019, 138, 103847. [Google Scholar] [CrossRef]
- Xiong, M.; Bao, Y.; Xu, X.; Wang, H.; Han, Z.; Wang, Z.; Liu, Y.; Huang, S.; Song, Z.; Chen, J.; et al. Selective Killing of Helicobacter pylori with pH-responsive Helix–coil Conformation Transitionable Antimicrobial Polypeptides. Proc. Natl. Acad. Sci. USA 2017, 114, 12675–12680. [Google Scholar] [CrossRef] [Green Version]
- Mazaheri Assadi, M.; Chamanrokh, P.; Whitehouse, C.A.; Huq, A. Methods for Detecting the Environmental Coccoid Form of Helicobacter pylori. Front. Public Heal. 2015, 3, 147. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.S.; Ghosh, A.R. Assessment of Bacterial Viability: A Comprehensive Review on Recent Advances and Challenges. Microbiology (U.K.) 2019, 165, 593–610. [Google Scholar] [CrossRef]
- Nilsson, H.-O.; Blom, J.; Abu-Al-Soud, W.; Ljungh, A.A.; Andersen, L.P.; Wadström, T. Effect of Cold Starvation, Acid Stress, and Nutrients on Metabolic Activity of Helicobacter pylori. Appl. Environ. Microbiol. 2002, 68, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enroth, H.; Wreiber, K.; Rigo, R.; Risberg, D.; Uribe, A.; Engstrand, L. In vitro Aging of Helicobocter pylori: Changes in Morphology, Intracellular Composition and Surface Properties. Helicobacter 1999, 4, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Achbergerová, L.; Nahálka, J. Polyphosphate—An Ancient Energy Source and Active Metabolic Regulator. Microb. Cell Fact. 2011, 10, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, N.; Kwan, B.W.; Wood, T.K. Persistence Increases in the Absence of the Alarmone Guanosine Tetraphosphate by Reducing Cell Growth. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Bode, G.; Mauch, F.; Ditschuneit, H.; Malfertheiner, P. Identification of Structures Containing Polyphosphate in Helicobacter pylori. J. Gen. Microbiol. 1993, 139, 3029–3033. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.X.; Zhou, Y.N.; Yang, Y.; Jin, D.J. Polyphosphate Binds to the Principal Sigma Factor of RNA Polymerase during Starvation Response in Helicobacter pylori. Mol. Microbiol. 2010, 77, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Bruhn-Olszewska, B.; Szczepaniak, P.; Matuszewska, E.; Kuczyńska-Wiśnik, D.; Stojowska-Swędrzyńska, K.; Moruno Algara, M.; Laskowska, E. Physiologically Distinct Subpopulations Formed in Escherichia coli Cultures in Response to Heat Shock. Microbiol. Res. 2018, 209, 33–42. [Google Scholar] [CrossRef]
- Kim, J.S.; Chowdhury, N.; Yamasaki, R.; Wood, T.K. Viable But Non-culturable and Persistence Describe the Same Bacterial Stress State. Environ. Microbiol. 2018, 20, 2038–2048. [Google Scholar] [CrossRef]
- Zhao, F.; Bi, X.; Hao, Y.; Liao, X. Induction of Viable but Nonculturable Escherichia coli O157:H7 by High Pressure CO2 and Its Characteristics. PLoS ONE 2013, 8, e62388. [Google Scholar] [CrossRef] [Green Version]
- Ayrapetyan, M.; Williams, T.; Oliver, J.D. Relationship between the Viable but Nonculturable State and Antibiotic Persister Cells. J. Bacteriol. 2018, 200, e00249-18. [Google Scholar] [CrossRef] [Green Version]
- Pu, Y.; Li, Y.; Jin, X.; Tian, T.; Ma, Q.; Zhao, Z.; Lin, S.Y.; Chen, Z.; Li, B.; Yao, G.; et al. ATP-Dependent Dynamic Protein Aggregation Regulates Bacterial Dormancy Depth Critical for Antibiotic Tolerance. Mol. Cell 2019, 73, 143–156.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, A.; Park, A.; Nagata, K.; Sato, E.F.; Kashiba, M.; Tamura, T.; Inoue, M. Oxidative Cellular Damage Associated with Transformation of Helicobacter pylori from a Bacillary to a Coccoid Form. Free Radic. Biol. Med. 2000, 28, 1611–1618. [Google Scholar] [CrossRef]
- Park, A.M.; Li, Q.; Nagata, K.; Tamura, T.; Shimono, K.; Sato, E.F.; Inoue, M. Oxygen Tension Regulates Reactive Oxygen Generation and Mutation of Helicobacter pylori. Free Radic. Biol. Med. 2004, 36, 1126–1133. [Google Scholar] [CrossRef] [PubMed]
- Monstein, H.J.; De La Cour, C.D.; Jonasson, J. Probing 23S Ribosomal RNA Cleavage Sites in Coccoid Helicobacter pylori. Helicobacter 2001, 6, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Monstein, H.J.; Tiveljung, A.; Jonasson, J. Non-random Fragmentation of Ribosomal RNA in Helicobacter pylori during Conversion to the Coccoid Form. FEMS Immunol. Med. Microbiol. 1998, 22, 217–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grande, R.; Di Marcantonio, M.C.; Robuffo, I.; Pompilio, A.; Celia, C.; Di Marzio, L.; Paolino, D.; Codagnone, M.; Muraro, R.; Stoodley, P.; et al. Helicobacter pylori ATCC 43629/NCTC 11639 Outer Membrane Vesicles (OMVs) from Biofilm and Planktonic Phase Associated with Extracellular DNA (eDNA). Front. Microbiol. 2015, 6, 1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puca, V.; Ercolino, E.; Celia, C.; Bologna, G.; Di Marzio, L.; Mincione, G.; Marchisio, M.; Miscia, S.; Muraro, R.; Lanuti, P.; et al. Detection and Quantification of eDNA-Associated Bacterial Membrane Vesicles by Flow Cytometry. Int. J. Mol. Sci. 2019, 20, 5307. [Google Scholar] [CrossRef] [Green Version]
- Grande, R.; Di Giulio, M.; Bessa, L.J.; Di Campli, E.; Baffoni, M.; Guarnieri, S.; Cellini, L. Extracellular DNA in Helicobacter pylori Biofilm: A Backstairs Rumour. J. Appl. Microbiol. 2011, 110, 490–498. [Google Scholar] [CrossRef]
- Grande, R.; Di Campli, E.; Di Bartolomeo, S.; Verginelli, F.; Di Giulio, M.; Baffoni, M.; Bessa, L.J.; Cellini, L. Helicobacter pylori Biofilm: A Protective Environment for Bacterial Recombination. J. Appl. Microbiol. 2012, 113, 669–676. [Google Scholar] [CrossRef]
- Yoon, J.-H.; Lee, S.-Y. Characteristics of Viable-but-nonculturable Vibrio parahaemolyticus Induced by Nutrient-deficiency at Cold Temperature. Crit. Rev. Food Sci. Nutr. 2019, 1–19. [Google Scholar] [CrossRef]
- Lee, S.; Bae, S. Molecular Viability Testing of Viable But Non-culturable Bacteria Induced by Antibiotic Exposure. Microb. Biotechnol. 2018, 11, 1008–1016. [Google Scholar] [CrossRef] [Green Version]
- Lazou, T.P.; Iossifidou, E.G.; Gelasakis, A.I.; Chaintoutis, S.C.; Dovas, C.I. Viability Quantitative PCR Utilizing Propidium Monoazide, Spheroplast Formation, and Campylobacter coli as a Bacterial Model. Appl. Environ. Microbiol. 2019, 85, e01499-19. [Google Scholar] [CrossRef] [PubMed]
- Robben, C.; Witte, A.K.; Schoder, D.; Stessl, B.; Rossmanith, P.; Mester, P. A Fast and Easy ATP-based Approach Enables MIC Testing for Non-resuscitating VBNC Pathogens. Front. Microbiol. 2019, 10, 1365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Zimaity, H.; Serra, S.; Szentgyorgyi, E.; Vajpeyi, R.; Samani, A. Gastric Biopsies: The Gap between Evidence-based Medicine and Daily Practice in the Management of Gastric Helicobacter pylori Infection. Can. J. Gastroenterol. 2013, 27, e25–e30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalali, B.; Formichella, L.; Gerhard, M. Diagnosis of Helicobacter pylori: Changes towards the Future. Diseases 2015, 3, 122–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.K.; Pratap, C.B.; Jain, A.K.; Gulati, A.K.; Nath, G. Diagnosis of Helicobacter pylori: What Should be the Gold Standard? World J. Gastroenterol. 2014, 20, 12847–12859. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | H. pylori Strain(s) | Technic Determining Morphology | Technic Determining Viability | Main Results Covering the Morphological Transformation of H. pylori | Authors Interpretation | Bibliography |
|---|---|---|---|---|---|---|
| SUL TAZ CLV AMX AMX + CLV | ATCC 43504 | Light microscopy, TEM | Culture | - Viability ↓ <1 log, 6 h, 10× MIC (CLV, SUL, and TAZ) - Coccoid forms with CLV, SUL, and TAZ (6 h, 10× MIC) - Viability ↓ 1 log, 3 h, MIC (AMX, AMX+CLV) - Coccoid forms with AMX and AMX+CLV (3 h, MIC) - Separation of membranes for all antimicrobials (3–6 h, MIC) | Bactericidal activity | [37] |
| AMX | ATCC 43504 | TEM | Culture, Fluorescence (membrane integrity) | - Viability ↓ (3 log, 24 h, MIC) - Coccoid forms (2 h, MIC) - Membrane permeabilization (2 h, MIC) | Bactericidal activity | [38,39] * |
| CLR TET MTZ AMX | Tx30a | Light microscopy | Culture | - Total viability ↓ for all antibiotics (72 h, MIC) - ≥85% of coccoid forms for all antibiotics (72 h, MIC) - Approx. 30% (¼× MIC) and 70% (½× MIC) of coccoid forms with AMX (72 h) - <15% (¼× MIC) and 25–30% (½× MIC) of coccoid forms with CLR, TET, or MTZ (72 h) | Bactericidal activity, AMX is the strongest inducer of a spiral-to-coccoid form transformation | [40,41] * |
| AMX MTZ CLR | ATCC 700392 | Light microscopy | Flow cytometry (membrane integrity) | - Approx. 30%, 40%, and 95% of coccoid forms with CLR, MTZ, and AMX, respectively (72 h, ½× MIC) - Viability ↓ of coccoid forms by 99.8%, 62.6%, and 0.03% with CLR, MTZ, and AMX, respectively (24 h, 2× MIC) | Bactericidal activity, lower susceptibility of coccoid forms to antibiotics (especially to AMX) | [42] |
| AMX | SS1 | SEM | Culture, Fluorescence (membrane integrity) | - Total viability ↓ of spiral forms (24 h, MIC) - Viability ↓ of coccoid forms only by 20% (24 h, 20× MIC) | Bactericidal activity, coccoid forms are more tolerant to AMX than spiral forms | [43] * |
| Omeprazole | Clinical isolates | Light microscopy | Culture | - Viability ↓ was directly associated with an amount of coccoid forms - 10%, 40%, and 90% of coccoid forms after 3, 6, and 9 days, respectively (½× MIC) - Viability and spiral shape were recovered after 9–12 days of the omeprazole removal | Inhibitory activity with the reversibility of morphology | [44] |
| Compound | H. pylori Strain(s) | Technic Determining Morphology | Technic Determining Viability | Main Results Covering the Morphological Transformation of H. pylori | Authors Interpretation | Bibliography |
|---|---|---|---|---|---|---|
| Cranberry extract | NCTC 11637 | SEM | Culture | - Viability ↓ (2 log, 48 h, MIC) - 33% of coccoid forms (48 h, MIC) | Inhibitory activity | [45] |
| Eugenol Cinnamaldehyde | ATCC 700392 | Light microscopy | Culture | - Total viability ↓ (9 h, MBC or 6 h, 2× MBC) - Viability ↓ (>3 log, 4 h, 2× MBC) - 70% of coccoid forms (4 h, 2× MBC) | Inhibitory activity | [46] |
| Boropinol A | B128 | SEM | Culture | - Viability ↓ (several times lower OD, 48 h, MIC) - Spiral forms with shorter length and blebbing (24 h, MIC) | Inhibitory activity | [47] |
| 7-O-Butylnaringenin | ATCC 700392 | SEM | Culture | - Viability ↓ (<3 log, 24 h, MIC) - Coccoid-like cells with irregular shapes (24 h, MIC) | Inhibitory activity | [48] |
| β-ACPM | ATCC 43504 | TEM | Culture | - Total viability ↓ (18 h, MIC) - Coccoid forms with blebbing, detachment of the outer cell membrane, and membrane invaginations (24 h, MIC) | Bactericidal activity | [49] |
| Methyl gallate Benzoic acid PGG Paeonol | ATCC 43504 | Light microscopy | Culture | - Viability ↓ (1–2 log, 48 h, MIC) - Viability ↓ (≥5 log, 48 h, 2× MIC) - 43–52% of coccoid forms (48 h, MIC) - 74–91% of coccoid forms (48 h, 2× MIC) | Bactericidal activity | [50] |
| DCL from Magnolia sieboldii leaves | ATCC 43504 | SEM | Culture | - Total viability ↓ (36 h, 2× MIC) - Viability ↓ (2 log, 48 h, MIC) - 49% and 94% of coccoid forms (48 h, MIC and 2× MIC, respectively) | Bactericidal activity | [51] |
| Isocoumarin paepalantine | ATCC 43504 | SEM | Culture | - Total viability ↓ (72 h, MIC) - Spiral forms (72 h, ½× MIC) - Coccoid forms (72 h, MIC) | Bactericidal activity | [52] |
| Patchouli alcohol | NCTC 11637 | SEM, TEM | Culture | - Viability ↓ (1 log, 6 h, 2× MIC) - Mix of both spiral and coccoid forms (72 h, sub-MICs) - Coccoid forms with blebbing, cell wall damage, and lysis of the cytoplasmic membrane (1 h, MIC or 2 h, 2× MIC) | Bactericidal activity | [53] |
| Chrysin (CH) Pinocembrin (P) Galangin (G) Caffeic acid (CA) | ATCC 43504 | TEM | Culture | - Viability ↓ (4 log, 12 h, FIC of CH+P) - Viability ↓ (3 log, 12 h, FIC of G+CA) - Ghost (degenerative) cells with coccoid-like shape, membrane damage, and membrane vesicles formation (12 h, FIC of CH+P or G+CA) | Bactericidal activity | [54] |
| Daphnetin | ATCC 43504 | SEM, TEM | Culture, Fluorescence + CLSM + Flow cytometry (membrane integrity) | - Spiral forms (72 h, ¼× MIC) - Single coccoid forms (72 h, ½× MIC) - Budding, rough outer membrane, peculiar detachments between membrane and cytoplasm, and numerous membrane vesicles (72 h, ½× MIC) - No membrane permeability and depolarization (24 h, ½× MIC) - DNA damage (24 h, ½× MIC) | Daphnetin has a potential to be an effective anti-H. pylori agent | [55] |
| Compound | H. pylori Strain(s) | Technic Determining Morphology | Technic Determining Viability | Main Results Covering the Morphological Transformation of H. pylori | Authors Interpretation | Bibliography |
|---|---|---|---|---|---|---|
| DFMO | SS1 | Light microscopy, TEM | Culture, Luminescence (ATP level) | - Viability ↓ (<1 log, 24 h, MIC) - Coccoid forms (6 h, MIC) - Reversion from coccoid to spiral forms (6 h after the DFMO removal) - ↑ iATP level (0–12 h, MIC), no difference later (24 h, MIC) | Inhibitory activity with the reversibility of morphology | [56] |
| Niclosamide | ATCC 49503 | SEM | Culture, Fluorescence (membrane integrity) | - Viability ↓ (approx. 1.5 log, 24 h, 4× MIC) - Coccoid forms produced in a dose-dependent manner (3 h, MIC to 8× MIC) - No membrane permeabilization (1 h, 64× MIC) | Inhibitory activity | [57] |
| Polyethyleneimine functionalized zinc oxide | ATCC 700392 | SEM, TEM | Culture, Luminescence (ATP level), qRT-PCR (RNA degradation), Fluorescence (membrane integrity) | - Viability ↓ (1 log, 24 h, MIC) - 10% (0.5 h, MIC) and 33% (3 h, MIC) of coccoid forms - Membrane damage (3 h, MIC) - ↓ iATP level (3 h, MIC) - Degradation of rRNA (time-dependent, MIC) | Inhibitory activity | [58] |
| Polyoxometalates: As4W40 Sb9W21 SiVW11 | ATCC 43504 | SEM | Culture | - Total viability ↓ for As4W40 and Sb9W21 (10× MIC, 24 h and 12 h, respectively) - Stable viability for SiVW11 (10× MIC, 24 h) - Intensive formation of coccoids for As4W40 and Sb9W21 (36 h, > MICs) - Weak formation of coccoids for SiVW11 (36 h, > MICs) | Bactericidal (As4W40 and SB9W21) or bacteriostatic (SiVW11) activity | [59] |
| NE-2001 | ATCC 43504 | TEM | Culture | - Viability ↓ (1 log, 24 h, MIC) - Coccoid forms with an intact membrane (6 h, ½× MIC) - Ghost (degenerative) cells with coccoid-like shape with a loosening of the outer membrane (6 h, MIC) | Bactericidal activity | [60] |
| TG44 | ATCC 43504 | SEM, TEM | Culture | - Total viability ↓ (24 h, 8× MIC) - Viability ↓ (<1 log, 24 h, MIC to 4× MIC) - Spiral forms with membrane vesicles around (3 h, ½× MIC to 4× MIC) - Coccoid forms with blebbing and outer membrane detachment (3 h, 64× MIC) | Bactericidal activity | [61] |
| SQ109 | G27 | TEM | Culture, Fluorescence (membrane integrity) | - Total viability ↓ (8 h, 2× MBC) - Coccoid forms (98–99%) with disintegration of the inner membrane (8 h, 2× MBC) - Ghost (degenerative) cells (1–2%) (8 h, 2× MBC) - No membrane disruption using fluorescence analysis (10 h, 2× MBC) | Bactericidal activity | [62] |
| 3-Bromopyruvate | Tx30a J99 | Light microscopy, SEM | Culture, Fluorescence (membrane integrity) | - Total viability ↓ (6 h, 4 h, and 2 h for MIC, 2× MIC, and 4× MIC, respectively) - 33.5% (Tx30a) and 57.5% (J99) of coccoid forms (2 h, 4× MIC) - 97–98% of coccoid forms for both (24 h, 4× MIC) - ↓ green/red fluorescence (time-dependent, MIC to 4× MIC) | Bactericidal activity | [40] |
| Sertraline | Tx30a J99 | Light microscopy, SEM | Culture, Fluorescence (membrane integrity) | - Total viability ↓ (8 h, 4× MIC for J99 or 24 h, 4× MIC for Tx30a) - Viability ↓ (2–4 log, 24 h, MIC) - > 85% of coccoid forms (8 h, 4× MIC for J99 or 24 h, 4× MIC for Tx30a) - ↓ green/red fluorescence (time-dependent, MIC to 4× MIC) | Bactericidal activity | [41] |
| Compound | H. pylori Strain(s) | Technic Determining Morphology | Technic Determining Viability | Main Results Covering the Morphological Transformation of H. pylori | Authors Interpretation | Bibliography |
|---|---|---|---|---|---|---|
| Docosahexaenoic acid | ATCC 700392, SS1 | SEM | Culture | - Viability ↓ (<2 log, 48 h, MIC) - Mixed population of spiral and coccoid forms (48 h, MIC) | Inhibitory activity | [63] |
| Sodium butyrate | TK1402 | SEM | Culture, PCR (eDNA detection) | - Stable viability (24 h, MBC) - Viability ↓ (4 log, 48 h, MBC) - Coccoid forms with blebs and cell envelope alternations (24 h, MBC) - ↑ eDNA amount (24 h, MBC) | Bactericidal activity | [64] |
| Zinc linolenate | G27 | TEM | Culture, Luminescence (ATP level), Fluorescence (membrane integrity) | - Total viability ↓ (70 min, MIC) - Coccoid forms with membrane detachments, cytoplasmic content leakage, and vesiculation (2 h, MIC) - Membrane permeabilization (2 h, MIC) - ↓ iATP (2 h, MIC) and ↑ eATP (2 h, MIC) | Bactericidal activity | [65] |
| Liposomal linolenic acid | SS1 | SEM | Culture, Fluorescence (membrane integrity) | - Total viability ↓ influenced by the morphology - MBC was higher for coccoid forms than spiral forms and killed cells more slowly (24 h, 400 µg/mL vs. 0.5 h, 67 µg/mL) - Disruption of bacterial membranes and cell clusters formation (0.5 h, MBC) | Bactericidal activity with coccoid forms being more tolerant | [43] |
| Compound | H. pylori Strain(s) | Technic Determining Morphology | Technic Determining Viability | Main Results Covering the Morphological Transformation of H. pylori | Authors Interpretation | Bibliography |
|---|---|---|---|---|---|---|
| C12K-2β12 | G27 | TEM | Culture, Fluorescence (membrane integrity) | - Viability ↓ (>2 log, 8 h, 3× MIC) - Membrane permeabilization (8 h, 3× MIC) - Ghost (degenerative) cells with coccoid-like shape, blebbing, vesicularization, and loosening of the outer membrane (16 h, 3× MIC) | Bactericidal activity | [66] |
| Epinecidin-1 | ATCC 43504 | TEM | Culture, Fluorescence (membrane integrity) | - Viability ↓ (2 log, 12 h, 2× MIC or 2 log, 24 h, MIC) - Membrane permeabilization (6 h, MIC) - Ghost (degenerative) cells with coccoid-like shape, pore formation, blebbing, vesicularization, and loosening of the outer membrane (8 h, MIC) | Bactericidal activity | [38] |
| Tilapia Piscidin 4 | ATCC 43504 | TEM | Culture, Fluorescence (membrane integrity) | - Viability ↓ (2 log, 3 h, MIC or 3 log, 3 h, 2× MIC) - Membrane permeabilization (6 h, MIC) - Ghost (degenerative) cells with coccoid-like shape, micellization, and loosening of the outer membrane (2 h, MIC) | Bactericidal activity | [39] |
| Cbf-K16 | SS1 | TEM | Culture, Fluorescence + CLSM + Flow cytometry (membrane integrity) | - Viability ↓ (0.5 log, 8 h, 4× MIC or 1 log, 12 h, 4× MIC) - Ghost (degenerative) cells with coccoid-like shape, separation of membranes, and a total loss of cytoplasmic content (8 h, 4× MIC) - Membrane permeabilization (time-dependent, 4× MIC) | Bactericidal activity | [67] |
| (PGA)m-r-(PHLG-MHH)n | SS1 | SEM | Culture, Fluorescence (membrane integrity) | - Total viability ↓ in pH = 3 (1 h, MIC) - Coccoid forms with membrane damage in pH = 3 (0.5 h, MIC) - Membrane permeabilization in pH = 3 (1 h, MIC) | Bactericidal activity in acidic environment | [68] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzyżek, P.; Grande, R. Transformation of Helicobacter pylori into Coccoid Forms as a Challenge for Research Determining Activity of Antimicrobial Substances. Pathogens 2020, 9, 184. https://doi.org/10.3390/pathogens9030184
Krzyżek P, Grande R. Transformation of Helicobacter pylori into Coccoid Forms as a Challenge for Research Determining Activity of Antimicrobial Substances. Pathogens. 2020; 9(3):184. https://doi.org/10.3390/pathogens9030184
Chicago/Turabian StyleKrzyżek, Paweł, and Rossella Grande. 2020. "Transformation of Helicobacter pylori into Coccoid Forms as a Challenge for Research Determining Activity of Antimicrobial Substances" Pathogens 9, no. 3: 184. https://doi.org/10.3390/pathogens9030184
APA StyleKrzyżek, P., & Grande, R. (2020). Transformation of Helicobacter pylori into Coccoid Forms as a Challenge for Research Determining Activity of Antimicrobial Substances. Pathogens, 9(3), 184. https://doi.org/10.3390/pathogens9030184