A Sensitive, One-Way Sequential Sieving Method to Isolate Helminths’ Eggs and Protozoal Oocysts from Lettuce for Genetic Identification

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Recovery of Parasites in Spiked Lettuce Samples
2.2. Detection of Taeniids, Toxocara spp. and Toxoplasma gondii in Lettuce Purchased in Farmer Markets and Supermarkets in Zürich
2.3. Validation of Viability Testing: In Vivo Test for E. multilocularis Eggs and by Microscopy for Toxocara Positive Samples
3. Discussion
4. Materials and Methods
4.1. Establishment and Validation of a Method for the Detection of Helminth and Protozoal Environmental Stages in Lettuce
4.1.1. Parasites Eggs and Oocysts
4.1.2. Spiking of Lettuce with Helminth and Protozoan Parasite Environmental Stages
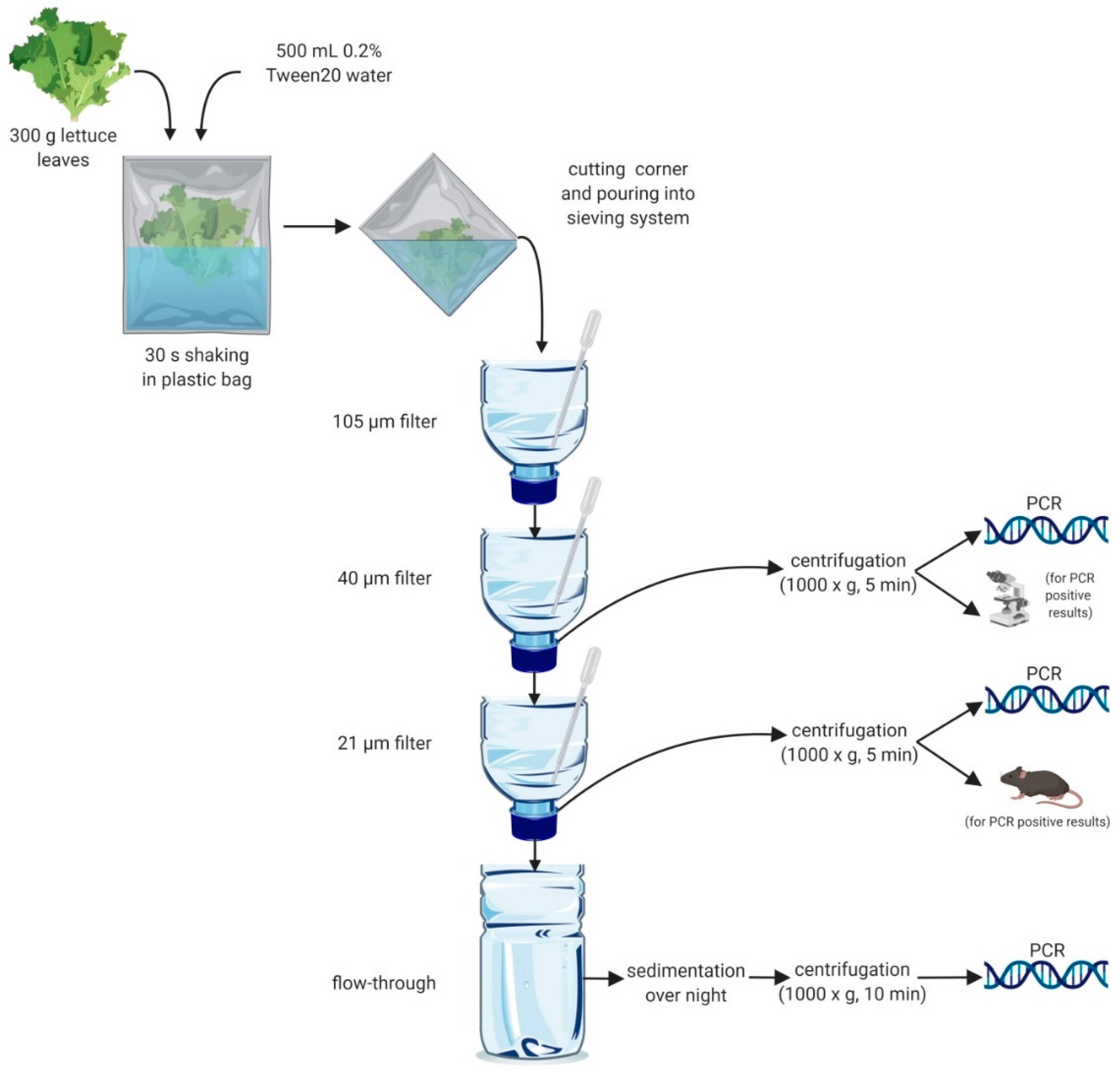
4.1.3. Sieving System for Isolation of Parasites
4.1.4. Washing of Lettuce
4.1.5. Molecular Detection of Environmental Stages of Taeniids, Toxocara and Toxoplasma
4.2. Field Study for the Assessment of Parasite Contamination of Lettuce
4.3. Assessment of Viability of Eggs
4.4. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- De Silva, N.R.; Brooker, S.; Hotez, P.J.; Montresor, A.; Engels, D.; Savioli, L. Soil-transmitted helminth infections: Updating the global picture. Trends Parasitol. 2003, 19, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Bethony, J.; Brooker, S.; Albonico, M.; Geiger, S.M.; Loukas, A.; Diemert, D.; Hotez, P.J. Soil-transmitted helminth infections: Ascariasis, trichuriasis, and hookworm. Lancet 2006, 367, 1521–1532. [Google Scholar] [CrossRef]
- Huang, D.B.; White, A.C. An updated review on cryptosporidium and giardia. Gastroenterol. Clin. N. Am. 2006, 35, 291–314. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, S.M.; Stark, D.; Harkness, J.; Ellis, J. Enteric protozoa in the developed world: A public health perspective. Clin. Microbiol. Rev. 2012, 25, 420–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prüss-Üstün, A.; Bos, R.; Gore, F.; Bartram, J. Safer Water, Better Health: Costs, Benefits and Sustainability of Interventions to Protect and Promote Health; World Health Organization: Geneva, Switzerland, 2008. [Google Scholar]
- Otranto, D.; Cantacessi, C.; Dantas-Torres, F.; Brianti, E.; Pfeffer, M.; Genchi, C.; Guberti, V.; Capelli, G.; Deplazes, P. The role of wild canids and felids in spreading parasites to dogs and cats in Europe. Part II: Helminths and arthropods. Vet. Parasitol. 2015, 213, 24–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deplazes, P.; van Knapen, F.; Schweiger, A.; Overgaauw, P.A. Role of pet dogs and cats in the transmission of helminthic zoonoses in Europe, with a focus on echinococcosis and toxocarosis. Vet. Parasitol. 2011, 182, 41–53. [Google Scholar] [CrossRef]
- Bastien, M.; Vaniscotte, A.; Combes, B.; Umhang, G.; Germain, E.; Gouley, V.; Pierlet, A.; Quintaine, T.; Forin-Wiart, M.A.; Villena, I.; et al. High density of fox and cat faeces in kitchen gardens and resulting rodent exposure to Echinococcus multilocularis and Toxoplasma gondii. Folia Parasitol. 2018, 65. [Google Scholar] [CrossRef]
- Bastien, M.; Vaniscotte, A.; Combes, B.; Umhang, G.; Raton, V.; Germain, E.; Villena, I.; Aubert, D.; Boué, F.; Poulle, M.L. Identifying drivers of fox and cat faecal deposits in kitchen gardens in order to evaluate measures for reducing contamination of fresh fruit and vegetables. Food Waterborne Parasitol. 2019, 14, e00034. [Google Scholar] [CrossRef]
- Macpherson, C.N.L.; Meslin, F.X.; Wandeler, A.I. Dogs, Zoonoses, and Public Health; CABI: Wallingford, CT, USA, 2000. [Google Scholar]
- Deplazes, P.; Hegglin, D.; Gloor, S.; Romig, T. Wilderness in the city: The urbanization of Echinococcus multilocularis. Trends Parasitol. 2004, 20, 77–84. [Google Scholar] [CrossRef]
- Liccioli, S.; Giraudoux, P.; Deplazes, P.; Massolo, A. Wilderness in the ‘city’ revisited: Different urbes shape transmission of Echinococcus multilocularis by altering predator and prey communities. Trends Parasitol. 2015, 31, 297–305. [Google Scholar] [CrossRef]
- WHO. Waterborne Zoonoses: Identification, Causes and Control; IWA Publishing: East Sussex, UK, 2004. [Google Scholar]
- Kern, P.; Ammon, A.; Kron, M.; Sinn, G.; Sander, S.; Petersen, L.R.; Gaus, W.; Kern, P. Risk factors for alveolar echinococcosis in humans. Emerg. Infect. Dis. 2004, 10, 2088. [Google Scholar] [CrossRef] [PubMed]
- Kapperud, G.; Jenum, P.A.; Stray-Pedersen, B.; Melby, K.K.; Eskild, A.; Eng, J. Risk factors for Toxoplasma gondii infection in pregnancy. Results of a prospective case-control study in Norway. Am. J. Epidemiol. 1996, 144, 405–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, A.J.; Gilbert, R.E.; Buffolano, W.; Zufferey, J.; Petersen, E.; Jenum, P.A.; Foulon, W.; Semprini, A.E.; Dunn, D.T. Sources of toxoplasma infection in pregnant women: European multicentre case-control study. European Research Network on Congenital Toxoplasmosis. BMJ 2000, 321, 142–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.L.; Dargelas, V.; Roberts, J.; Press, C.; Remington, J.S.; Montoya, J.G. Risk factors for Toxoplasma gondii infection in the United States. Clin. Infect. Dis. 2009, 49, 878–884. [Google Scholar] [CrossRef] [Green Version]
- Maurelli, M.P.; Santaniello, A.; Fioretti, A.; Cringoli, G.; Rinaldi, L.; Menna, L.F. The presence of Toxocara eggs on dog’s fur as potential zoonotic risk in animal-assisted interventions: A systematic review. Animals 2019, 9, 827. [Google Scholar] [CrossRef] [Green Version]
- FAO/WHO. Multicriteria-Based Ranking for Risk Management of Food-Borne Parasites; Food and Agriculture Organization of the United Nations (FAO)/World Health Organization (WHO): Rome, Italy, 2014; p. 302. [Google Scholar]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; Cesare, A.; Herman, L.; Hilbert, F.; et al. Public health risks associated with food-borne parasites. EFSA J. 2018, 16. [Google Scholar] [CrossRef]
- Alvarez Rojas, C.A.; Mathis, A.; Deplazes, P. Assessing the contamination of food and the environment with Taenia and Echinococcus eggs and their zoonotic transmission. Curr. Clin. Microbiol. Rep. 2018, 5, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Tamarozzi, F.; Deplazes, P.; Casulli, A. Reinventing the wheel of Echinococcus granulosus sensu lato transmission to humans. Trends Parasitol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Robertson, L.J.; Troell, K.; Woolsey, I.D.; Kapel, C.M. Fresh fruit, vegetables, and mushrooms as transmission vehicles for Echinococcus multilocularis in Europe: Inferences and concerns from sample analysis data from Poland. Parasitol. Res. 2016, 115, 2485–2488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousseau, A.; La Carbona, S.; Dumètre, A.; Robertson, L.J.; Gargala, G.; Escotte-Binet, S.; Favennec, L.; Villena, I.; Gérard, C.; Aubert, D. Assessing viability and infectivity of foodborne and waterborne stages (cysts/oocysts) of Giardia duodenalis, Cryptosporidium spp., and Toxoplasma gondii: A review of methods. Parasite 2018, 25, 14. [Google Scholar] [CrossRef] [Green Version]
- Federer, K.; Armua-Fernandez, M.T.; Hoby, S.; Wenker, C.; Deplazes, P. In vivo viability of Echinococcus multilocularis eggs in a rodent model after different thermo-treatments. Exp. Parasitol. 2015, 154, 14–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trachsel, D.; Deplazes, P.; Mathis, A. Identification of taeniid eggs in the faeces from carnivores based on multiplex PCR using targets in mitochondrial DNA. Parasitology 2007, 134, 911–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burg, J.L.; Grover, C.M.; Pouletty, P.; Boothroyd, J.C. Direct and sensitive detection of a pathogenic protozoan, Toxoplasma gondii, by polymerase chain reaction. J. Clin. Microbiol. 1989, 27, 1787–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathis, A.; Deplazes, P.; Eckert, J. An improved test system for PCR-based specific detection of Echinococcus multilocularis eggs. J. Helminthol. 1996, 70, 219–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federer, K.; Armua-Fernandez, M.T.; Gori, F.; Hoby, S.; Wenker, C.; Deplazes, P. Detection of taeniid (Taenia spp., Echinococcus spp.) eggs contaminating vegetables and fruits sold in European markets and the risk for metacestode infections in captive primates. Int. J. Parasitol. Parasites Wildl. 2016, 5, 249–253. [Google Scholar] [CrossRef] [Green Version]
- Adler, F.R.; Green, A.M.; Şekercioğlu, Ç.H. Citizen science in ecology: A place for humans in nature. Ann. N. Y. Acad. Sci. 2020, 26. [Google Scholar] [CrossRef]
- Breen, N.; Berrigan, D.; Jackson, J.S.; Wong, D.W.S.; Wood, F.B.; Denny, J.C.; Zhang, X.; Bourne, P.E. Translational health disparities research in a data-rich world. Health Equity 2019, 3, 588–600. [Google Scholar] [CrossRef]
- Frey, C.F.; Oakley, J.R.; Lobanov, V.A.; Marreros, N.; Schurer, J.M.; Lalonde, L.F. A novel protocol to isolate, detect and differentiate taeniid eggs in leafy greens and berries using real-time PCR with melting curve analysis. Parasites Vectors 2019, 12, 590. [Google Scholar] [CrossRef]
- Malkamäki, S.; Näreaho, A.; Lavikainen, A.; Oksanen, A.; Sukura, A. A new SYBR green real-time PCR assay for semi-quantitative detection of Echinococcus multilocularis and Echinococcus canadensis DNA on bilberries (Vaccinium myrtillus). Food Waterborne Parasitol. 2019, 17, e00068. [Google Scholar] [CrossRef]
- Temesgen, T.T.; Robertson, L.J.; Tysnes, K.R. A novel multiplex real-time PCR for the detection of Echinococcus multilocularis, Toxoplasma gondii, and Cyclospora cayetanensis on berries. Food Res. Int. 2019, 125, 108636. [Google Scholar] [CrossRef]
- Lass, A.; Szostakowska, B.; Myjak, P.; Korzeniewski, K. The first detection of Echinococcus multilocularis DNA in environmental fruit, vegetable, and mushroom samples using nested PCR. Parasitol. Res. 2015, 114, 4023–4029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchioro, A.A.; Tiyo, B.T.; Colli, C.M.; de Souza, C.Z.; Garcia, J.L.; Gomes, M.L.; Falavigna-Guilherme, A.L. First detection of Toxoplasma gondii DNA in the fresh leafs of vegetables in South America. Vector Borne Zoonotic Dis. 2016, 16, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Hohweyer, J.; Cazeaux, C.; Travaillé, E.; Languet, E.; Dumètre, A.; Aubert, D.; Terryn, C.; Dubey, J.P.; Azas, N.; Houssin, M.; et al. Simultaneous detection of the protozoan parasites Toxoplasma, Cryptosporidium and Giardia in food matrices and their persistence on basil leaves. Food Microbiol. 2016, 57, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Lass, A.; Pietkiewicz, H.; Szostakowska, B.; Myjak, P. The first detection of Toxoplasma gondii DNA in environmental fruits and vegetables samples. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1101–1108. [Google Scholar] [CrossRef] [Green Version]
- Reischl, U.; Bretagne, S.; Krüger, D.; Ernault, P.; Costa, J.M. Comparison of two DNA targets for the diagnosis of Toxoplasmosis by real-time PCR using fluorescence resonance energy transfer hybridization probes. BMC Infect. Dis. 2003, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Deplazes, P.; Eichenberger, R.M.; Grimm, F. Wildlife-transmitted Taenia and Versteria cysticercosis and coenurosis in humans and other primates. Int. J. Parasitol. Parasites Wildl. 2019, 9, 342–358. [Google Scholar] [CrossRef]
- Hofer, S.; Gloor, S.; MÜLler, U.; Mathis, A.; Hegglin, D.; Deplazes, P. High prevalence of Echinococcus multilocularis in urban red foxes (Vulpes vulpes) and voles (Arvicola terrestris) in the city of Zürich, Switzerland. Parasitology 2000, 120, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Brossard, M.; Andreutti, C.; Siegenthaler, M. Infection of red foxes with Echinococcus multilocularis in western Switzerland. J. Helminthol. 2007, 81, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Lalonde, L.F.; Gajadhar, A.A. Detection of Cyclospora cayetanensis, Cryptosporidium spp., and Toxoplasma gondii on imported leafy green vegetables in Canadian survey. Food Waterborne Parasitol. 2016, 2, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Bethea, D.A. Detection of Toxoplasma Gondii in Fresh Produce; University of Georgia: Athens, GA, USA, 2014. [Google Scholar]
- Ahmad, S.O.; El Fadaly, H.; Zaki, M.S.; Barakat, A. Incidence of zoonotic parasites in Egyptian raw vegetable salads. Life Sci. J. 2016, 13, 27–31. [Google Scholar]
- Al-Megrm, W.I. Prevalence of intestinal parasites in leafy vegetables in Riyadh, Saudi Arabia. Int. J. Trop. Med. 2010, 5, 20–23. [Google Scholar]
- Tschanz, B.; Hegglin, D.; Gloor, S.; Bontadina, F. Hunters and non-hunters: Skewed predation rate by domestic cats in a rural village. Eur. J. Wildl. Res. 2011, 57, 597–602. [Google Scholar] [CrossRef]
- Frey, C.; Berger-Schoch, A.; Herrmann, D.; Schares, G.; Müller, N.; Bernet, D.; Doherr, M.; Gottstein, B. Incidence and genotypes of Toxoplasma gondii in the muscle of sheep, cattle, pigs as well as in cat feces in Switzerland. Schweiz. Arch. Tierheilkd. 2012, 154, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Zottler, E.M.; Bieri, M.; Basso, W.; Schnyder, M. Intestinal parasites and lungworms in stray, shelter and privately owned cats of Switzerland. Parasitol. Int. 2019, 69, 75–81. [Google Scholar] [CrossRef]
- Moazeni, M.; Rakhshandehroo, E. In vitro viability test for the eggs of Echinococcus granulosus: A rapid method. Parasitol. Res. 2012, 110, 925–930. [Google Scholar] [CrossRef]
- Eckert, J.; Deplazes, P.; Craig, P.; Gemmell, M.A.; Gottstein, B.; Heath, D.; Jenkins, D.; Kamiya, M.; Lightowlers, M.W. Echinococcosis in animals: Clinical aspects, diagnosis and treatment. In WHO/OIE Manual on Echinococcosis in Humans and Animals: A Public Health Problem of Global Concern; Eckert, J., Gemmell, M.A., Meslin, F.X., Pawłowski, Z.S., Eds.; World Organisation for Animal Health: Paris, France, 2001. [Google Scholar]
- Deplazes, P.; Eckert, J. Mass collection and storage of Taenia hydatigena eggs and isolation of viable oncospheres. Schweiz. Arch. Tierheilkd. 1988, 130, 307–320. [Google Scholar]
- Fahrion, A.S.; Staebler, S.; Deplazes, P. Patent Toxocara canis infections in previously exposed and in helminth-free dogs after infection with low numbers of embryonated eggs. Vet. Parasitol. 2008, 152, 108–115. [Google Scholar] [CrossRef]
- Stefanić, S.; Shaikenov, B.S.; Deplazes, P.; Dinkel, A.; Torgerson, P.R.; Mathis, A. Polymerase chain reaction for detection of patent infections of Echinococcus granulosus (“sheep strain”) in naturally infected dogs. Parasitol. Res. 2004, 92, 347–351. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Environmental Stages Used for Spiking | |||||
|---|---|---|---|---|---|
| Embryonated Toxocara Eggs | Echinococcus multilocularis and Taenia polyacantha Eggs Containing Sodium Hypochlorate Resistant (SHR) Oncospheres | ||||
| # Eggs | PCR 1 | Microscopy | # Eggs | PCR 2 E. multilocularis | PCR 2 Taenia |
| 20 | 5/5 | 5/5 | 20 | 5/5 | 2/5 |
| 4 | 5/5 | 2/5 | 4 | 5/5 | 3/5 |
| 0 | 0/3 | 0/3 | 0 | 0/3 | 0/3 |
| Environmental Stages Used for Spiking | ||||
|---|---|---|---|---|
| Echinococcus multilocularis Eggs Containing SHR Oncospheres | Toxoplasma gondii Oocysts | |||
| # Eggs (Volume of Spiking Suspension) | PCR 1 1st half | PCR 2nd half | # Oocysts | PCR 2 |
| 120 (779 µL) | 5/5 | 86 | 5/5 | |
| 40 (260 µL) | 5/5 | 18 | 4/5 | |
| 20 (130 µL) | 5/5 | 0 | 0/3 | |
| 10 (65 µL) | 5/5 | 5/5 | ||
| 2 3 (20 µL) | 5/5 | 1/5 | ||
| 0 (none) | 0/3 | 0/3 | ||
| 0 (779 µL 4) | 0/3 | - | ||
| 0 (130 µL 4) | 0/3 | - | ||
| Location Purchased | Season | Parasite Species | Number of PCR 1 Positives |
|---|---|---|---|
| Farmer markets (n = 120) | Summer (n = 57) | Hydatigera taeniaeformis (Syn. Taenia taeniaeformis) | 1 |
| T. martis | 1 | ||
| T. polyacantha | 2 | ||
| Toxocara cati | 2 | ||
| Autumn (n = 63) | Echinococcus multilocularis | 2 | |
| Hydatigera taeniaeformis | 3 | ||
| T. polyacantha | 1 | ||
| Toxocara cati | 1 | ||
| Total:13 (10.8%, 95% CI 5.2 to 16.4) | |||
| Supermarkets (n = 37) | Summer (n = 23) | T. cati | 1 |
| Autumn (n = 14) | - | 0 | |
| Total: 1 (2.7%, 95% CI −2.8 to 8.2) |
| Location Purchased | Season | Number of PCR 1 Positives |
|---|---|---|
| Farmer markets (n = 76) | Summer (n = 15) | 1 |
| Autumn (n = 61) | 3 | |
| Supermarkets (n = 24) | Summer (n = 10) | 0 |
| Autumn (n = 14) | 2 |
| Egg Suspensions | Echinococcus multilocularis and Taenia polyacantha Eggs (Inoculum 1) | E. multilocularis Eggs (Inoculum 2) | ||
|---|---|---|---|---|
| # Eggs | In Vivo Development 1 | # Eggs | In Vivo Development 1 | |
| Inoculum | 40 | 5/5 | 60 | 5/5 |
| 10 | 2/5 | 20 | 2/5 | |
| 2 | 2/5 | 10 | 1/5 | |
| Recovered after spiking/washing | 20 | 0/5 | 120 | 1/5 |
| 4 | 1/5 | 40 | 0/5 | |
| - | - | 20 | 1/5 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guggisberg, A.R.; Alvarez Rojas, C.A.; Kronenberg, P.A.; Miranda, N.; Deplazes, P. A Sensitive, One-Way Sequential Sieving Method to Isolate Helminths’ Eggs and Protozoal Oocysts from Lettuce for Genetic Identification. Pathogens 2020, 9, 624. https://doi.org/10.3390/pathogens9080624
Guggisberg AR, Alvarez Rojas CA, Kronenberg PA, Miranda N, Deplazes P. A Sensitive, One-Way Sequential Sieving Method to Isolate Helminths’ Eggs and Protozoal Oocysts from Lettuce for Genetic Identification. Pathogens. 2020; 9(8):624. https://doi.org/10.3390/pathogens9080624
Chicago/Turabian StyleGuggisberg, Annina R., Cristian A. Alvarez Rojas, Philipp A. Kronenberg, Nadia Miranda, and Peter Deplazes. 2020. "A Sensitive, One-Way Sequential Sieving Method to Isolate Helminths’ Eggs and Protozoal Oocysts from Lettuce for Genetic Identification" Pathogens 9, no. 8: 624. https://doi.org/10.3390/pathogens9080624
APA StyleGuggisberg, A. R., Alvarez Rojas, C. A., Kronenberg, P. A., Miranda, N., & Deplazes, P. (2020). A Sensitive, One-Way Sequential Sieving Method to Isolate Helminths’ Eggs and Protozoal Oocysts from Lettuce for Genetic Identification. Pathogens, 9(8), 624. https://doi.org/10.3390/pathogens9080624