
Allium-Based Phytobiotic for Laying Hens’ Supplementation: Effects on Productivity, Egg Quality, and Fecal Microbiota

 , , ,
, , ,  , ,
, ,
Abstract
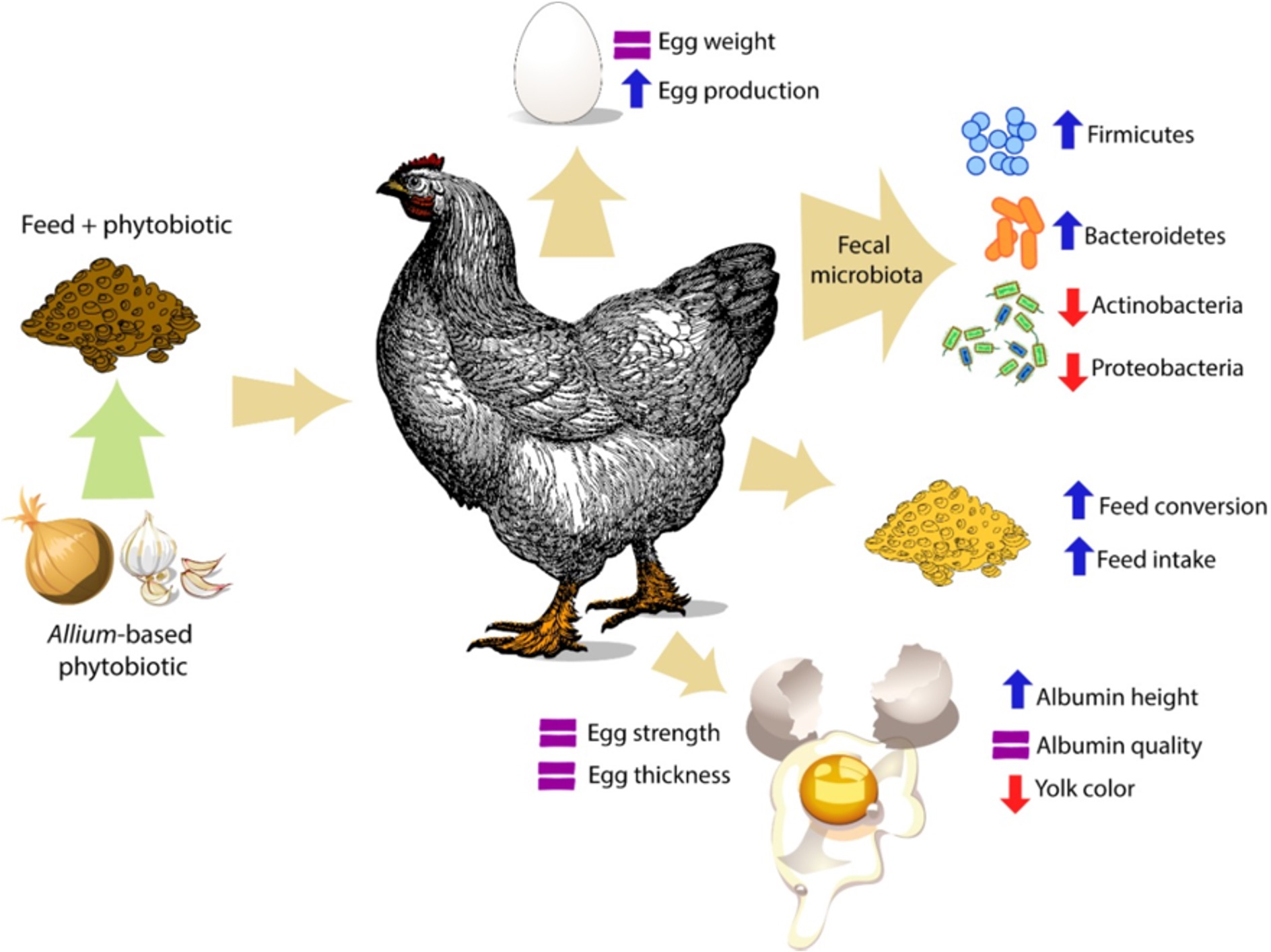
:
1. Introduction
2. Materials and Methods
2.1. Phytobiotic Mixture
2.2. Hens Housing and Feeding
2.3. Salmonella Infection
2.4. Feed Intake, Productivity and Egg Quality
2.5. Fecal-Associated Bacterial Communities
2.5.1. Sampling and DNA Extraction
2.5.2. Preparation of Libraries and Sequencing Procedure
2.5.3. Bioinformatics
2.5.4. Experimental Design
3. Results
3.1. Hens’ Productivity
3.2. Egg Quality
3.3. Fecal-Associated Bacterial Communities’ Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yamashiro, Y. Gut Microbiota in Health and Disease. Ann. Nutr. Metab. 2018, 71, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Kraimi, N.; Dawkins, M.; Gebhardt-Henrich, S.G.; Velge, P.; Rychlik, I.; Volf, J.; Creach, P.; Smith, A.; Colles, F.; Leterrier, C. Influence of the Microbiota-Gut-Brain Axis on Behavior and Welfare in Farm Animals: A Review. Physiol. Behav. 2019, 210, 112658. [Google Scholar] [CrossRef] [PubMed]
- Guitton, E.; Faurie, A.; Lavillatte, S.; Chaumeil, T.; Gaboriaud, P.; Bussière, F.; Laurent, F.; Lacroix-Lamandé, S.; Guabiraba, R.; Schouler, C. Production of Germ-Free Fast-Growing Broilers from a Commercial Line for Microbiota Studies. J. Vis. Exp. 2020, 18, e61148. [Google Scholar] [CrossRef]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The Role of Short-Chain Fatty Acids in the Interplay between Diet, Gut Microbiota, and Host Energy Metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azad, M.A.K.; Gao, J.; Ma, J.; Li, T.; Tan, B.; Huang, X.; Yin, J. Opportunities of Prebiotics for the Intestinal Health of Monogastric Animals. Anim. Nutr. 2020, 6, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.Y.; Groer, M.; Dutra, S.V.O.; Sarkar, A.; McSkimming, D.I. Gut Microbiota and Immune System Interactions. Microorganisms 2020, 8, 1587. [Google Scholar] [CrossRef]
- Khan, S.; Moore, R.J.; Stanley, D.; Chousalkar, K.K. The Gut Microbiota of Laying Hens and Its Manipulation with Prebiotics and Probiotics to Enhance Gut Health and Food Safety. Appl. Environ. Microbiol. 2020, 86, e00600-20. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, L.; Sun, X.; Wan, X.; Sun, G.; Jiang, R.; Li, W.; Tian, Y.; Liu, X.; Kang, X. Characteristics of the Fecal Microbiota of High-and Low-Yield Hens and Effects of Fecal Microbiota Transplantation on Egg Production Performance. Res. Vet. Sci. 2020, 129, 164–173. [Google Scholar] [CrossRef] [PubMed]
- Skoufos, I.; Bonos, E.; Anastasiou, I.; Tsinas, A.; Tzora, A. Effects of Phytobiotics in Healthy or Disease Challenged Animals. Feed Addit. 2020, 18, 311–337. [Google Scholar]
- Yıldız, A.Ö.; Şentürk, E.T.; Olgun, O. Use of Alfalfa Meal in Layer Diets a Review. World’s Poult. Sci. J. 2020, 76, 134–143. [Google Scholar] [CrossRef]
- Avila-Sosa, R.; Navarro-Cruz, A.R.; Sosa-Morales, M.E.; López-Malo, A.; Palou, E. Bergamot (Citrus Bergamia) Oils; Preedy, V.R., Ed.; Academic Press: San Diego, CA, USA, 2016; Volume 1, Chapter 27; pp. 247–252. [Google Scholar]
- Abdel-Wareth, A.A.A.; Lohakare, J.D. Effect of Dietary Supplementation of Peppermint on Performance, Egg Quality, and Serum Metabolic Profile of Hy-Line Brown Hens during the Late Laying Period. Anim. Feed Sci. Technol. 2014, 197, 114–120. [Google Scholar] [CrossRef]
- Sadarman; Febrina, D.; Yendraliza; Haq, M.S.; Nurfitriani, R.A.; Barkah, N.N.; Sholikin, M.M.; Yunilas; Qomariyah, N.; Jayanegara, A.; et al. Effect of Dietary Black Cumin Seed (Nigella Sativa) on Performance, Immune Status, and Serum Metabolites of Small Ruminants: A Meta-Analysis. Small Rumin. Res. 2021, 204, 106521. [Google Scholar] [CrossRef]
- Vicente, J.L.; Lopez, C.; Avila, E.; Morales, E.; Hargis, B.M.; Tellez, G. Effect of Dietary Natural Capsaicin on Experimental Salmonella Enteritidis Infection and Yolk Pigmentation in Laying Hens. Int. J. Poult. Sci. 2007, 6, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Alizadeh, M.R.; Mahdavi, A.H.; Rahmani, H.R.; Jahanian, E. Effects of Different Levels of Clove Bud (Syzygium Aromaticum) on Yolk Biochemical Parameters and Fatty Acids Profile, Yolk Oxidative Stability, and Ovarian Follicle Numbers of Laying Hens Receiving Different n-6 to n-3 Ratios. Anim. Feed Sci. Technol. 2015, 206, 67–75. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Peng, Q.Y.; Liu, Y.R.; Ma, Q.G.; Zhang, J.Y.; Guo, Y.P.; Xue, Z.; Zhao, L.H. Effects of Oregano Essential Oil as an Antibiotic Growth Promoter Alternative on Growth Performance, Antioxidant Status, and Intestinal Health of Broilers. Poult. Sci. 2021, 100, 101163. [Google Scholar] [CrossRef]
- Dosoky, W.M.; Zeweil, H.S.; Ahmed, M.H.; Zahran, S.M.; Shaalan, M.M.; Abdelsalam, N.R.; Abdel-Moneim, A.-M.E.; Taha, A.E.; El-Tarabily, K.A.; Abd El-Hack, M.E. Impacts of Onion and Cinnamon Supplementation as Natural Additives on the Performance, Egg Quality and Immunity in Laying Japanese Quail. Poult. Sci. 2021, 100, 101482. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Miyata, G. The Nutraceutical Benefit, Part IV: Garlic. Nutrition 2000, 16, 787–788. [Google Scholar] [CrossRef]
- Alagawany, M.; Elnesr, S.S.; Farag, M.R.; Abd El-Hack, M.E.; Barkat, R.A.; Gabr, A.A.; Foda, M.A.; Noreldin, A.E.; Khafaga, A.F.; El-Sabrout, K.; et al. Potential Role of Important Nutraceuticals in Poultry Performance and Health-A Comprehensive Review. Res. Vet. Sci. 2021, 137, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Dal Pont, G.C.; Farnell, M.B.; Jarvis, S.; Battaglia, M.; Arsenault, R.J.; Kogut, M.H. Supplementing Chestnut Tannins in the Broiler Diet Mediates a Metabolic Phenotype of the Ceca. Poult. Sci. 2021, 100, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.J.; Kuo, C.M.; Chuang, Y.C.; Hong, J.W.; Chou, R.L.; Chen, T.I. The Effects of Garlic-Supplemented Diets on Antibacterial Activity against Streptococcus Iniae and on Growth in Orange-Spotted Grouper, Epinephelus Coioides. Aquaculture 2012, 364, 33–38. [Google Scholar] [CrossRef]
- Lebdah, M.; Tantawy, L.; Elgamal, A.M.; Abdelaziz, A.M.; Yehia, N.; Alyamani, A.A.; ALmoshadak, A.S.; Mohamed, M.E. The Natural Antiviral and Immune Stimulant Effects of Allium Cepa Essential Oil Onion Extract against Virulent Newcastle Disease Virus. Saudi J. Biol. Sci. 2021, in press. [CrossRef]
- Lestari, S.R.; Christina, Y.I.; Athóillah, M.F.; Rifái, M. Single-Bulb Garlic Oil Regulates Toll-like Receptors and Nrf2 Cross-Talk and IL-17 Production in Mice Fed with High-Fat Diet. Saudi J. Biol. Sci. 2021, 11, 6515–6522. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Deng, P.; Xu, Y.; Lu, S.; Wang, J. Quantification and Analysis of Anthocyanin and Flavonoids Compositions, and Antioxidant Activities in Onions with Three Different Colors. J. Integr. Agric. 2016, 15, 2175–2181. [Google Scholar] [CrossRef] [Green Version]
- Shalini, R.; Krishna, J.; Sankaranarayanan, M.; Antony, U. Enhancement of Fructan Extraction from Garlic and Fructooligosaccharide Purification Using an Activated Charcoal Column. LWT 2021, 148, 111703. [Google Scholar] [CrossRef]
- Martins, N.; Petropoulos, S.; Ferreira, I.C.F.R. Chemical Composition and Bioactive Compounds of Garlic (Allium sativum L.) as Affected by Pre- and Post-Harvest Conditions: A Review. Food Chem. 2016, 211, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Author Correction: Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 1091. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for Prediction of Metagenome Functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Caspi, R.; Billington, R.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Midford, P.E.; Ong, W.K.; Paley, S.; Subhraveti, P.; Karp, P.D. The MetaCyc Database of Metabolic Pathways and Enzymes-a 2019 Update. Nucleic Acids Res. 2020, 48, 445–453. [Google Scholar] [CrossRef] [Green Version]
- Putnik, P.; Gabrić, D.; Roohinejad, S.; Barba, F.J.; Granato, D.; Mallikarjunan, K.; Lorenzo, J.M.; Bursać Kovačević, D. An Overview of Organosulfur Compounds from Allium Spp.: From Processing and Preservation to Evaluation of Their Bioavailability, Antimicrobial, and Anti-Inflammatory Properties. Food Chem. 2019, 276, 680–691. [Google Scholar] [CrossRef]
- Aljumaah, M.R.; Suliman, G.M.; Abdullatif, A.A.; Abudabos, A.M. Effects of Phytobiotic Feed Additives on Growth Traits, Blood Biochemistry, and Meat Characteristics of Broiler Chickens Exposed to Salmonella Typhimurium. Poult. Sci. 2020, 99, 5744–5751. [Google Scholar] [CrossRef] [PubMed]
- Ogbuewu, I.P.; Okoro, V.M.; Mbajiorgu, C.A. Meta-Analysis of the Responses of Laying Hens to Garlic (Allium sativum) Supplementation. Anim. Feed Sci. Technol. 2021, 275, 114866. [Google Scholar] [CrossRef]
- Damaziak, K.; Riedel, J.; Gozdowski, D.; Niemiec, J.; Siennicka, A.; Róg, D. Productive Performance and Egg Quality of Laying Hens Fed Diets Supplemented with Garlic and Onion Extracts. J. Appl. Poult. Res. 2017, 26, 337–349. [Google Scholar] [CrossRef]
- Ayed, M.; Aïssa, A.; Noumi, M. A Comparative Study between the Effects of Feed Inclusion with Garlic (Allium sativum), Cloves and Turmeric (Curcuma longa) Rhizome Powder on Laying Hen Performance and Egg Quality. Iran. J. Appl. Anim. Sci. 2018, 8, 693–701. [Google Scholar]
- Gilani, S.M.H.; Rashid, Z.; Galani, S.; Ilyas, S.; Sahar, S.; Zahoor-ul-Hassan; Al-Ghanim, K.; Zehra, S.; Azhar, A.; Al-Misned, F.; et al. Growth Performance, Intestinal Histomorphology, Gut Microflora and Ghrelin Gene Expression Analysis of Broiler by Supplementing Natural Growth Promoters: A Nutrigenomics Approach. Saudi J. Biol. Sci. 2021, 28, 3438–3447. [Google Scholar] [CrossRef]
- Adaszyńska-Skwirzyńska, M.; Szczerbińska, D. The Effect of Lavender (Lavandula angustifolia) Essential Oil as a Drinking Water Supplement on the Production Performance, Blood Biochemical Parameters, and Ileal Microflora in Broiler Chickens. Poult. Sci. 2019, 98, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Abouelezz, K.; Abou-Hadied, M.; Yuan, J.; Elokil, A.A.; Wang, G.; Wang, S.; Wang, J.; Bian, G. Nutritional Impacts of Dietary Oregano and Enviva Essential Oils on the Performance, Gut Microbiota and Blood Biochemicals of Growing Ducks. Animal 2019, 13, 2216–2222. [Google Scholar] [CrossRef] [PubMed]
- Rodjan, P.; Wattanasit, S.; Faroongsarng, D.; Thongprajukaew, K.; Theapparat, Y. Garlic Oil Granules Coated with Enteric Polymer: Effects on Performance, Egg Quality, Yolk Antioxidants, Yolk Cholesterol, Blood Biochemistry and Hepatic Lipid Metabolism in Laying Hens. Anim. Feed Sci. Technol. 2021, 274, 114854. [Google Scholar] [CrossRef]
- Oladipo Kolawole, F.S.; Folake, A.O. Egg Traits and Productive Performance of Isa-Brown Laying Hens Fed Garlic Supplemented Diets. Asian J. Appl. Sci. 2019, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Nemati, Z.; Moradi, Z.; Alirezalu, K.; Besharati, M.; Raposo, A. Impact of Ginger Root Powder Dietary Supplement on Productive Performance, Egg Quality, Antioxidant Status and Blood Parameters in Laying Japanese Quails. Int. J. Environ. Res. Public Health 2021, 18, 2995. [Google Scholar] [CrossRef] [PubMed]
- Świątkiewicz, S.; Arczewska-Włosek, A.; Krawczyk, J.; Szczurek, W.; Puchała, M.; Józefiak, D. Effect of Selected Feed Additives on Egg Performance and Eggshell Quality in Laying Hens Fed a Diet with Standard or Decreased Calcium Content. Ann. Anim. Sci. 2018, 18, 167–183. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, K.Z.; Gharaibeh, S.M.; Zakaria, H.A.; Qatramiz, A.M. Garlic (Allium sativum) Supplementation: Influence on Egg Production, Quality, and Yolk Cholesterol Level in Layer Hens. Asian-Australas J. Anim. Sci. 2010, 23, 1503–1509. [Google Scholar] [CrossRef]
- Ghasemi, R.; Zarei, M.; Torki, M. Adding Medicinal Herbs Including Garlic (Allium sativum) and Thyme (Thymus vulgaris) to Diet of Laying Hens and Evaluating Productive Performance and Egg Quality Characteristics. Am. J. Anim. Vet. Sci. 2010, 5, 151–154. [Google Scholar] [CrossRef]
- Gong, H.Z.; Lang, W.Y.; Lan, H.N.; Fan, Y.Y.; Wang, T.P.; Chu, Q.R.; Wang, J.H.; Li, D.; Zheng, X.; Wu, M. Effects of Laying Breeder Hens Dietary β-Carotene, Curcumin, Allicin, and Sodium Butyrate Supplementation on the Jejunal Microbiota and Immune Response of Their Offspring Chicks. Poult. Sci. 2020, 99, 3807–3816. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Han, C.; Li, S.; Geng, Y.; Wei, Y.; Shi, W.; Bao, Y. High-Throughput Sequencing–Based Analysis of the Intestinal Microbiota of Broiler Chickens Fed with Compound Small Peptides of Chinese Medicine. Poult. Sci. 2021, 100, 100897. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Rehman, M.U.; Huang, S.; Jiang, X.; Zhang, H.; Li, J. Comparative Analysis of Gut Microbial Community in Healthy and Tibial Dyschondroplasia Affected Chickens by High Throughput Sequencing. Microb. Pathog. 2018, 118, 133–139. [Google Scholar] [CrossRef]
- van der Eijk, J.A.J.; de Vries, H.; Kjaer, J.B.; Naguib, M.; Kemp, B.; Smidt, H.; Rodenburg, T.B.; Lammers, A. Differences in Gut Microbiota Composition of Laying Hen Lines Divergently Selected on Feather Pecking. Poult. Sci. 2019, 98, 7009–7021. [Google Scholar] [CrossRef]
- Quinteros, J.A.; Scott, P.C.; Wilson, T.B.; Anwar, A.M.; Scott, T.; Muralidharan, C.; Van, T.T.H.; Moore, R.J. Isoquinoline Alkaloids Induce Partial Protection of Laying Hens from the Impact of Campylobacter Hepaticus (Spotty Liver Disease) Challenge. Poult. Sci. 2021, 100, 101423. [Google Scholar] [CrossRef]
- Aljumaah, M.R.; Alkhulaifi, M.M.; Aljumaah, R.S.; Abudabos, A.M.; Abdullatif, A.A.; Suliman, G.M.; Al-Ghadi, M.Q.; Stanley, D. Influence of Sanguinarine-Based Phytobiotic Supplementation on Post Necrotic Enteritis Challenge Recovery. Heliyon 2020, 6, 05361. [Google Scholar] [CrossRef]
- Chaves Hernández, A.J. Poultry and Avian Diseases. In Encyclopedia of Agriculture and Food Systems; Van Alfen, N.K., Ed.; Elsevier: Cambridge, MA, USA, 2014; pp. 504–520. [Google Scholar]
- Kouam, M.K.; Biekop, H.M.F.; Katte, B.; Teguia, A. Risk Factors of Salmonella Infection in Laying Hens in Menoua Division, Western Region of Cameroon (Central Africa). Comp. Immunol. Microbiol. Infect. Dis. 2019, 67, 101370. [Google Scholar] [CrossRef] [PubMed]
- Salem, W.M.; Shibat El-hamed, D.M.W.; Sayed, W.F.; Elamary, R.B. Alterations in Virulence and Antibiotic Resistant Genes of Multidrug-Resistant Salmonella Serovars Isolated from Poultry: The Bactericidal Efficacy of Allium Sativum. Microb. Pathog. 2017, 108, 91–100. [Google Scholar] [CrossRef]
- Pirgozliev, V.; Bravo, D.; Mirza, M.W.; Rose, S.P. Growth Performance and Endogenous Losses of Broilers Fed Wheat-Based Diets with and without Essential Oils and Xylanase Supplementation. Poult. Sci. 2015, 94, 1227–1232. [Google Scholar] [CrossRef] [PubMed]
- Agbai, K.N.; Omage, J.J.; Sekoni, A.A.; Afolayan, M. Effect of Different Levels of Biotronic® Top Forte (Commercial Blended Organic Acids) as a Replacement for Antibiotic Growth Promoter on the Growth Performance and Villi Morphology of Pullet Chicks (0–8 weeks). Niger. J. Anim. Sci. Technol. (NJAST) 2020, 3, 13–21. [Google Scholar]
- Hameed, H. Feed Additives in Poultry. Assiut Vet. Med. J. 2021, 67, 87–100. [Google Scholar]
- Abudabos, A.M.; Hussein, E.O.S.; Ali, M.H.; Al-Ghadi, M.Q. The Effect of Some Natural Alternative to Antibiotics on Growth and Changes in Intestinal Histology in Broiler Exposed to Salmonellachallenge. Poult. Sci. 2019, 98, 1441–1446. [Google Scholar] [CrossRef] [PubMed]
- Salah, A.S.; Ahmed-Farid, O.A.; El-Tarabany, M.S. Effects of Guanidinoacetic Acid Supplements on Laying Performance, Egg Quality, Liver Nitric Oxide and Energy Metabolism in Laying Hens at the Late Stage of Production. J. Agric. Sci. 2020, 158, 241–246. [Google Scholar] [CrossRef]
- Han, Y.-K.; Thacker, P.A. Influence of Energy Level and Glycine Supplementation on Performance, Nutrient Digestibility and Egg Quality in Laying Hens. Asian-Australas J. Anim. Sci. 2011, 24, 1447–1455. [Google Scholar] [CrossRef]
- Stampfli, A.R.; Blankenfeldt, W.; Seebeck, F.P. Structural Basis of Ergothioneine Biosynthesis. Curr. Opin. Struct. Biol. 2020, 65, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Abdalkarim Salih, S.; Daghigh-Kia, H.; Mehdipour, M.; Najafi, A. Does Ergothioneine and Thawing Temperatures Improve Rooster Semen Post-Thawed Quality? Poult. Sci. 2021, 100, 101405. [Google Scholar] [CrossRef] [PubMed]
- Svihus, B. Starch Digestion Capacity of Poultry. Poult. Sci. 2014, 93, 2394–2399. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Acosta, E. Almidón Resistente En La Nutrición de Animales Monogástricos I: Concepto, Clasificación y Fuentes. Nutr. Anim. Trop. 2018, 12, 59–79. [Google Scholar] [CrossRef] [Green Version]
- Kimball, S.R.; Jefferson, L.S. Regulation of Protein Synthesis by Branched-Chain Amino Acids. Curr. Opin. Clin. Nutr. Metab. Care 2001, 4, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Fouad, A.M.; El-Senousey, H.K.; Ruan, D.; Wang, S.; Xia, W.; Zheng, C. Tryptophan in Poultry Nutrition: Impacts and Mechanisms of Action. J. Anim. Physiol. Anim. Nutr. 2021, 105, 1146–1153. [Google Scholar] [CrossRef]
- Wen, J.; Helmbrecht, A.; Elliot, M.A.; Thomson, J.; Persia, M.E. Evaluation of the Tryptophan Requirement of Small-Framed First Cycle Laying Hens. Poult. Sci. 2019, 98, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Zheng, H.; Yang, L.; Li, H.; Tian, Y.; Wang, Y.; Lyu, S.; Brockmann, G.A.; Kang, X.; Liu, X. Dynamic Expression Profile, Regulatory Mechanism and Correlation with Egg-Laying Performance of ACSF Gene Family in Chicken (Gallus gallus). Sci. Rep. 2018, 8, 8457. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Age (Weeks) | Egg Production (Eggs/100 hens) | Feed Intake (g/day) | Feed Conversion Index Ratio | Mortality (%) |
|---|---|---|---|---|---|
| G1 | 21 | 76.23 ± 0.0223 a,B,A | 96.80 ± 0.8347 a,C | 2.67 ± 0.0621 a,A | 0 ± 0 a,A |
| G2 | 72.91 ± 0.0220 a,B,A | 94.44 ± 0.0 a,C | 2.82 ± 0.1033 a,B,A | 0 ± 0 a,A | |
| G3 | 72.56 ± 0.0107 a,B | 85.58 ± 1.5274 a,B | 2.39 ± 0.0482 a,B,A | 0 ± 0 a,A | |
| G4 | 71.53 ± 0.0175 a,B | 91.48 ± 1.0434 a,C | 2.82 ± 0.0687 a,B | 0 ± 0 a,A | |
| G1 | 22–31 | 83.49 ± 0.002 a,B | 107.80 ± 0.218 a,A | 2.48 ± 0.009 b,A | 0 ± 0 a,A |
| G2 | 81.51 ± 0.002 a,B | 105.77 ± 0.197 a,A | 2.59 ± 0.014 b,A | 0 ± 0 a,A | |
| G3 | 65.89 ± 0.002 b,B | 95.96 ± 0.176 b,A | 3.02 ± 0.019 b,a,A | 0.03 ± 0.002 a,A | |
| G4 | 63.93 ± 0.003 b,B | 98.48 ± 0.179 b,A | 3.661 ± 0.043 a,A | 0.03 ± 0.002 a,A | |
| G1 | 32–41 | 78.70 ± 0.001 b,A | 103.25 ± 0.096 a,B | 2.26 ± 0.005 a,B | 0 ± 0 a,A |
| G2 | 79.32 ± 0.002 b,A | 98.165 ± 0.174 b,B | 2.28 ± 0.009 a,B | 0 ± 0 a,A | |
| G3 | 89.84 ± 0.001 a,A | 103.25 ± 0.128 a,A | 1.94 ± 0.003 b,B | 0 ± 0 a,A | |
| G4 | 82.82 ± 0.002 b,A | 106.79 ± 0.199 a,B | 2.232 ± 0.006 a,B | 0 ± 0 a,A |
| Age (w) | Egg Weight (g) | Albumin Height (mm) | Yolk Color (−) | Albumin Quality (Haugh units) | Egg Shell Strength (Kgf) | Egg Shell Thickness (mm) | |
|---|---|---|---|---|---|---|---|
| G1 | 21 | 51.55 ± 0.271 a,B | 7.83 ± 0.066 a,A | 8.18 ± 0.048 a,B | 90.22 ± 0.295 a,A | 6.21 ± 0.052 a,A | 0.405 ± 0.002 a,A |
| G2 | 49.17 ± 0.183 a,B | 6.04 ± 0.085 b,B | 7.83 ± 0.069 a,B | 80.55 ± 0.538 b,A | 5.87 ± 0.090 a,A | 0.396 ± 0.002 a,A | |
| G3 | 49.34 ± 0.206 a,C | 5.85 ± 0.080 b,B | 7.41 ± 0.097 a,B | 79.14 ± 0.616 b,B | 5.62 ± 0.075 a,A | 0.398 ± 0.002 a,A | |
| G4 | 49.98 ± 0.459 a,B | 6.39 ± 0.115 b,B | 8.16 ± 0.069 a,B | 82.14 ± 0.809 b,B | 5.47 ± 0.081 a,A | 0.398 ± 0.002 a,A | |
| G1 | 31 | 61.74 ± 0.343 b,a,A | 8.72 ± 0.108 a,A | 11.66 ± 0.041 a,A | 92.50 ± 0.591 a,A | 4.83 ± 0.055 a,B | 0.350 ± 0.002 a,B |
| G2 | 59.18 ± 0.443 b,A | 8.27 ± 0.141 a,A | 8.83 ± 0.092 b,A | 90.40 ± 0.818 a,A | 4.54 ± 0.129 a,B,A | 0.344 ± 0.002 a,B | |
| G3 | 63.77 ± 0.221 a,A | 8.91 ± 0.126 a,A | 8.66 ± 0.108 b,A | 92.67 ± 0.803 a,A | 4.73 ± 0.104 a,B,A | 0.361 ± 0.003 a,B | |
| G4 | 61.94 ± 0.363 b,a,A | 9.02 ± 0.153 a,A | 11.66 ± 0.054 a,A | 93.04 ± 0.995 a,A | 4.87 ± 0.057 a,B,A | 0.358 ± 0.002 a,B | |
| G1 | 41 | 61.95 ± 0.437 a,A | 8.04 ± 0.211 a,A | 11.63 ± 0.158 a,A | 87.76 ± 1.218 a,A | 4.29 ± 0.108 a,B | 0.350 ± 0.002 a,B |
| G2 | 58.96 ± 0.582 a,A | 6.76 ± 0.225 a,B,A | 7.45 ± 0.094 b,B | 80.73 ± 1.579 a,A | 4.23 ± 0.116 a,B | 0.356 ± 0.003 a,B | |
| G3 | 58.48 ± 0.308 a,B | 7.29 ± 0.123 a,B,A | 7.45 ± 0.137 b,B | 83.60 ± 1.416 a,B,A | 4.35 ± 0.085 a,B | 0.339 ± 0.002 a,B | |
| G4 | 60.28 ± 0.426 a,A | 8.11 ± 0.175 a,A | 11.90 ± 0.094 a,A | 89.69 ± 0.622 a,B,A | 4.26 ± 0.114 a,B | 0.355 ± 0.004 a,B |
| Treatment | Age (w) | Raw Reads | Filtered Reads | Chao1 | Shannon | Simpson |
|---|---|---|---|---|---|---|
| G1 | 21 | 348,114 | 78,428 | 11 | 1.24 | 0.6 |
| G1 | 31 | 130,675 | 36,487 | 67 | 2.84 | 0.85 |
| G2 | 31 | 415,451 | 106,401 | 106 | 3.25 | 0.87 |
| G3 | 31 | 337,005 | 84,476 | 98 | 2.55 | 0.74 |
| G4 | 31 | 202,520 | 60,949 | 92 | 2.91 | 0.85 |
| G1 | 41 | 405,511 | 106,061 | 80 | 3.32 | 0.92 |
| G2 | 41 | 224,088 | 53,304 | 61 | 2.07 | 0.67 |
| G3 | 41 | 556,605 | 147,318 | 67 | 1.52 | 0.52 |
| G4 | 41 | 299,560 | 74,802 | 74 | 3.27 | 0.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruesga-Gutiérrez, E.; Ruvalcaba-Gómez, J.M.; Gómez-Godínez, L.J.; Villagrán, Z.; Gómez-Rodríguez, V.M.; Heredia-Nava, D.; Ramírez-Vega, H.; Arteaga-Garibay, R.I. Allium-Based Phytobiotic for Laying Hens’ Supplementation: Effects on Productivity, Egg Quality, and Fecal Microbiota. Microorganisms 2022, 10, 117. https://doi.org/10.3390/microorganisms10010117
Ruesga-Gutiérrez E, Ruvalcaba-Gómez JM, Gómez-Godínez LJ, Villagrán Z, Gómez-Rodríguez VM, Heredia-Nava D, Ramírez-Vega H, Arteaga-Garibay RI. Allium-Based Phytobiotic for Laying Hens’ Supplementation: Effects on Productivity, Egg Quality, and Fecal Microbiota. Microorganisms. 2022; 10(1):117. https://doi.org/10.3390/microorganisms10010117
Chicago/Turabian StyleRuesga-Gutiérrez, Edmundo, José Martín Ruvalcaba-Gómez, Lorena Jacqueline Gómez-Godínez, Zuamí Villagrán, Victor M. Gómez-Rodríguez, Darwin Heredia-Nava, Humberto Ramírez-Vega, and Ramón Ignacio Arteaga-Garibay. 2022. "Allium-Based Phytobiotic for Laying Hens’ Supplementation: Effects on Productivity, Egg Quality, and Fecal Microbiota" Microorganisms 10, no. 1: 117. https://doi.org/10.3390/microorganisms10010117
APA StyleRuesga-Gutiérrez, E., Ruvalcaba-Gómez, J. M., Gómez-Godínez, L. J., Villagrán, Z., Gómez-Rodríguez, V. M., Heredia-Nava, D., Ramírez-Vega, H., & Arteaga-Garibay, R. I. (2022). Allium-Based Phytobiotic for Laying Hens’ Supplementation: Effects on Productivity, Egg Quality, and Fecal Microbiota. Microorganisms, 10(1), 117. https://doi.org/10.3390/microorganisms10010117