
Comparative Genomics of Typical and Atypical Aeromonas salmonicida Complete Genomes Revealed New Insights into Pathogenesis Evolution

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Material and Methods
2.1. Bacterial Characterization
2.1.1. Aeromonas salmonicida Strains
2.1.2. Bacterial Culture Conditions
2.1.3. Phenotypic, Biochemical, and Enzymatic Testing
2.1.4. Hemolysin Assay
2.1.5. Siderophores Secretion Assay
2.1.6. SDS-PAGE and Western Blot
2.2. Genomic Analyses
2.2.1. DNA Extraction, Sequencing, Genome Assembly, Annotation, and Mapping
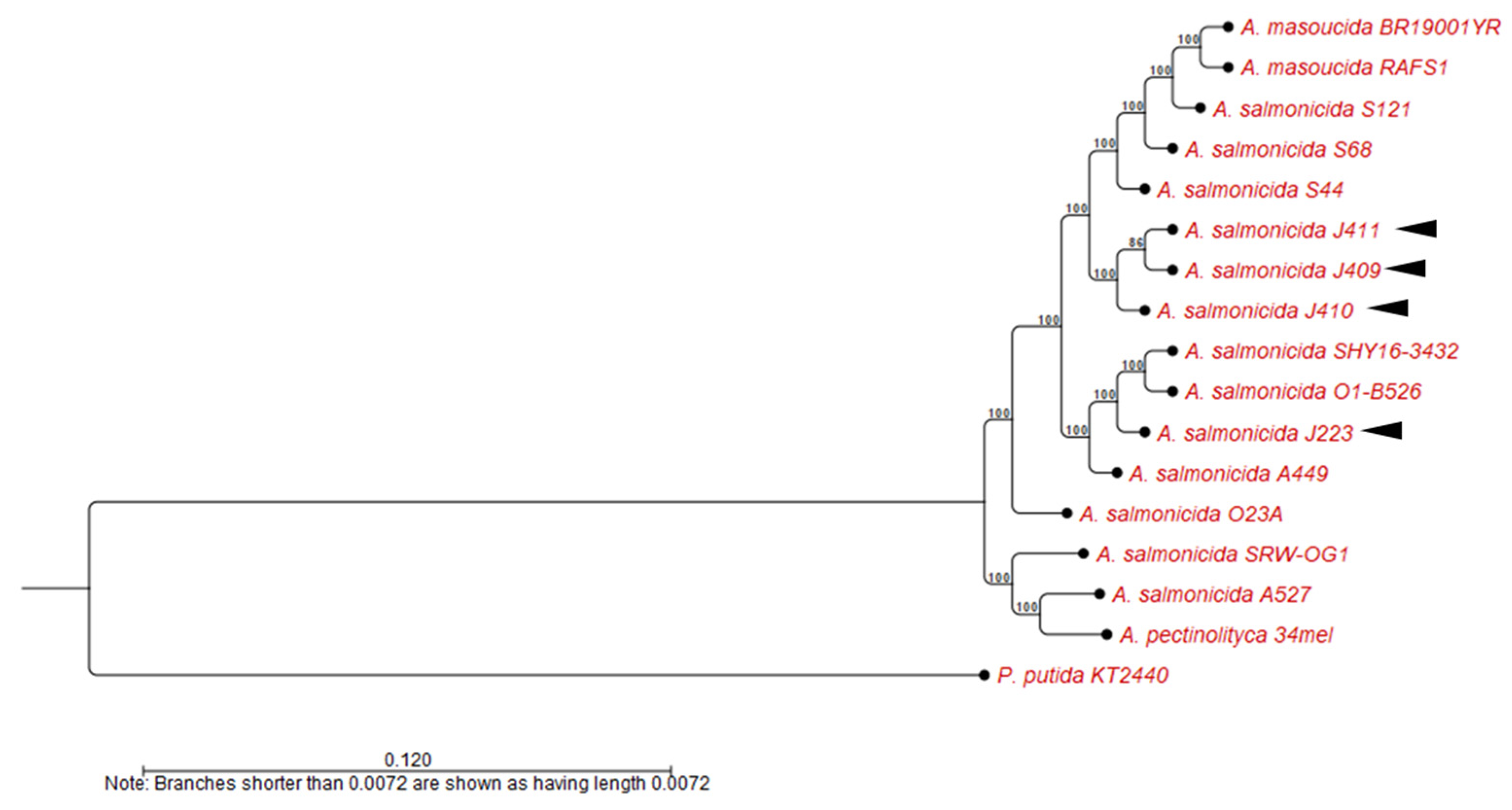
2.2.2. Whole-Genome Alignment and Phylogenetic Analysis
2.2.3. Non-Coding RNAs Prediction
2.2.4. Virulence Factors, Insertion Sequences (ISs), and Antibiotic Resistant Gene Analysis
2.3. Statistical Analysis
3. Results
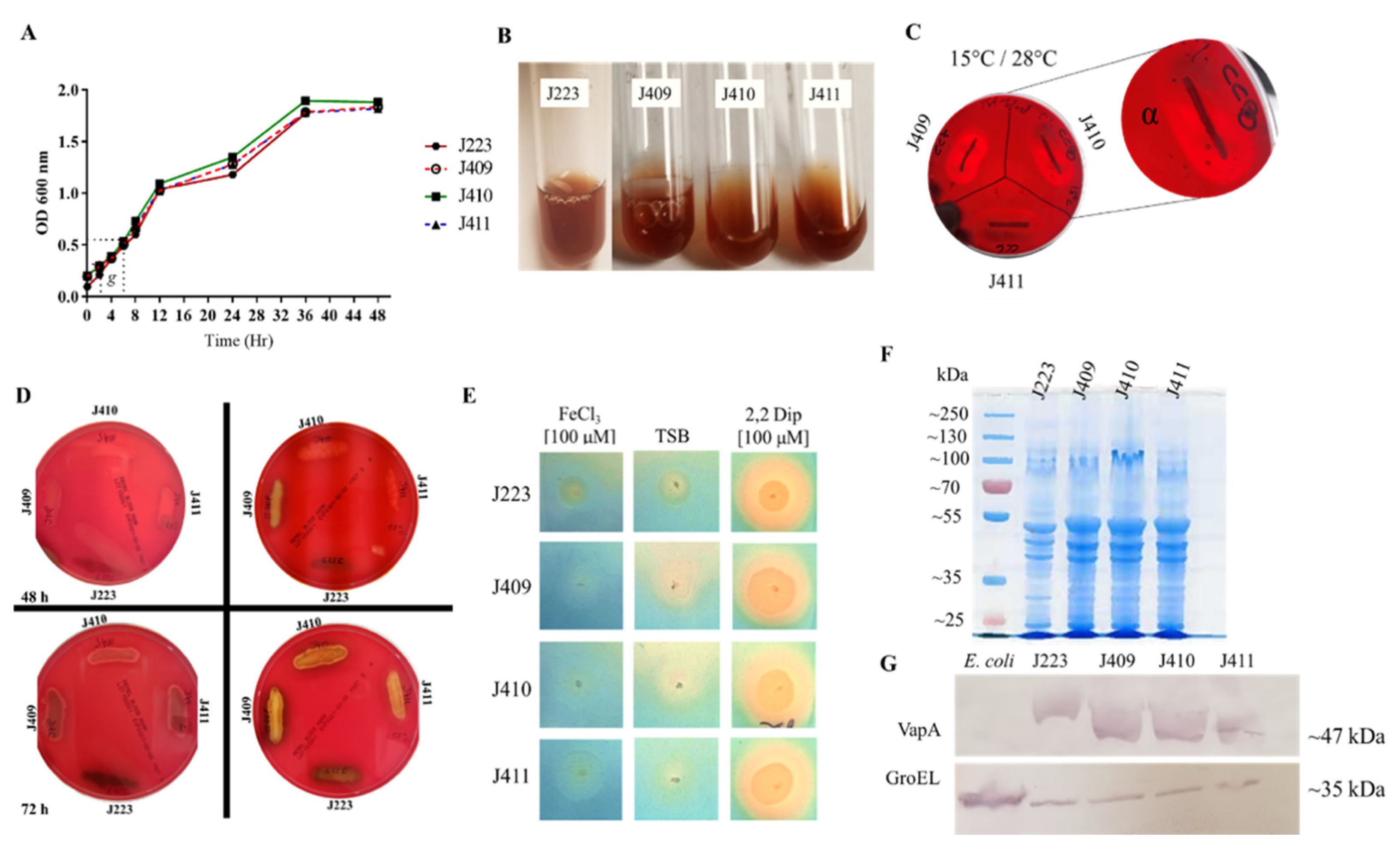
3.1. A. salmonicida Phenotypic Characteristics, Biochemical, and Enzymatic Profiles
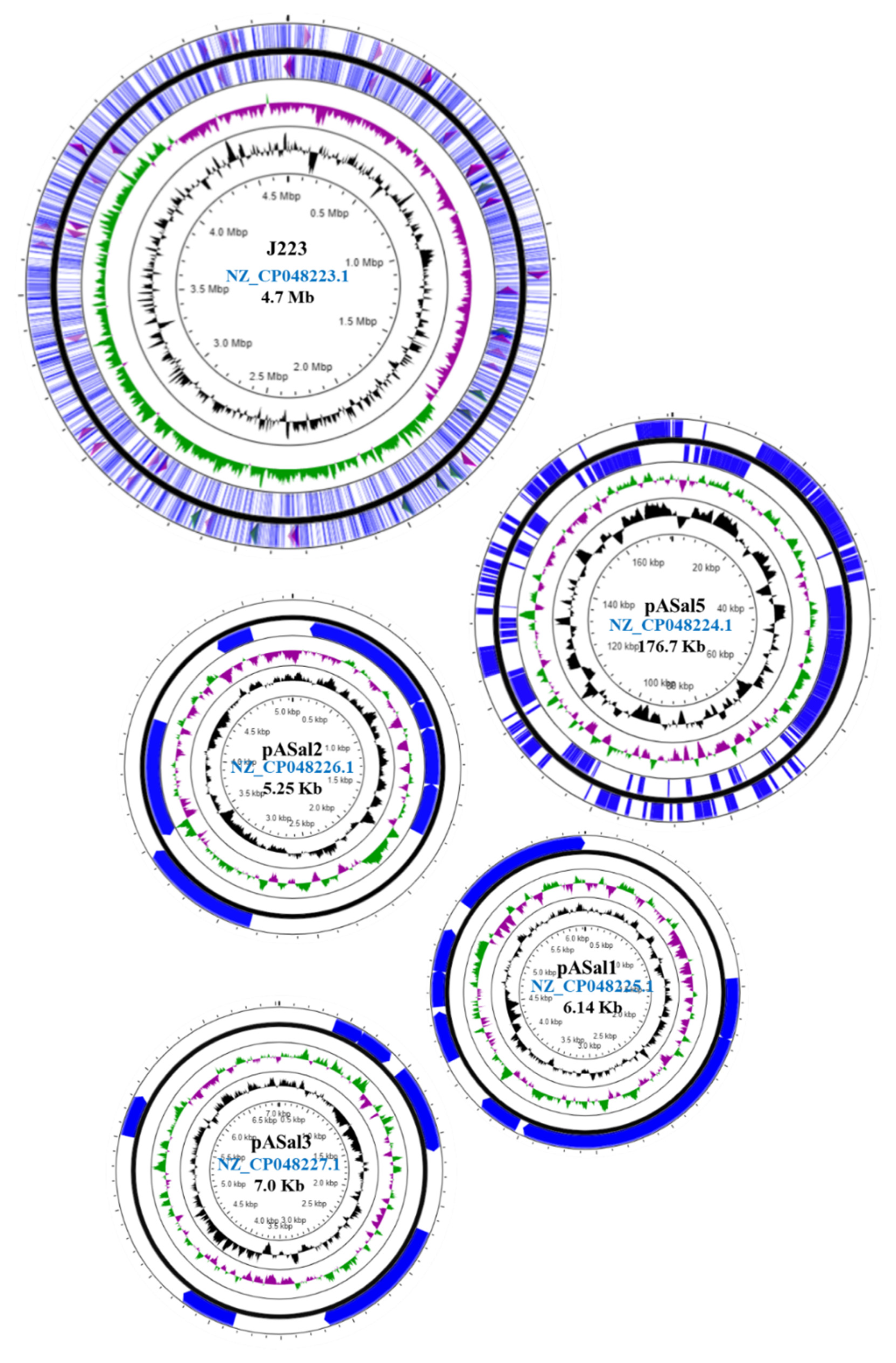
3.2. Genome Sequencing and Annotation
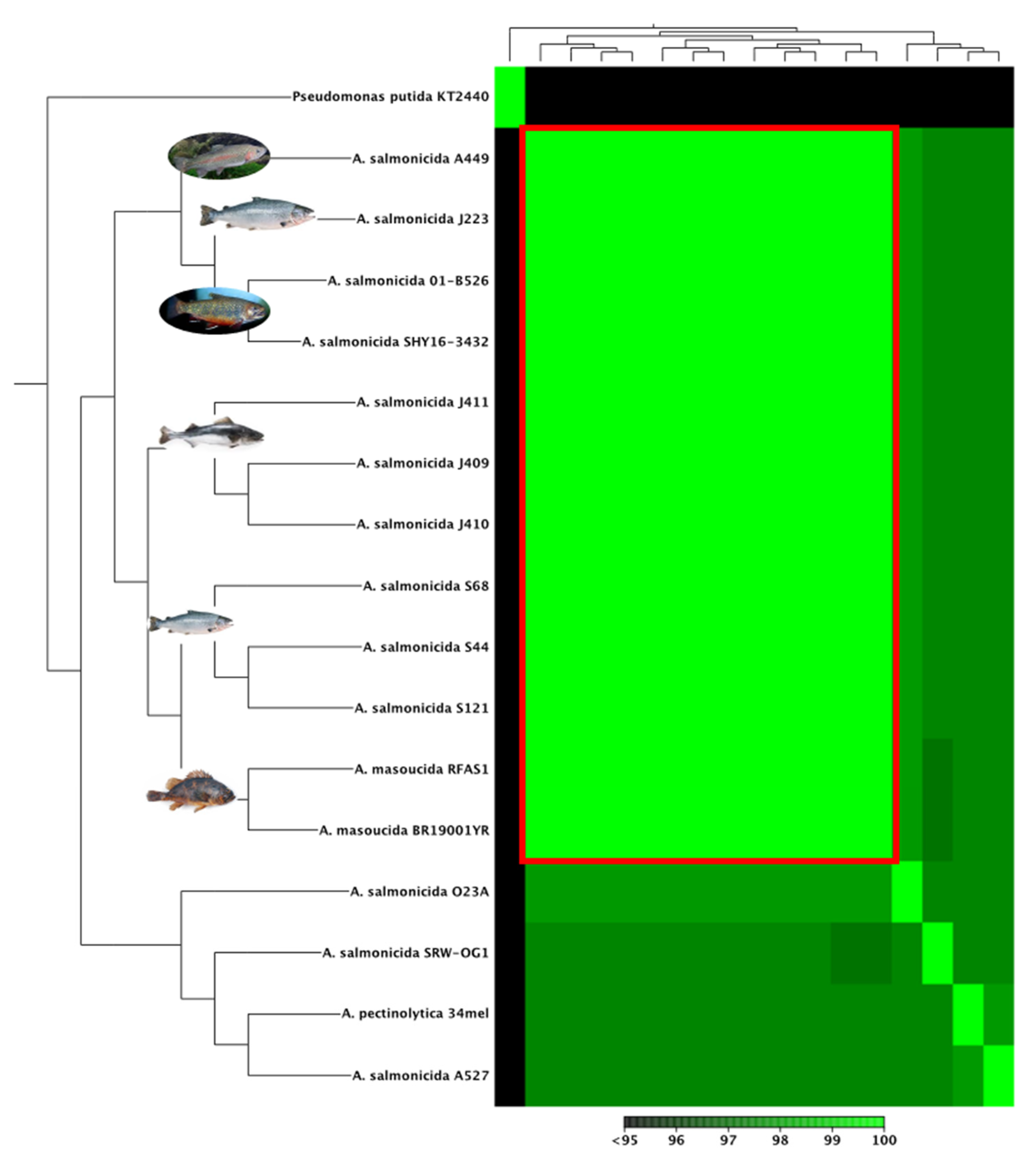
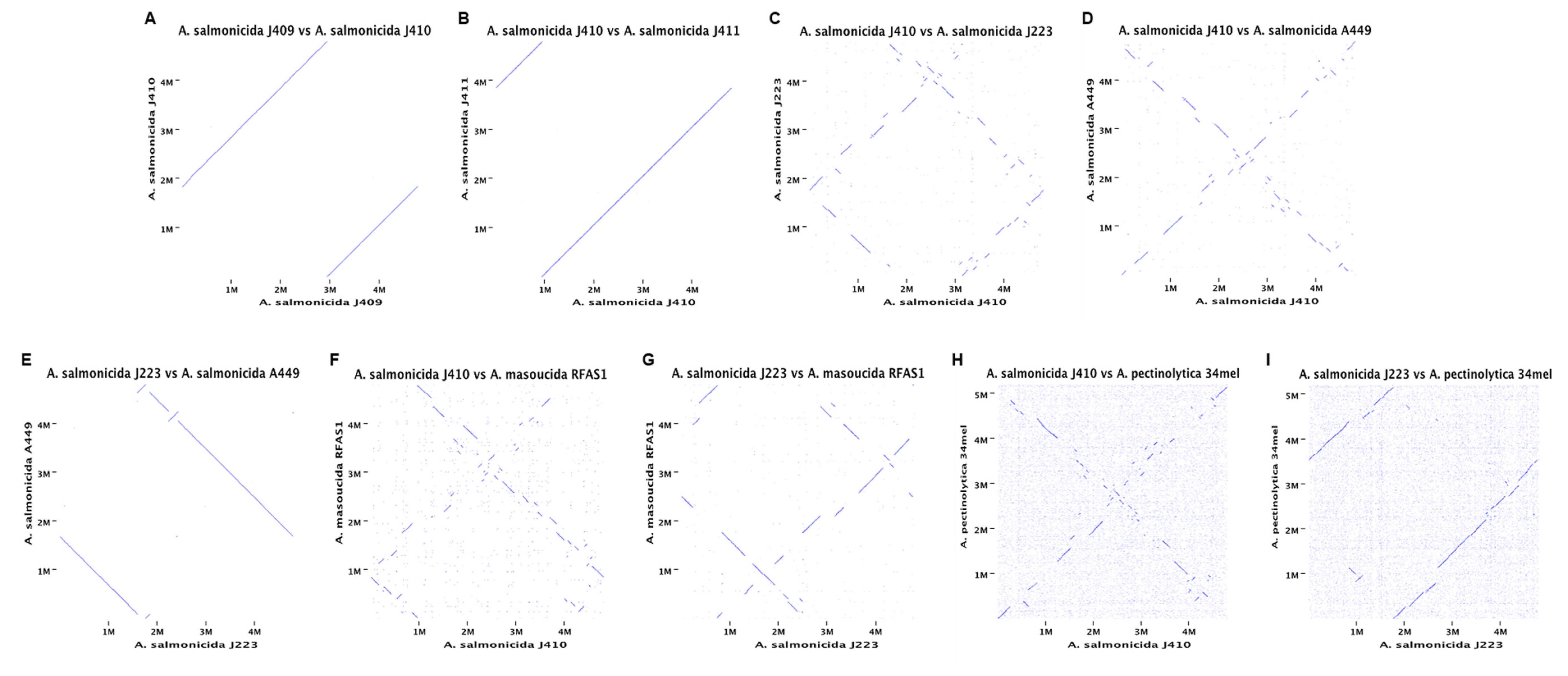
3.3. Comparative Analyses of A. salmonicida Chromosome
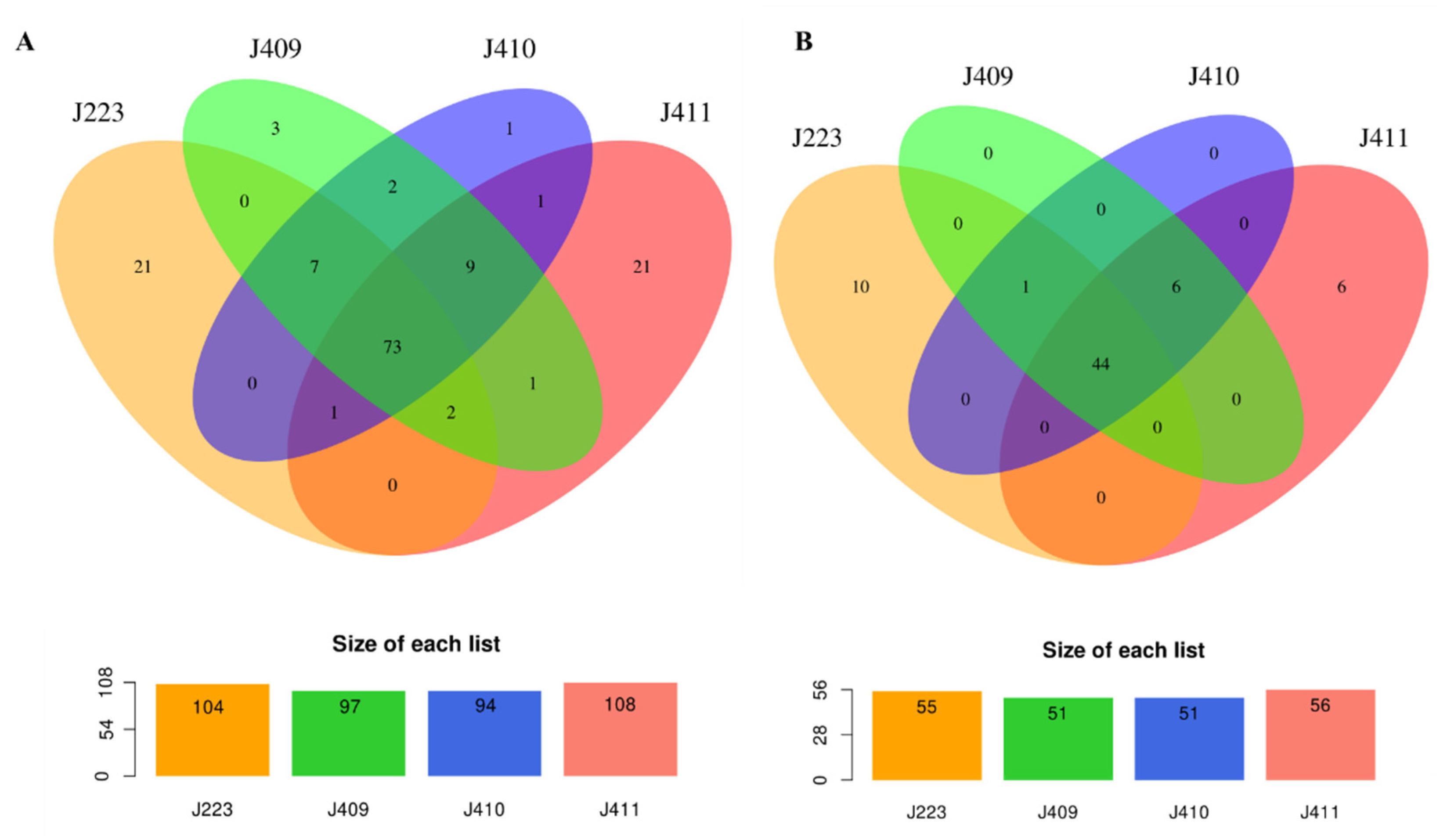
3.4. Pathogenesis and Environmental Adaptation-Associated Genes in A. salmonicida Genome
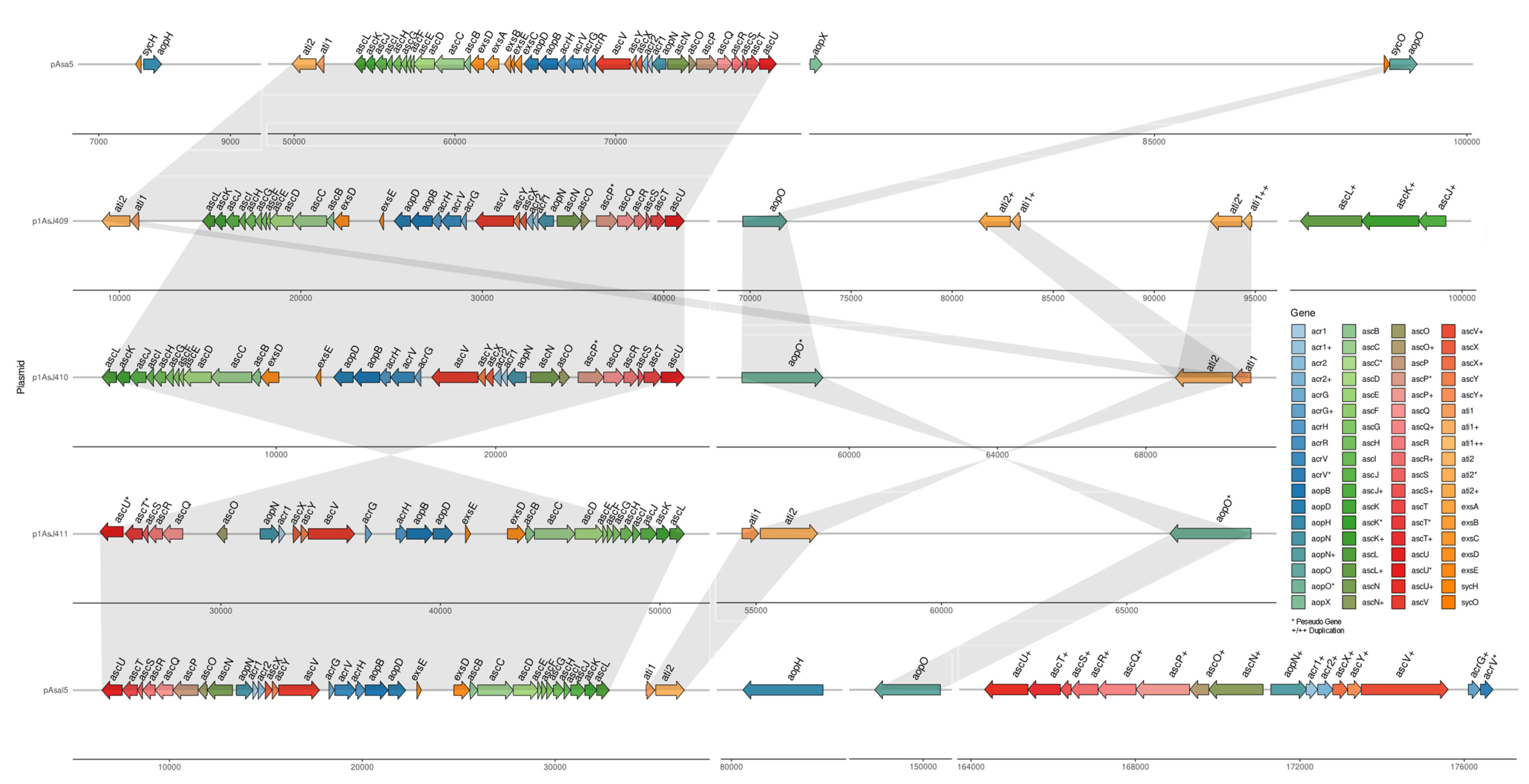
3.5. Plasmidome of A. salmonicida Sequenced Strains
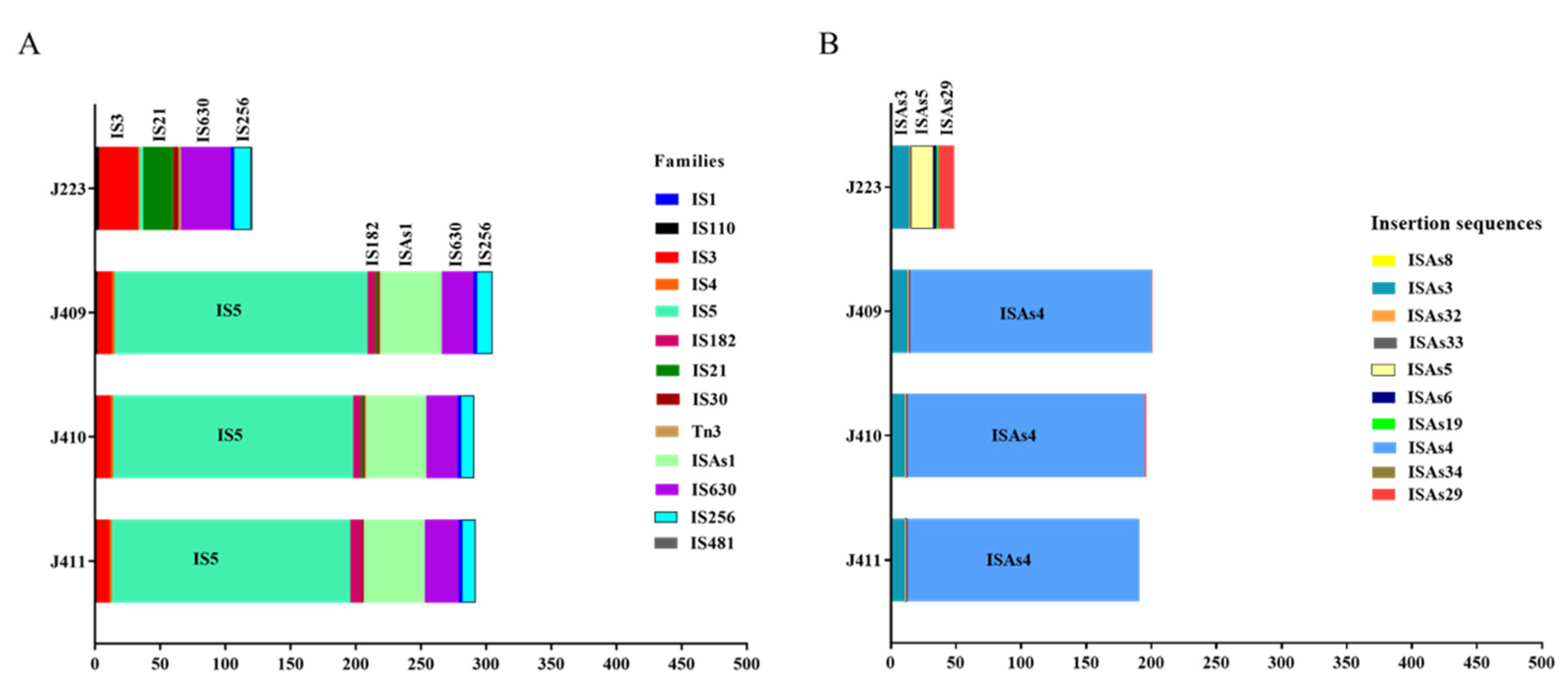
3.6. A. salmonicida Insertion Sequences (ISs)
3.7. Typical and Atypical A. salmonicida Non-Coding RNAs Repertory
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| °C | Degree centigrade |
| µL | Microliter |
| CFU | Colony forming unit |
| h | Hour |
| Kb | Kilo base pair |
| L | Liter |
| Mb | Mega base pair |
| mg | Milligram |
| min | Minute |
| mL | Milliliter |
| OD | Optical density |
| PBS | Phosphate buffered saline |
| TSB | Tryptic Soy Broth media |
| TSA | Tryptic Soy Agar |
| SEM | Standard error of the mean |
| Subsp. | Subspecies |
| ISs | Insertion sequences |
| GGs | genomes gaps |
References
- Cipriano, R.C.; Bullock, G.L. Furunculosis and Other Diseases Caused by Aeromonas salmonicida; Fish Disease Leaflet; US Fish & Wildlife Publications: Falls Church, VA, USA, 2001. [Google Scholar]
- Dallaire-Dufresne, S.; Tanaka, K.H.; Trudel, M.V.; Lafaille, A.; Charette, S.J. Virulence, genomic features, and plasticity of Aeromonas salmonicida subsp. salmonicida, the causative agent of fish furunculosis. Vet. Microbiol. 2014, 169, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wiklund, T.; Dalsgaard, I. Occurrence and significance of atypical Aeromonas salmonicida in non-salmonid and salmonid fish species: A review. Dis. Aquat. Org. 1998, 32, 49–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colston, S.M.; Fullmer, M.S.; Beka, L.; Lamy, B.; Gogarten, J.P.; Graf, J. Bioinformatic Genome Comparisons for Taxonomic and Phylogenetic Assignments Using Aeromonas as a Test Case. mBio 2014, 5, e02136-14. [Google Scholar] [CrossRef] [Green Version]
- Vincent, A.T.; Trudel, M.V.; Freschi, L.; Nagar, V.; Gagné-Thivierge, C.; Levesque, R.C.; Charette, S.J. Increasing genomic diversity and evidence of constrained lifestyle evolution due to insertion sequences in Aeromonas salmonicida. BMC Genom. 2016, 17, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Qiao, Y.; Chai, B.; Qiu, C.; Chen, X. Identification and Molecular Characterization of the Homogentisate Pathway Responsible for Pyomelanin Production, the Major Melanin Constituents in Aeromonas media WS. PLoS ONE 2015, 10, e0120923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, B.; Austin, D.; Dalsgaard, I.; Gudmundsdóttir, B.; Høie, S.; Thornton, J.; Larsen, J.; O’Hici, B.; Powell, R. Characterization of Atypical Aeromonas salmonicida Different Methods. Syst. Appl. Microbiol. 1998, 21, 50–64. [Google Scholar] [CrossRef]
- Emond-Rheault, J.-G.; Vincent, A.T.; Trudel, M.V.; Brochu, F.; Boyle, B.; Tanaka, K.H.; Attéré, S.A.; Jubinville, É.; Loch, T.P.; Winters, A.D.; et al. Variants of a genomic island in Aeromonas salmonicida subsp. salmonicida link isolates with their geographical origins. Vet. Microbiol. 2015, 175, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Chu, S.; Noonan, B.; Cavaignac, S.; Trust, T.J. Endogenous mutagenesis by an insertion sequence element identifies Aeromonas salmonicida AbcA as an ATP-binding cassette transport protein required for biogenesis of smooth lipopolysaccharide. Proc. Natl. Acad. Sci. USA 1995, 92, 5754–5758. [Google Scholar] [CrossRef] [Green Version]
- Lund, V.; Mikkelsen, H. Genetic diversity among A-proteins of atypical strains of Aeromonas salmonicida. Dis. Aquat. Org. 2004, 61, 257–262. [Google Scholar] [CrossRef]
- Gulla, S.; Lund, V.; Kristoffersen, A.B.; Sørum, H.; Colquhoun, D.J. vapA (A-layer) typing differentiates Aeromonas salmonicida subspecies and identifies a number of previously undescribed subtypes. J. Fish Dis. 2016, 39, 329–342. [Google Scholar] [CrossRef]
- Reith, M.E.; Singh, R.K.; Curtis, B.A.; Boyd, J.M.; Bouevitch, A.; Kimball, J.; Munholland, J.; Murphy, C.; Sarty, D.; Williams, J.; et al. The genome of Aeromonas salmonicida subsp. salmonicida A449: Insights into the evolution of a fish pathogen. BMC Genom. 2008, 9, 1–15. [Google Scholar] [CrossRef]
- Trudel, M.V.; Tanaka, K.H.; Filion, G.; Daher, R.K.; Frenette, M.; Charette, S.J. Insertion sequenceAS5(ISAS5) is involved in the genomic plasticity of Aeromonas salmonicida. Mob. Genet. Elem. 2013, 3, e25640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, A.T.; Le Breton, A.; Bernatchez, A.; Gagné-Thivierge, C.; Paquet, V.E.; Thibault, E.; Charette, S.J.; Gantelet, H. Draft Genome Sequences of Four Aeromonas salmonicida subsp. achromogenes Strains, 23051, 23053, 23055, and 23056, Isolated from Senegalese Sole (Solea senegalensis). Microbiol. Resour. Announc. 2019, 8, e00631-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.H.; Frenette, M.; Charette, S.J. IS-mediated loss of virulence by Aeromonas salmonicida. Mob. Genet. Elem. 2013, 3, e23498. [Google Scholar] [CrossRef]
- Gustafson, C.E.; Chu, S.; Trust, T.J. Mutagenesis of the Paracrystalline Surface Protein Array of Aeromonas salmonicida by Endogenous Insertion Elements. J. Mol. Biol. 1994, 237, 452–463. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.T.; Hosseini, N.; Charette, S.J. The Aeromonas salmonicida plasmidome: A model of modular evolution and genetic diversity. Ann. N. Y. Acad. Sci. 2021, 1488, 16–32. [Google Scholar] [CrossRef]
- Rivas, E.; Klein, R.J.; Jones, T.A.; Eddy, S.R. Computational identification of noncoding RNAs in E. coli by comparative genomics. Curr. Biol. 2001, 11, 1369–1373. [Google Scholar] [CrossRef] [Green Version]
- Wassarman, K.M.; Repoila, F.; Rosenow, C.; Storz, G.; Gottesman, S. Identification of novel small RNAs using comparative genomics and microarrays. Genes Dev. 2001, 15, 1637–1651. [Google Scholar] [CrossRef] [Green Version]
- Argaman, L.; Hershberg, R.; Vogel, J.; Bejerano, G.; Wagner, E.H.; Margalit, H.; Altuvia, S. Novel small RNA-encoding genes in the intergenic regions of Escherichia coli. Curr. Biol. 2001, 11, 941–950. [Google Scholar] [CrossRef] [Green Version]
- Wagner, E.G.H.; Romby, P. Small RNAs in Bacteria and Archaea: Who they are, what they do, and how they do it. Adv. Genet. 2015, 90, 133–208. [Google Scholar] [CrossRef]
- Babski, J.; Maier, L.-K.; Heyer, R.; Jaschinski, K.; Prasse, D.; Jäger, D.; Randau, L.; A Schmitz, R.; Marchfelder, A.; Soppa, J. Small regulatory RNAs in Archaea. RNA Biol. 2014, 11, 484–493. [Google Scholar] [CrossRef] [Green Version]
- Segovia, C.; Arias-Carrasco, R.; Yañez, A.J.; Maracaja-Coutinho, V.; Santander, J. Core non-coding RNAs of Piscirickettsia salmonis. PLoS ONE 2018, 13, e0197206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyrud, M.L.J. A Study of Regulatory Small RNAs in Vibrio Salmonicida: Construction of a Knock-Out Mutant and a cDNA Library. Master’s Thesis, Universitetet i Tromsø, Tromsø, Norway, 2008. [Google Scholar]
- Ahmad, R.; Hansen, G.Å.; Hansen, H.; Hjerde, E.; Pedersen, H.L.; Paulsen, S.M.; Nyrud, M.L.J.; Strauss, A.; Willassen, N.-P.; Haugen, P. Prediction, Microarray and Northern Blot Analyses Identify New Intergenic Small RNAs in Aliivibrio salmonicida. J. Mol. Microbiol. Biotechnol. 2012, 22, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Valderrama, K.; Saravia, M.; Santander, J. Phenotype of Aeromonas salmonicida sp. Salmonicida cyclic adenosine 3′,5′-monophosphate receptor protein (Crp) mutants and its virulence in rainbow trout (Oncorhynchus mykiss). J. Fish Dis. 2017, 40, 1849–1856. [Google Scholar] [CrossRef] [PubMed]
- Cipriano, R.C.; Bertolini, J. Selection for virulence in the fish pathogen Aeromonas salmonicida, using Coomassie Brilliant Blue aga. J. Wildl. Dis. 1988, 24, 672–678. [Google Scholar] [CrossRef]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of Blue Agar CAS Assay for Siderophore Detection. J. Microbiol. Biol. Educ. 2011, 12, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Wood, E.J. Molecular Cloning. A Laboratory Manual. Biochem. Educ. 1983, 11, 82. [Google Scholar] [CrossRef]
- Ryan, K.J.; Schoenknecht, F.D.; Kirby, W.M.M. Disc Sensitivity Testing. Hosp. Pract. 1970, 5, 91–100. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, W. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Press: New York, NY, USA, 2001. [Google Scholar]
- Dower, W.J.; Miller, J.F.; Ragsdale, C.W. High efficiency transformation of E.coli by high voltage electroporation. Nucleic Acids Res. 1988, 16, 6127–6145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SDS sample buffer (2X). Cold Spring Harb. Protoc. 2009. Available online: https://doi.org/10.1101/pdb.rec11975 (accessed on 29 November 2021).
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 29 November 2021).
- Chin, C.-S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E.; et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 2013, 10, 563–569. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.Y.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasquez, I.; Cao, T.; Chakraborty, S.; Gnanagobal, H.; O’Brien, N.; Monk, J.; Boyce, D.; Westcott, J.D.; Santander, J. Comparative Genomics Analysis of Vibrio anguillarum Isolated from Lumpfish (Cyclopterus lumpus) in Newfoundland Reveal Novel Chromosomal Organizations. Microorganisms 2020, 8, 1666. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Jukes, T.H.; Cantor, C.R. Chapter 24—Evolution of Protein Molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: Cambridge, MA, USA, 1969; pp. 21–132. [Google Scholar]
- Teru, Y.; Hikima, J.-I.; Kono, T.; Sakai, M.; Takano, T.; Hawke, J.P.; Takeyama, H.; Aoki, T. Whole-Genome Sequence of Photobacterium damselae subsp. piscicida Strain 91-197, Isolated from Hybrid Striped Bass (Morone sp.) in the United States. Genome Announc. 2017, 5, e00600-17. [Google Scholar] [CrossRef] [Green Version]
- Vasquez, I.; Cao, T.; Hossain, A.; Valderrama, K.; Gnanagobal, H.; Dang, M.; Leeuwis, R.; Ness, M.; Campbell, B.; Gendron, R.; et al. Aeromonas salmonicida infection kinetics and protective immune response to vaccination in sablefish (Anoplopoma fimbria). Fish Shellfish. Immunol. 2020, 104, 557–566. [Google Scholar] [CrossRef]
- Boyd, J.; Williams, J.; Curtis, B.; Kozera, C.; Singh, R.; Reith, M. Three small, cryptic plasmids from Aeromonas salmonicida subsp. salmonicida A449. Plasmid 2003, 50, 131–144. [Google Scholar] [CrossRef]
- Michel, C. Furunculosis of salmonids: Vaccination attempts in rainbow trout (Salmo gairdneri) by formalin-killed germs. Ann. Rech. Vet. Ann. Vet. Res. 1979, 10, 33–40. [Google Scholar]
- Charette, S.J.; Brochu, F.; Boyle, B.; Filion, G.; Tanaka, K.H.; Derome, N. Draft Genome Sequence of the Virulent Strain 01-B526 of the Fish Pathogen Aeromonas salmonicida. J. Bacteriol. 2012, 194, 722–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.H.; Vincent, A.T.; Emond-Rheault, J.-G.; Adamczuk, M.; Frenette, M.; Charette, S.J. Plasmid composition in Aeromonas salmonicida subsp. salmonicida 01-B526 unravels unsuspected type three secretion system loss patterns. BMC Genom. 2017, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Massicotte, M.-A.; Vincent, A.T.; Schneider, A.; Paquet, V.E.; Frenette, M.; Charette, S.J. One Aeromonas salmonicida subsp. salmonicida isolate with a pAsa5 variant bearing antibiotic resistance and a pRAS3 variant making a link with a swine pathogen. Sci. Total Environ. 2019, 690, 313–320. [Google Scholar] [CrossRef]
- Pfeiffer, F.; Zamora-Lagos, M.-A.; Blettinger, M.; Yeroslaviz, A.; Dahl, A.; Gruber, S.; Habermann, B.H. The complete and fully assembled genome sequence of Aeromonas salmonicida subsp. pectinolytica and its comparative analysis with other Aeromonas species: Investigation of the mobilome in environmental and pathogenic strains. BMC Genom. 2018, 19, 20. [Google Scholar] [CrossRef] [Green Version]
- Vincent, A.T.; Rouleau, F.D.; Moineau, S.; Charette, S.J. Study of mesophilic Aeromonas salmonicida A527 strain sheds light on the species’ lifestyles and taxonomic dilemma. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhrynowski, W.; Decewicz, P.; Dziewit, L.; Radlinska, M.; Krawczyk, P.S.; Lipinski, L.; Adamska, D.; Drewniak, L. Analysis of the Genome and Mobilome of a Dissimilatory Arsenate Reducing Aeromonas sp. O23A Reveals Multiple Mechanisms for Heavy Metal Resistance and Metabolism. Front. Microbiol. 2017, 8, 936. [Google Scholar] [CrossRef]
- Kim, A.; Nguyen, T.L.; Kim, D.-H. Complete Genome Sequence of the Virulent Aeromonas salmonicida subsp. masoucida Strain RFAS1. Genome Announc. 2018, 6, e00470-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, K.E.; Weinel, C.; Paulsen, I.; Dodson, R.J.; Hilbert, H.; Dos Santos, V.A.P.M.; Fouts, D.E.; Gill, S.R.; Pop, M.; Holmes, M.; et al. Complete genome sequence and comparative analysis of the metabolically versatile Pseudomonas putida KT2440. Environ. Microbiol. 2002, 4, 799–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arias-Carrasco, R.; Vásquez-Morán, Y.; Nakaya, H.I.; Maracaja-Coutinho, V. StructRNAfinder: An automated pipeline and web server for RNA families prediction. BMC Bioinform. 2018, 19, 55. [Google Scholar] [CrossRef] [Green Version]
- Nawrocki, E.P.; Eddy, S.R. Infernal 1.1: 100-fold faster RNA homology searches. Bioinformatics 2013, 29, 2933–2935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denman, R.B. Using RNAFOLD to predict the activity of small catalytic RNAs. Biotechniques 1993, 15, 1090–1095. [Google Scholar] [PubMed]
- Kalvari, I.; Nawrocki, E.P.; Ontiveros-Palacios, N.; Argasinska, J.; Lamkiewicz, K.; Marz, M.; Griffiths-Jones, S.; Toffano-Nioche, C.; Gautheret, D.; Weinberg, Z.; et al. Rfam 14: Expanded coverage of metagenomic, viral and microRNA families. Nucleic Acids Res. 2021, 49, D192–D200. [Google Scholar] [CrossRef]
- Quinlan, A.R. BEDTools: The Swiss-Army Tool for Genome Feature Analysis. Curr. Protoc. Bioinform. 2014, 47, 11–12. [Google Scholar] [CrossRef] [PubMed]
- Hadley, W. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Bardou, P.; Mariette, J.; Escudié, F.; Djemiel, C.; Klopp, C. jvenn: An interactive Venn diagram viewer. BMC Bioinform. 2014, 15, 293. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Yang, J.; Yu, J.; Yao, Z.; Sun, L.; Shen, Y.; Jin, Q. VFDB: A reference database for bacterial virulence factors. Nucleic Acids Res. 2005, 33, D325–D328. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [Green Version]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 2016, 45, D566–D573. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.P.; Lowenthal, A.C.; Harshey, R.M.; Ottemann, K.M. CheV: CheW-like coupling proteins at the core of the chemotaxis signaling network. Trends Microbiol. 2010, 18, 494–503. [Google Scholar] [CrossRef] [Green Version]
- Canals, R.; Ramirez, S.; Vilches, S.; Horsburgh, G.; Shaw, J.G.; Tomás, J.M.; Merino, S. Polar Flagellum Biogenesis in Aeromonas hydrophila. J. Bacteriol. 2006, 188, 464–468. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.; Ly, P.M.; Feldman, M.F. The Tip of the VgrG Spike Is Essential to Functional Type VI Secretion System Assembly in Acinetobacter baumannii. mBio 2020, 11, e02761-19. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Garcia, F.; Ruiz-Perez, F.; Cataldi, Á.; Larzabal, M. Type VI Secretion System in Pathogenic Escherichia coli: Structure, Role in Virulence, and Acquisition. Front. Microbiol. 2019, 10, 1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Origgi, F.; Benedicenti, O.; Segner, H.; Sattler, U.; Wahli, T.; Frey, J. Aeromonas salmonicida type III secretion system-effectors-mediated immune suppression in rainbow trout (Oncorhynchus mykiss). Fish Shellfish. Immunol. 2017, 60, 334–345. [Google Scholar] [CrossRef]
- Lian, Z.; Bai, J.; Hu, X.; Lü, A.; Sun, J.; Guo, Y.; Song, Y. Detection and characterization of Aeromonas salmonicida subsp. salmonicida infection in crucian carp Carassius auratus. Vet. Res. Commun. 2020, 44, 61–72. [Google Scholar] [CrossRef]
- Casas, C.; Anderson, E.C.; Ojo, K.K.; Keith, I.; Whelan, D.; Rainnie, D.; Roberts, M.C. Characterization of pRAS1-like plasmids from atypical North American psychrophilic Aeromonas salmonicida. FEMS Microbiol. Lett. 2005, 242, 59–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, B.; Austin, D.A. Bacterial Fish Pathogens: Disease of Farmed and Wild Fish, 5th ed.; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Martin-Carnahan, A.; Joseph, S. Aeromonadales ord. nov. In Bergey’s Manual® of Systematic Bacteriology, 2nd ed.; Springer: New York, NY, USA, 2005. [Google Scholar]
- Merino, S.; De Mendoza, E.; Canals, R.; Tomás, J.M. Functional Genomics of the Aeromonas salmonicida Lipopolysaccharide O-Antigen and A-Layer from Typical and Atypical Strains. Mar. Drugs 2015, 13, 3791–3808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awan, F.; Dong, Y.; Liu, J.; Wang, N.; Mushtaq, M.H.; Lu, C.; Liu, Y. Comparative genome analysis provides deep insights into Aeromonas hydrophila taxonomy and virulence-related factors. BMC Genom. 2018, 19, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Rouleau, F.D.; Vincent, A.T.; Charette, S.J. Genomic and phenotypic characterization of an atypical Aeromonas salmonicida strain isolated from a lumpfish and producing unusual granular structures. J. Fish Dis. 2018, 41, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Ratledge, C.; Dover, L.G. Iron Metabolism in Pathogenic Bacteria. Annu. Rev. Microbiol. 2000, 54, 881–941. [Google Scholar] [CrossRef]
- Hirono, I.; Aoki, T. Cloning and characterization of three hemolysin genes from Aeromonas salmonicida. Microb. Pathog. 1993, 15, 269–282. [Google Scholar] [CrossRef]
- Liu, R.; Ochman, H. Origins of Flagellar Gene Operons and Secondary Flagellar Systems. J. Bacteriol. 2007, 189, 7098–7104. [Google Scholar] [CrossRef] [Green Version]
- Canals, R.; Altarriba, M.; Vilches, S.; Horsburgh, G.; Shaw, J.G.; Tomás, J.M.; Merino, S. Analysis of the Lateral Flagellar Gene System of Aeromonas hydrophila AH-3. J. Bacteriol. 2006, 188, 852–862. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.D.; Ozinsky, A. Toll-Like Receptor-5 and the Innate Immune Response to Bacterial Flagellin. Curr. Top. Microbiol. Immunol. 2002, 270, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.; Akira, S.; Underhill, D.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef]
- Soto-Dávila, M.; Valderrama, K.; Inkpen, S.M.; Hall, J.R.; Rise, M.L.; Santander, J. Effects of Vitamin D2 (Ergocalciferol) and D3 (Cholecalciferol) on Atlantic Salmon (Salmo salar) Primary Macrophage Immune Response to Aeromonas salmonicida subsp. salmonicida Infection. Front. Immunol. 2020, 10, 3011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frey, J.; Origgi, F.C. Type III Secretion System of Aeromonas salmonicida Undermining the Host’s Immune Response. Front. Mar. Sci. 2016, 3, 130. [Google Scholar] [CrossRef]
- Burr, S.E.; Stuber, K.; Wahli, T.; Frey, J. Evidence for a Type III Secretion System in Aeromonas salmonicida subsp. salmonicida. J. Bacteriol. 2002, 184, 5966–5970. [Google Scholar] [CrossRef] [Green Version]
- Dacanay, A.; Knickle, L.; Solanky, K.S.; Boyd, J.M.; Walter, J.A.; Brown, L.L.; Johnson, S.C.; Reith, M. Contribution of the type III secretion system (TTSS) to virulence of Aeromonas salmonicida subsp. salmonicida. Microbiology 2006, 152, 1847–1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebanks, R.O.; Knickle, L.C.; Goguen, M.; Boyd, J.M.; Pinto, D.M.; Reith, M.; Ross, N.W. Expression of and secretion through the Aeromonas salmonicida type III secretion system. Microbiology 2006, 152, 1275–1286. [Google Scholar] [CrossRef] [Green Version]
- Dang, M.; Dove, S.; Cao, T.; Hossein, A.; Gnanagobal, H.; Soto-Dávila, M.; Vasquez, I.; Chakraborty, S.; Rise, M.L.; Fleming, I.; et al. Atlantic Salmon (Salmo salar L.) wild type, farm, and hybrid crosses susceptibility to Aeromonas salmonicida subsp. salmonicida. (Oral presentation). In Proceedings of the Science Atlantic Aquaculture and Fisheries and Biology Conference, St. John’s, NL, Canada, 1 March 2021. [Google Scholar]
- Chakraborty, S.; Cao, T.; Hossain, A.; Gnanagobal, H.; Kumar, S.; Boyce, D.; Santander, J. Aeromonas salmonicida infection and vaccination in Cyclopterus lumpus (Oral Presentation). In Proceedings of the Conference of Research Workers in Animal Diseases, Chicago, IL, USA, 2–5 November 2019. [Google Scholar]
- Tanaka, K.H.; Dallaire-Dufresne, S.; Daher, R.K.; Frenette, M.; Charette, S.J. An Insertion Sequence-Dependent Plasmid Rearrangement in Aeromonas salmonicida Causes the Loss of the Type Three Secretion System. PLoS ONE 2012, 7, e33725. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, W.; Gudkovs, N.; Strom, M. Atypical strains of Aeromonas salmonicida contain multiple copies of insertion element ISAsa4, useful as a genetic marker and a target for PCR assay. Dis. Aquat. Org. 2006, 70, 209–217. [Google Scholar] [CrossRef]
- Eddy, S.R. Noncoding RNA genes. Curr. Opin. Genet. Dev. 1999, 9, 695–699. [Google Scholar] [CrossRef]
- Quereda, J.J.; Cossart, P. Regulating Bacterial Virulence with RNA. Annu. Rev. Microbiol. 2017, 71, 263–280. [Google Scholar] [CrossRef]
- Gottesman, S. Micros for microbes: Non-coding regulatory RNAs in bacteria. Trends Genet. 2005, 21, 399–404. [Google Scholar] [CrossRef]
- Massé, E.; Vanderpool, C.K.; Gottesman, S. Effect of RyhB Small RNA on Global Iron Use in Escherichia coli. J. Bacteriol. 2005, 187, 6962–6971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, P.; Li, Y.; Osborne, M.J.; Pomerantseva, E.; Liu, Q.; Gehring, K. Solution Structure of the Carbon Storage Regulator Protein CsrA from Escherichia coli. J. Bacteriol. 2005, 187, 3496–3501. [Google Scholar] [CrossRef] [Green Version]
- Timmermans, J.; Van Melderen, L. Post-transcriptional global regulation by CsrA in bacteria. Cell. Mol. Life Sci. 2010, 67, 2897–2908. [Google Scholar] [CrossRef]
- Dobrzanski, T.; Pobre, V.; Moreno, L.F.; Barbosa, H.C.D.S.; Monteiro, R.A.; Pedrosa, F.D.O.; De Souza, E.M.; Arraiano, C.M.; Steffens, M.B.R. In silico prediction and expression profile analysis of small non-coding RNAs in Herbaspirillum seropedicae SmR1. BMC Genom. 2020, 21, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Söderbom, F.; Wagner, E.G.H.; Binnie, U.; Binns, N.; Masters, M. PcnB is required for the rapid degradation of RNAI, the antisense RNA that controls the copy number of CoIE1-related plasmids. Mol. Microbiol. 1993, 9, 1131–1142. [Google Scholar] [CrossRef] [PubMed]
- Schmidtke, C.; Findeiss, S.; Sharma, C.M.; Kuhfuß, J.; Hoffmann, S.; Vogel, J.; Stadler, P.F.; Bonas, U. Genome-wide transcriptome analysis of the plant pathogen Xanthomonas identifies sRNAs with putative virulence functions. Nucleic Acids Res. 2011, 40, 2020–2031. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. salmonicida Subspecies | Geographic Origin, Host Specie | Accession Number | N° of Plasmids | References |
|---|---|---|---|---|
| salmonicida A449 | France/ Salmo gairdneri | CP000644-46 | 5 | [12,42,43] |
| salmonicida J223 (Wild-type 3.173) | Pittsford, VT, USA, 1999/ Salmo salar | CP048223-27 | 4 | [26]/This study |
| salmonicida J409 | Canada/ Anoplopoma fimbria | CP047374-75 | 1 | This study |
| salmonicida J410 | Canada/ Anoplopoma fimbria | CP047376-77 | 1 | This study |
| salmonicida J411 | Canada/ Anoplopoma fimbria | CP052034-35 | 1 | This study |
| salmonicida O1-B526 | Canada/ Salvelinus fontinalis | CP027000 | 6 | [44,45] |
| salmonicida SHY16-3432 | Canada/ Salvelinus fontinalis | CP038102-105 | 7 | [46] |
| pectinolytica 34mel | Argentina/ environmental sample | CP022426 | - | [47] |
| salmonicida S44 | China/ Salmo salar | CP022176-81 | 5 | - |
| salmonicida S121 | China/ Salmo salar | CP022170-75 | 5 | - |
| salmonicida S68 | China/ Salmo salar | CP022182-86 | 4 | - |
| masoucida BR19001YR | South Korea/ Sebastes schlegeli | CP060030-33 | 3 | - |
| salmonicida SRW-OG1 | China/ Epinephelus coioiaes | CP051883 | - | - |
| salmonicida A527 | India/Macrobrachium rosenbergii | CP022550 | - | [48] |
| salmonicida O23A | Poland/ environmental sample | CP021654-58 | 4 | [49] |
| masoucida RAFS1 | South Korea/ Sebastes schlegelii | CP017143-45 | 2 | [50] |
| Pseudomonas putida KT2440 | France/ Environmental sample | NC_002947 | - | [51] |
| Characteristic | J409 | J410 | J411 | J223 |
|---|---|---|---|---|
| Grown in TSB at: | ||||
| 4 °C | + | + | + | + |
| 15 °C | + | + | + | + |
| 28 °C | + | + | + | + |
| 37 °C | - | - | - | - |
| LB NaCl 0% | - | - | - | - |
| LB NaCl 0.5% | + | + | + | + |
| LB NaCl 2% | + | + | + | + |
| Motility | - | - | - | - |
| Fimbria type I | - | - | - | - |
| Siderophores synthesis | + | + | + | + |
| Hemolytic activity | + | + | + | + |
| Catalase | + | + | - | + |
| Oxidase | + | + | + | + |
| Antibiogram | mm | mm | mm | mm |
| Tetracycline (10 µg) | 38 (Susceptible) | 36 (Susceptible) | 16 (Susceptible) | 31 (Susceptible) |
| Oxytetracycline (30 µg) | 42 (Susceptible) | 40 (Susceptible) | 40 (Susceptible) | 32 (Susceptible) |
| Ampicillin (10 µg) | 0 (Resistant) | 0 (Resistant) | 16 (Susceptible) | 0 (Resistant) |
| Sulfamethoxazole (25 µg) | 24 (Susceptible) | 26 (Susceptible) | 39 (Susceptible) | 24 (Susceptible) |
| Chloramphenicol (30 µg) | 40 (Susceptible) | 36 (Susceptible) | 44 (Susceptible) | 13 (Susceptible) |
| Colistin sulphate (10 µg) | 12 (Susceptible) | 18 (Susceptible) | 21 (Susceptible) | 16 (Susceptible) |
| Vibriostatic O-129 (150 µg) | 0 (Resistant) | 0 (Resistant) | 0 (Resistant) | 0 (Resistant) |
| Oxolinic acid (2 µg) | 36 (Susceptible) | 42 (Susceptible) | 18 (Susceptible) | 34 (Susceptible) |
| Attribute | Data Provided | |||
|---|---|---|---|---|
| Annotation Pipeline | NCBI prokaryotic Genome Annotation pipeline | |||
| Annotation Method | Best placed reference protein set; GeneMarks v 4.10 | |||
| A. salmonicida genome | J409 | J410 | J411 | J223 |
| Genes (total) | 4532 | 4546 | 4586 | 4.626 |
| CDSs (total) | 4381 | 4395 | 4430 | 4.478 |
| Genes (coding) | 3948 | 3985 | 3456 | 4.207 |
| Genes (RNA) | 151 | 151 | 156 | 148 |
| rRNAs | 11, 10, 10 (5S, 16S, 23S) | 11, 10, 10 (5S, 16S, 23S) | 11, 10, 10 (5S, 16S, 23S) | 10, 9, 9 (5S, 16S, 23S) |
| Complete rRNAs | 11, 10, 10 (5S, 16S, 23S) | 11, 10, 10 (5S, 16S, 23S) | 11, 10, 10 (5S, 16S, 23S) | 10, 9, 9 (5S, 16S, 23S) |
| tRNAs | 116 | 116 | 121 | 116 |
| ncRNAs | 4 | 4 | 4 | 4 |
| Pseudogenes (total) | 433 | 410 | 974 | 271 |
| Pseudogenes (ambiguous residues) | 0 of 433 | 0 of 410 | 0 of 974 | 1 of 271 |
| Pseudogenes (frameshifted) | 182 of 433 | 162 of 410 | 728 of 974 | 157 of 271 |
| Pseudogenes (incomplete) | 250 of 433 | 243 of 410 | 263 of 974 | 90 of 271 |
| Pseudogenes (internal stop) | 40 of 433 | 40 of 410 | 45 of 974 | 47 of 271 |
| Pseudogenes (multiple problems) | 37 of 433 | 33 of 410 | 59 of 974 | 22 of 271 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasquez, I.; Hossain, A.; Gnanagobal, H.; Valderrama, K.; Campbell, B.; Ness, M.; Charette, S.J.; Gamperl, A.K.; Cipriano, R.; Segovia, C.; et al. Comparative Genomics of Typical and Atypical Aeromonas salmonicida Complete Genomes Revealed New Insights into Pathogenesis Evolution. Microorganisms 2022, 10, 189. https://doi.org/10.3390/microorganisms10010189
Vasquez I, Hossain A, Gnanagobal H, Valderrama K, Campbell B, Ness M, Charette SJ, Gamperl AK, Cipriano R, Segovia C, et al. Comparative Genomics of Typical and Atypical Aeromonas salmonicida Complete Genomes Revealed New Insights into Pathogenesis Evolution. Microorganisms. 2022; 10(1):189. https://doi.org/10.3390/microorganisms10010189
Chicago/Turabian StyleVasquez, Ignacio, Ahmed Hossain, Hajarooba Gnanagobal, Katherinne Valderrama, Briony Campbell, Michael Ness, Steve J. Charette, Anthony K. Gamperl, Rocco Cipriano, Cristopher Segovia, and et al. 2022. "Comparative Genomics of Typical and Atypical Aeromonas salmonicida Complete Genomes Revealed New Insights into Pathogenesis Evolution" Microorganisms 10, no. 1: 189. https://doi.org/10.3390/microorganisms10010189
APA StyleVasquez, I., Hossain, A., Gnanagobal, H., Valderrama, K., Campbell, B., Ness, M., Charette, S. J., Gamperl, A. K., Cipriano, R., Segovia, C., & Santander, J. (2022). Comparative Genomics of Typical and Atypical Aeromonas salmonicida Complete Genomes Revealed New Insights into Pathogenesis Evolution. Microorganisms, 10(1), 189. https://doi.org/10.3390/microorganisms10010189