
Whole-Genome Sequencing Characterization of Virulence Profiles of Listeria monocytogenes Food and Human Isolates and In Vitro Adhesion/Invasion Assessment

 , , , , , , , , ,
, , , , , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Whole-Genome Sequencing (WGS)
2.2.1. Multi Locus Sequence Typing (MLST)
2.2.2. Virulence-Associated Genes Detection
2.3. In Vitro Assays
2.3.1. Epithelial Cell Line
2.3.2. Adhesion Assay
2.3.3. Invasion Assay
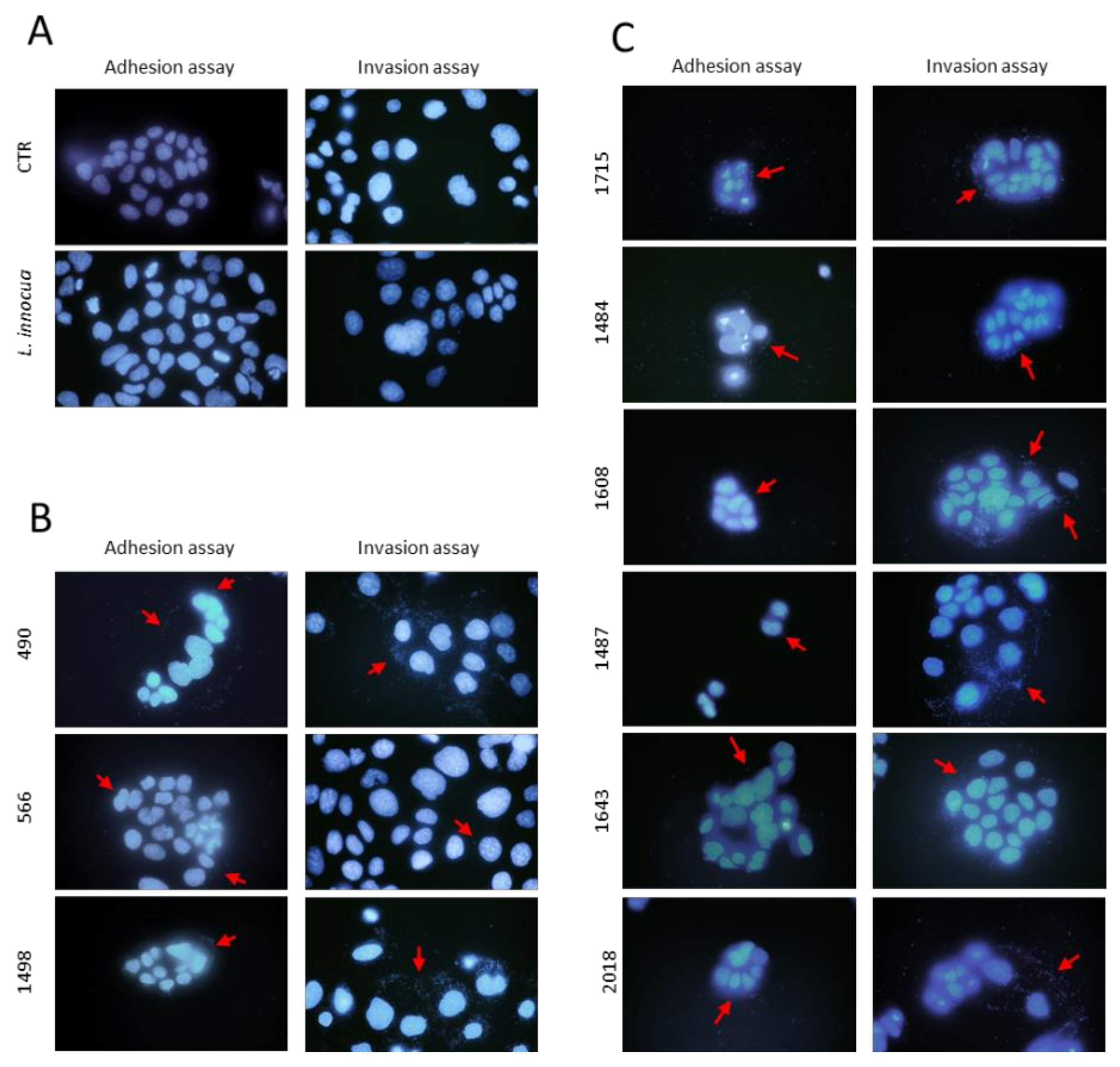
2.3.4. Hoechst Staining
2.3.5. Statistical Analysis
3. Results
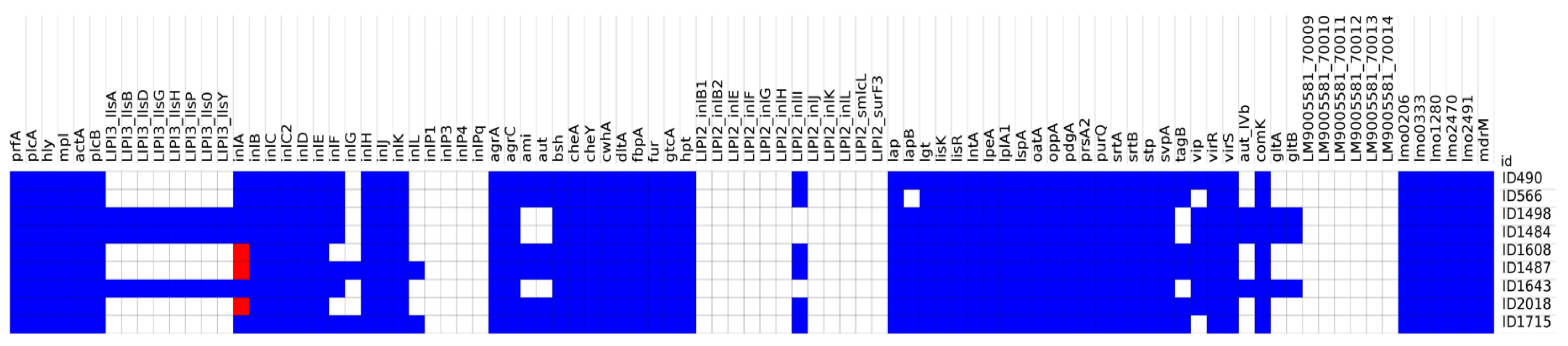
3.1. Whole-Genome Sequencing (WGS) and Bioinformatics Analysis
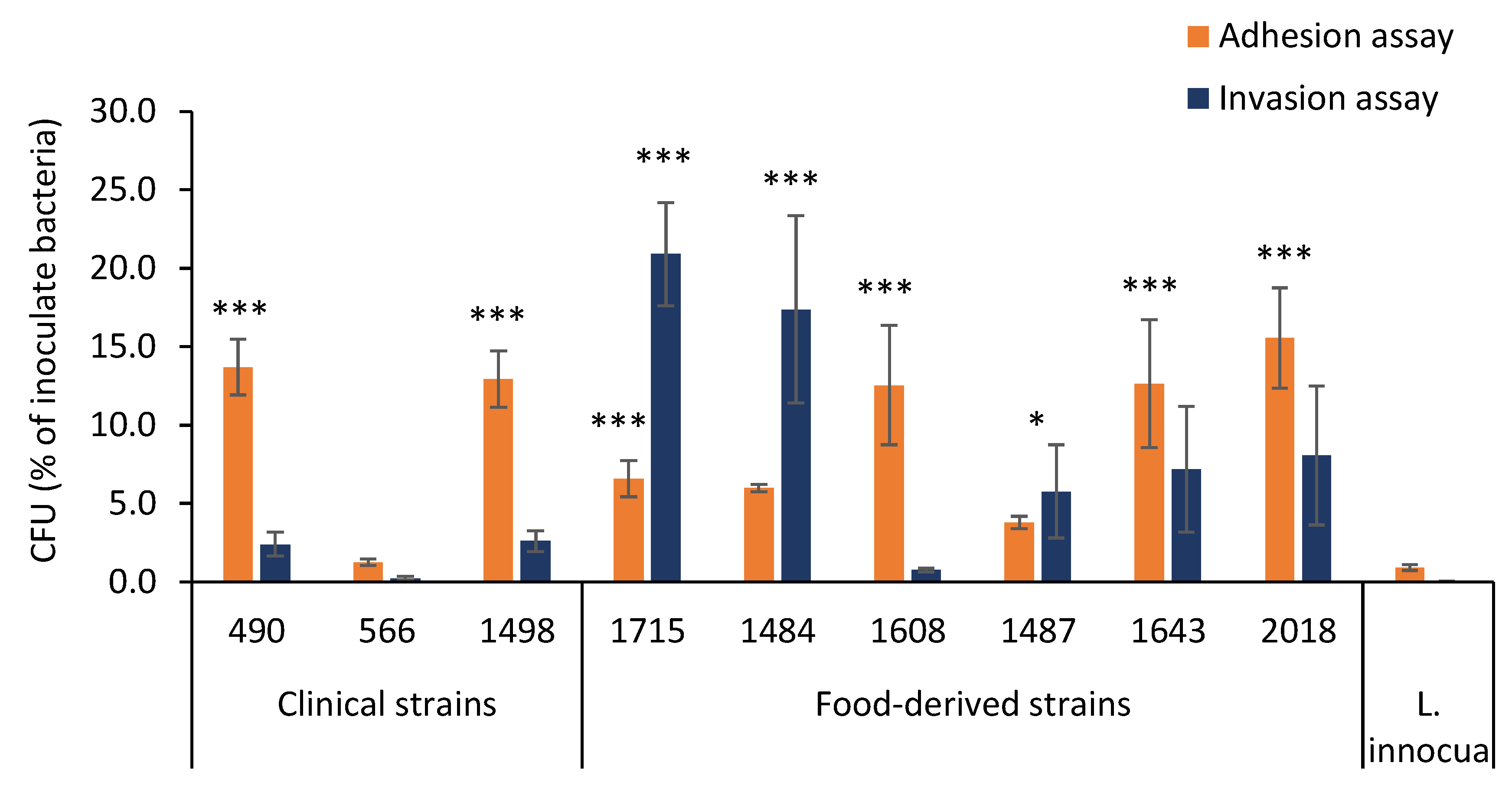
3.2. Adhesion and Invasion
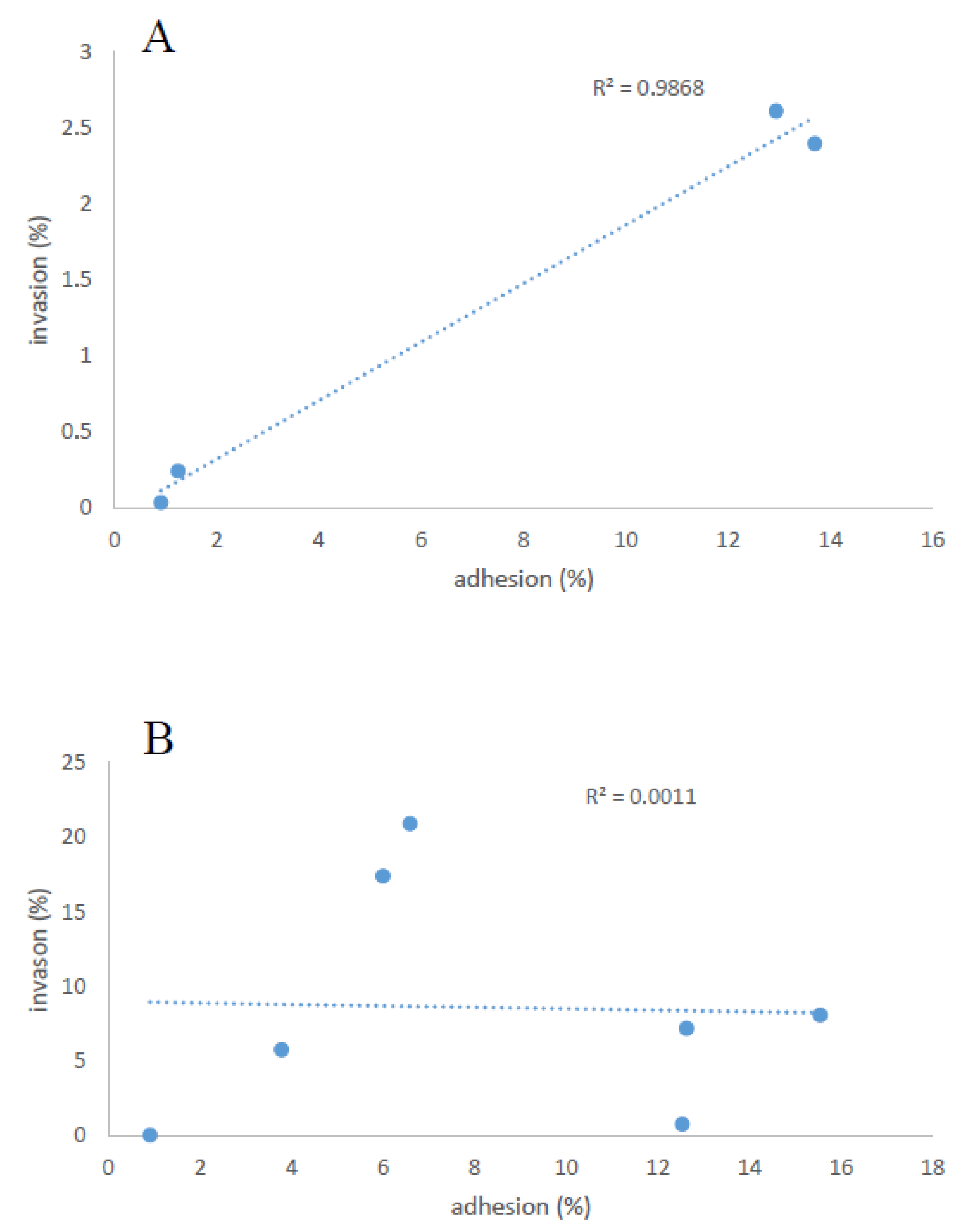
Correlation between Adhesion and Invasion Properties of All Strains
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, 6406. [Google Scholar]
- Radoshevich, L.; Cossart, P. Listeria monocytogenes: Towards a complete picture of its physiology and pathogenesis. Nat. Rev. Microbiol. 2018, 16, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Barbuddhe, S.B.; Chakraborty, T. Listeria as an enteroinvasive gastrointestinal pathogen. Curr. Top Microbiol. Immunol. 2009, 337, 173–195. [Google Scholar]
- Lomonaco, S.; Decastelli, L.; Nucera, D.; Gallina, S.; Bianchi, D.M.; Civera, T. Listeria monocytogenes in Gorgonzola: Subtypes, diversity and persistence overtime. Int. J. Food Microbiol. 2009, 128, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Kaptchouang Tchatchouang, C.D.; Fri, J.; De Santi, M.; Brandi, G.; Schiavano, G.F.; Amagliani, G.; Ateba, C.N. Listeriosis Outbreak in South Africa: A Comparative Analysis with Previously Reported Cases Worldwide. Microorganisms 2020, 8, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecuit, M. Listeria monocytogenes, a model in infection biology. Cell. Microbiol. 2020, 22, e13186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaminathan, B.; Gerner-Smidt, P. The epidemiology of human listeriosis. Microbes Infect. 2007, 9, 1236–1243. [Google Scholar] [CrossRef] [Green Version]
- Loo, K.Y.; Letchumanan, V.; Dhanoa, A.; Law, J.W.F.; Pusparajah, P.; Goh, B.H.; Ser, H.L.; Wong, S.H.; Ab Mutalib, N.S.; Chan, K.G.; et al. Exploring the Pathogenesis, Clinical Characteristics and Therapeutic Regimens of Listeria monocytogenes. Microbiology 2020, 3, 1–13. [Google Scholar]
- Xiao-Yong, Z.; Chao-Hui, H.; Qing-Yi, Z. Legionella Pathogenesis and Virulence Factors. Ann. Clin. Lab. Res. 2015, 3, 15. [Google Scholar]
- Su, X.; Cao, G.; Zhang, J.; Pan, H.; Zhang, D.; Kuang, D.; Yang, X.; Xu, X.; Shi, X.; Meng, J. Characterization of internalin genes in Listeria monocytogenes from food and humans, and their association with the invasion of Caco-2 cells. Gut Pathog. 2019, 11, 11–30. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, Y.; Pouillot, R.; Dennis, S.; Xian, Z.; Luchansky, J.B. Genetic diversity and profiles of genes associated with virulence and stress resistance among isolates from the 2010–2013 interagency Listeria monocytogenes market basket survey. PLoS ONE 2020, 15, e0231393. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, W.; Wang, J.; Xu, B.; Liu, H.; Dong, Q.; Zhang, X. 10-Year Molecular Surveillance of Listeria monocytogenes Using Whole-Genome Sequencing in Shanghai, China, 2009–2019. Front. Microbiol. 2020, 11, 3037. [Google Scholar] [CrossRef]
- Vilchis-Rangel, R.E.; Espinoza-Mellado, M.D.R.; Salinas-Jaramillo, I.J.; Martinez-Peña, M.D.; Rodas-Suárez, O.R. Association of Listeria monocytogenes LIPI-1 and LIPI-3 marker llsX with invasiveness. Curr. Microbiol. 2019, 76, 637–643. [Google Scholar] [CrossRef]
- De Melo Tavares, R.; da Silva, D.A.L.; Camargo, A.C.; Yamatogi, R.S.; Nero, L.A. Interference of the acid stress on the expression of llsX by Listeria monocytogenes pathogenic island 3 (LIPI-3) variants. Food Res. Int. 2020, 132, 109063. [Google Scholar] [CrossRef]
- Disson, O.; Moura, A.; Lecuit, M. Making Sense of the Biodiversity and Virulence of Listeria monocytogenes. Trends Microbiol. 2021, 11, 811–822. [Google Scholar] [CrossRef]
- Cotter, P.D.; Draper, L.A.; Lawton, E.M.; Daly, K.M.; Groeger, D.S.; Casey, P.G.; Ross, R.P.; Hill, C. Listeriolysin S, a Novel Peptide Haemolysin Associated with a Subset of Lineage I Listeria monocytogenes. PLoS Pathog. 2008, 4, e1000144. [Google Scholar] [CrossRef] [Green Version]
- Maury, M.M.; Tsai, Y.H.; Charlier, C.; Touchon, M.; Chenal-Francisque, V.; Leclercq, A.; Criscuolo, A.; Gautier, C.; Roussel, S.; Brisabois, A. Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat. Genet. 2016, 48, 308–313. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Cheng, J.; Zhang, J.; Chen, Y.; Zeng, H.; Xue, L.; Lei, T.; Pang, R.; Wu, S.; Wu, H.; et al. Isolation, Potential Virulence, and Population Diversity of Listeria monocytogenes From Meat and Meat Products in China. Front. Microbiol. 2019, 7, 946. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chen, M.; Wang, J.; Wu, Q.; Cheng, J.; Zhang, J.; Sun, Q.; Xue, L.; Zeng, H.; Lei, T.; et al. Heterogeneity, Characteristics, and Public Health Implications of Listeria monocytogenes in Ready-to-Eat Foods and Pasteurized Milk in China. Front. Microbiol. 2020, 11, 642. [Google Scholar] [CrossRef] [Green Version]
- Van Stelten, A.; Simpson, J.M.; Ward, T.J.; Nightingale, K.K. Revelation by single-nucleotide polymorphism genotyping that mutations leading to a premature stop codon in inlA are common among Listeria monocytogenes isolates from ready-to-eat foods but not human listeriosis cases. Appl. Environ. Microbiol. 2010, 76, 2783–2790. [Google Scholar] [CrossRef] [Green Version]
- Kyoui, D.; Takahashi, H.; Miya, S.; Kuda, T.; Kimura, B. Comparison of the major virulence-related genes of Listeria monocytogenes in Internalin A truncated strain 36-25-1 and a clinical wild-type strain. BMC Microbiol. 2014, 14, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nightingale, K.K.; Ivy, R.A.; Ho, A.J.; Fortes, E.D.; Njaa, B.L.; Peters, R.M.; Wiedmann, M. inlA premature stop codons are common among Listeria monocytogenes isolates from foods and yield virulence-attenuated strains that confer protection against fully virulent strains. Appl. Environ. Microbiol. 2008, 74, 6570–6583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akgul, A.; Al-Janabi, N.; Das, B.; Lawrence, M.; Karsi, A. Small molecules targeting LapB protein prevent Listeria attachment to catfish muscle. PLoS ONE 2017, 12, e0189809. [Google Scholar] [CrossRef] [Green Version]
- Doumith, M.; Buchrieser, C.; Glaser, P.; Jacquet, C.; Martin, P. Differentiation of the major Listeria monocytogenes serovars by multiplex PCR differentiation of the major Listeria monocytogenes serovars by multiplex PCR. J. Clin. Microbiol. 2004, 42, 3819–3822. [Google Scholar] [CrossRef] [Green Version]
- Orsi, R.H.; den Bakker, H.C.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Montero, D.; Bodero, M.; Riveros, G.; Lapierre, L.; Gaggero, A.; Vidal, R.M.; Vidal, M. Molecular epidemiology and genetic diversity of Listeria monocytogenes isolates from a wide variety of ready-to-eat foods and their relationship to clinical strains from listeriosis outbreaks in Chile. Front. Microbiol. 2015, 6, 384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maury, M.M.; Bracq-Dieye, H.; Huang, L.; Vales, G.; Lavina, M.; Thouvenot, P.; Disson, O.; Leclercq, A.; Brisse, S.; Lecuit, M. Hypervirulent Listeria monocytogenes clones’ adaption to mammalian gut accounts for their association with dairy products. Nat. Commun. 2019, 10, 2488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Lawrence, M.L.; Ainsworth, A.J.; Austin, F.W. Toward an improved laboratory definition of Listeria monocytogenes virulence. Int. J. Food Microbiol. 2007, 118, 101–115. [Google Scholar] [CrossRef]
- Medeiros, M.; Castro, V.H.L.; Mota, A.; Pereira, M.G.; De Martinis, E.C.P.; Perecmanis, S.; Santana, A.P. Assessment of Internalin A Gene Sequences and Cell Adhesion and Invasion Capacity of Listeria monocytogenes Strains Isolated from Foods of Animal and Related Origins. Foodborne Pathog. Dis. 2021, 18, 243–252. [Google Scholar] [CrossRef]
- Reddy, S.; Akgul, A.; Karsi, A.; Abdelhamed, H.; Wills, R.W.; Lawrence, M.L. The role of Listeria monocytogenes cell wall surface anchor protein LapB in virulence, adherence, and intracellular replication. Microb. Pathog. 2016, 92, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Union. Commission Regulation (EC) No 2073/2005 of 15 November 2005 on Microbiological Criteria for Foodstuffs; European Union: Brussels, Belgium, 2005. [Google Scholar]
- European Union. Commission Regulation (EC) No 1441/2007 of 5 December 2007 amending Regulation (EC) No 2073/2005 on Microbiological Criteria for Foodstuffs; European Union: Brussels, Belgium, 2007. [Google Scholar]
- Portmann, A.C.; Fournier, C.; Gimonet, J.; Ngom-Bru, C.; Barretto, C.; Baert, L. Validation Approach of an End-to-End Whole Genome Sequencing Workflow for Source Tracking of Listeria monocytogenes and Salmonella enterica. Front. Microbiol. 2018, 14, 446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cito, F.; Di Pasquale, A.; Cammà, C.; Cito, P. The Italian information system for the collection and analysis of complete genome sequence of pathogens isolated from animal, food and environment. Int. J. Infect. Dis. 2018, 73, 296–297. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Salcedo, C.; Arreaza, L.; Alcalá, B.; de la Fuente, L.; Vázquez, J.A. Development of a multilocus sequence typing method for analysis of Listeria monocytogenes clones. J. Clin. Microbiol. 2003, 41, 757–762. [Google Scholar] [CrossRef] [Green Version]
- Moura, A.; Criscuolo, A.; Pouseele, H.; Maury, M.M.; Leclercq, A.; Tarr, C.; Björkman, J.T.; Dallman, T.; Reimer, A.; Enouf, V.; et al. Whole genome-based population biology and epidemiological surveillance of Listeria monocytogenes. Nat. Microbiol. 2016, 10, 16185. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Austin, F. Adhesion and Invasion Assay Procedure Using Caco-2 Cells for Listeria monocytogenes. Bio-Protocol 2017, 7, e2267. [Google Scholar] [CrossRef] [PubMed]
- Timme, R.E.; Wolfgang, W.J.; Balkey, M. Optimizing open data to support one health: Best practices to ensure interoperability of genomic data from bacterial pathogens. One Health Outlook 2020, 20, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Amagliani, G.; Blasi, G.; Scuota, S.; Duranti, A.; Fisichella, S.; Gattuso, A.; Gianfranceschi, M.V.; Schiavano, G.F.; Brandi, G.; Pomilio, F.; et al. Detection and Virulence Characterization of Listeria monocytogenes Strains in Ready-to-Eat Products. Foodborne Pathog. Dis. 2021, 18, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Olier, M.; Pierre, F.; Rousseaux, S.; Lemaître, J.P.; Rousset, A.; Piveteau, P.; Guzzo, J. Expression of truncated Internalin A is involved in impaired internalization of some Listeria monocytogenes isolates carried asymptomatically by humans. Infect. Immun. 2003, 71, 1217–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guidi, F.; Orsini, M.; Chiaverini, A.; Torresi, M.; Centorame, P.; Acciari, V.A.; Salini, R.; Palombo, B.; Brandi, G.; Amagliani, G.; et al. Hypo- and Hyper-Virulent Listeria monocytogenes Clones Persisting in Two Different Food Processing Plants of Central Italy. Microorganisms 2021, 13, 376. [Google Scholar] [CrossRef]
- Painset, A.; Björkman, J.T.; Kiil, K.; Guillier, L.; Mariet, J.F.; Félix, B.; Amar, C.; Rotariu, O.; Roussel, S.; Perez-Reche, F.; et al. LiSEQ—whole-genome sequencing of a cross-sectional survey of Listeria monocytogenes in ready-to-eat foods and human clinical cases in Europe. Microb. Genom. 2019, 5, e000257. [Google Scholar] [CrossRef] [PubMed]
- Hilliard, A.; Leong, D.; O’Callaghan, A.; Culligan, E.P.; Morgan, C.A.; DeLappe, N.; Hill, C.; Jordan, K.; Cormican, M.; Gahan, C.G.M. Genomic Characterization of Listeria monocytogenes Isolates Associated with Clinical Listeriosis and the Food Production Environment in Ireland. Genes 2018, 9, 171. [Google Scholar] [CrossRef] [Green Version]
- Lüth, S.; Halbedel, S.; Rosner, B.; Wilking, H.; Holzer, A.; Roedel, A.; Dieckmann, R.; Vincze, S.; Prager, R.; Flieger, A. Backtracking and forward checking of human listeriosis clusters identified a multiclonal outbreak linked to Listeria monocytogenes in meat products of a single producer. Emerg. Microbes Infect. 2020, 9, 1600–1608. [Google Scholar] [CrossRef]
- Duranti, A.; Sabbatucci, M.; Blasi, G.; Acciari, V.A.; Ancora, M.; Bella, A.; Busani, L.; Centorame, P.; Cammà, C.; Conti, F.; et al. A severe outbreak of listeriosis in central Italy with a rare pulsotype associated with processed pork products. J. Med. Microbiol. 2018, 67, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Baba, H.; Kanamori, H.; Kakuta, R.; Sakurai, H.; Oshima, K.; Aoyagi, T.; Kaku, M. Genomic characteristics of listeria monocytogenes causing invasive listeriosis in Japan. Diagn. Microbiol. Infect. Dis. 2021, 99, 115233. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID | Source | Serotype | |
|---|---|---|---|
| Human | 490 | Blood | 1/2a |
| 566 | Blood | 1/2a | |
| 1498 | Cerebrospinal fluid | 4b | |
| RTE-food | 1484 | “Coppa di testa” head cheese | 1/2b |
| 1608 | “Coppa di testa” head cheese | 1/2a | |
| 1487 | Fresh salami | 4b | |
| 1643 | Salami | 4b | |
| 2018 | Spit roasted pork | 1/2a | |
| 1715 | “Coppa di testa” head cheese | 1/2a |
| ID | Vertical Coverage | N° Contigs | Total Length (bp) | N50 | L50 |
|---|---|---|---|---|---|
| Lm_490 | 105.7 | 46 | 3,023,546 | 308,142 | 3 |
| Lm_566 | 515.12 | 59 | 3,082,646 | 417,896 | 3 |
| Lm_1498 | 219.6 | 130 | 2,945,468 | 556,758 | 2 |
| Lm_1484 | 74 | 50 | 2,927,103 | 147,035 | 1 |
| Lm_1487 | 104 | 111 | 3,079,929 | 524,763 | 3 |
| Lm_1608 | 123 | 52 | 3,024,307 | 563,871 | 2 |
| Lm_1643 | 92.4 | 61 | 3,023,637 | 580,655 | 2 |
| Lm_1715 | 133 | 40 | 2,934,721 | 437,349 | 2 |
| Lm_2018 | 51.1 | 71 | 3,123,917 | 531,830 | 2 |
| ID Strain | Isolation Source | CC | InlAAllele | PMSC | PMSC Type | InlA Type | Mutation Position | InlA Length |
| 490 | human | CC101 | 21 | - | - | Full length | 800 aa | |
| 566 | human | CC31 | 153 | - | - | Full length | 800 aa | |
| 1498 | human | CC1 | 3 | - | - | Full length | 800 aa | |
| 1484 | food | CC1 | 3 | - | - | Full length | 800 aa | |
| 1487 | food | CC9 | 47 | + | 29 | Truncated | 1635 (deletion A) | 576 aa |
| 1608 | food | CC121 | 49 | + | 6 | Truncated | 1474 (C T) | 491 aa |
| 1643 | food | CC1 | 3 | - | - | Full length | 800 aa | |
| 1715 | food | CC7 | 2 | - | - | Full length | 800 aa | |
| 2018 | food | CC121 | 49 | + | 6 | Truncated | 1474 (C T) | 491 aa |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schiavano, G.F.; Ateba, C.N.; Petruzzelli, A.; Mele, V.; Amagliani, G.; Guidi, F.; De Santi, M.; Pomilio, F.; Blasi, G.; Gattuso, A.; et al. Whole-Genome Sequencing Characterization of Virulence Profiles of Listeria monocytogenes Food and Human Isolates and In Vitro Adhesion/Invasion Assessment. Microorganisms 2022, 10, 62. https://doi.org/10.3390/microorganisms10010062
Schiavano GF, Ateba CN, Petruzzelli A, Mele V, Amagliani G, Guidi F, De Santi M, Pomilio F, Blasi G, Gattuso A, et al. Whole-Genome Sequencing Characterization of Virulence Profiles of Listeria monocytogenes Food and Human Isolates and In Vitro Adhesion/Invasion Assessment. Microorganisms. 2022; 10(1):62. https://doi.org/10.3390/microorganisms10010062
Chicago/Turabian StyleSchiavano, Giuditta Fiorella, Collins Njie Ateba, Annalisa Petruzzelli, Veronica Mele, Giulia Amagliani, Fabrizia Guidi, Mauro De Santi, Francesco Pomilio, Giuliana Blasi, Antonietta Gattuso, and et al. 2022. "Whole-Genome Sequencing Characterization of Virulence Profiles of Listeria monocytogenes Food and Human Isolates and In Vitro Adhesion/Invasion Assessment" Microorganisms 10, no. 1: 62. https://doi.org/10.3390/microorganisms10010062
APA StyleSchiavano, G. F., Ateba, C. N., Petruzzelli, A., Mele, V., Amagliani, G., Guidi, F., De Santi, M., Pomilio, F., Blasi, G., Gattuso, A., Di Lullo, S., Rocchegiani, E., & Brandi, G. (2022). Whole-Genome Sequencing Characterization of Virulence Profiles of Listeria monocytogenes Food and Human Isolates and In Vitro Adhesion/Invasion Assessment. Microorganisms, 10(1), 62. https://doi.org/10.3390/microorganisms10010062