
Fungal Communities in Leaves and Roots of Healthy-Looking and Diseased Ulmus glabra

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
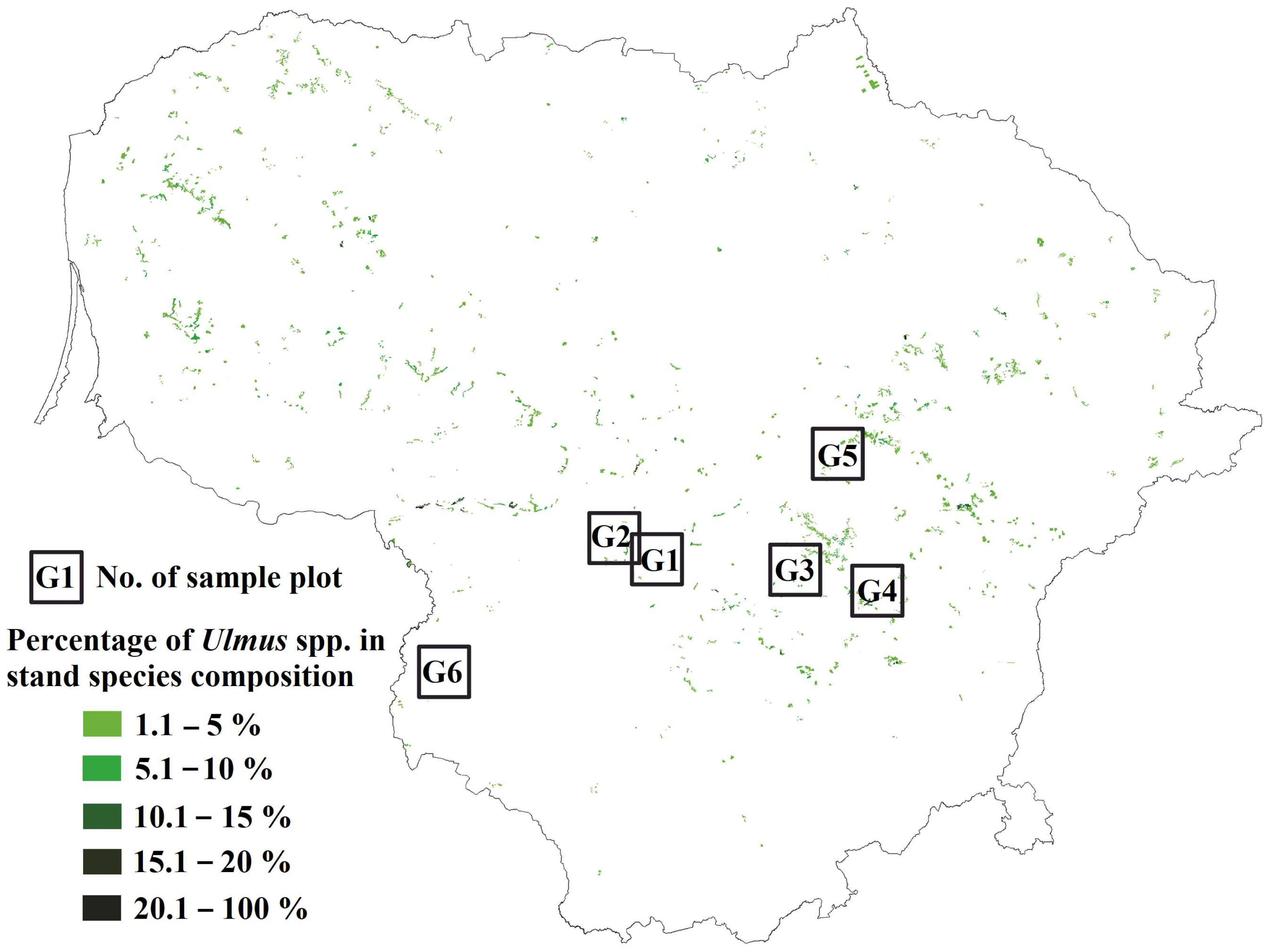
2.1. Study Sites and Sampling
2.2. DNA Isolation, Amplification, and Sequencing
2.3. Bioinformatics
2.4. Statistical Analyses
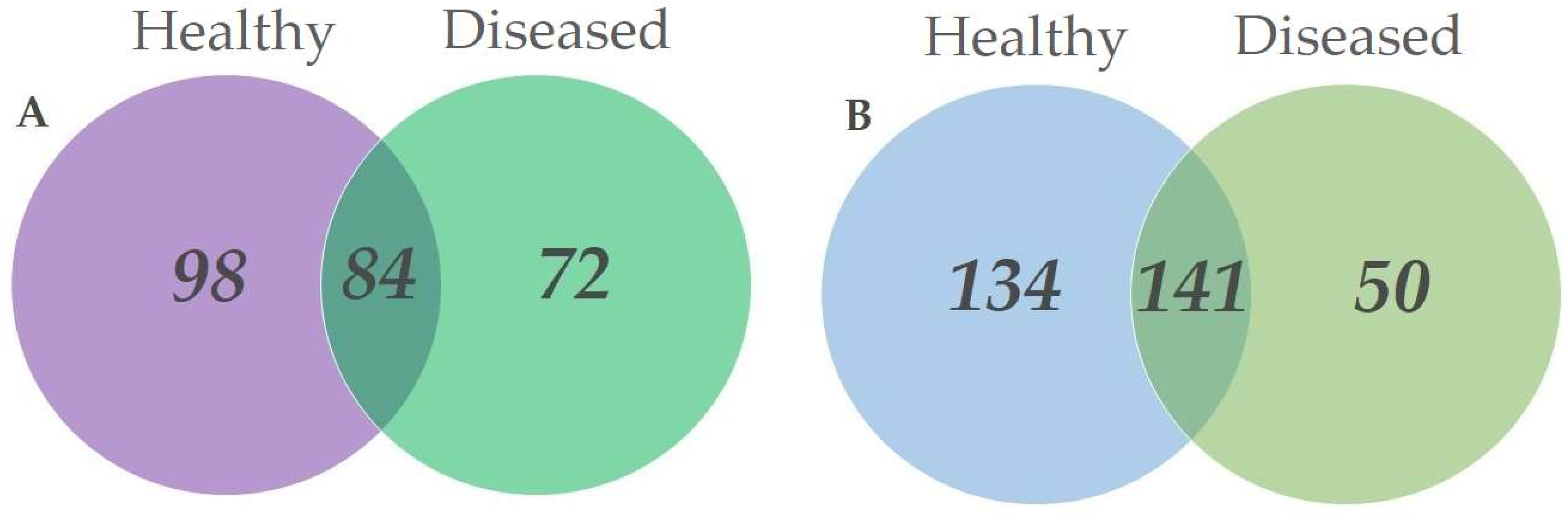
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jürisoo, L.; Adamson, K.; Padari, A.; Drenkhan, R. Health of elms and Dutch elm disease in Estonia. Eur. J. Plant Pathol. 2019, 154, 823–841. [Google Scholar] [CrossRef]
- Motiejūnaitė, J.; Kutorga, E.; Kasparavičius, J.; Lygis, V.; Norkutė, G. New records from Lithuania of fungi alien to Europe. Mycotaxon 2016, 131, 49–60. [Google Scholar] [CrossRef]
- Thor, G.; Johansson, P.; Jönsson, M.T. Lichen diversity and red-listed lichen species relationships with tree species and diameter in wooded meadows. Biodivers. Conserv. 2010, 19, 2307–2328. [Google Scholar] [CrossRef]
- Richens, R.H. Elm; Cambridge University Press: Cambridge, UK, 1983; p. 347. [Google Scholar]
- Heybroek, H.M. The Dutch Elm Breeding Program. In Dutch Elm Disease Research; Sticklen, M.B., Sherald, J.L., Eds.; Springer: New York, NY, USA, 1993; pp. 16–25. [Google Scholar] [CrossRef]
- Jüriado, I.; Liira, J.; Paal, J.; Suija, A. Tree and stand level variables influencing diversity of lichens on temperate broad-leaved trees in boreo-nemoral floodplain forests. Biodivers. Conserv. 2009, 18, 105–125. [Google Scholar] [CrossRef]
- Corfixen, P.; Parmasto, E. Hymenochaete ulmicola, sp.nov. (Hymenochaetales). Mycotaxon 2005, 91, 465–470. [Google Scholar]
- Kalamees, K.; Hausknecht, A.; Vauras, J. Checklist of the genera Conocybe and Pholiotina (Agaricales, Agaricomycetes) in Estonia. Folia Cryptogam. Est. 2013, 50, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C.M. Ophiostoma novo-ulmi sp. nov., causative agent of current Dutch elm disease pandemics. Mycopathologia 1991, 115, 151–161. [Google Scholar] [CrossRef]
- Brasier, C.M. Intercontinental spread and continuing evolution of the Dutch elm disease pathogens. In The Elms: Breeding, Conservation and Disease Management; Dunne, C.P., Ed.; Kluwer Academic Publishers: Boston, MA, USA, 2000; pp. 61–72. [Google Scholar]
- Yadeta, K.A.; Thomma, B.P.J. The xylem as battleground for plant hosts and vascular wilt pathogens. Front. Plant Sci. 2013, 4, 97. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, J.N.; Brasier, C.M. Correlation between cultural characters and pathogenicity in Ceratocystis ulmi from Britain, Europe and America. Nature 1973, 241, 381–383. [Google Scholar] [CrossRef]
- Paoletti, M.; Buck, K.W.; Brasier, C.M. Cloning and sequence analysis of the MA TB (MAT-2) genes from the three Dutch elm disease pathogens, Ophiostoma ulmi, O. novo-ulmi and O. himal-ulmi. Mycol. Res. 2005, 109, 983–991. [Google Scholar] [CrossRef]
- Miyamoto, T.; Masuya, H.; Koizumi, A.; Yamaguchi, T.; Ishihara, M.; Yamaoka, Y.; Ohara, M. A report of dieback and mortality of elm trees suspected of Dutch elm disease in Hokkaido, Japan. J. For. Res. 2019, 24, 396–400. [Google Scholar] [CrossRef]
- Brasier, C.M. A cytoplasmically transmitted disease of Ceratocystis ulmi. Nature 1983, 305, 220–223. [Google Scholar] [CrossRef]
- Fransen, J.J. Elm Disease, Elm Beetles and Their Control. PhD Thesis, Agricultural University, Wageningen, The Netherlands, 1939. [Google Scholar]
- Webber, J.F. Experimental studies on factors influencing the transmission of Dutch elm disease. For. Syst. 2004, 13, 197–205. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Xue, H.; Pritchard, H.W.; Wang, X. Changes in the mitochondrial protein profile due to ROS eruption during ageing of elm (Ulmus pumila L.) seeds. Plant Physiol. Biochem. 2017, 114, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Venturas, M.; Nanos, N.; Gil, L. The reproductive ecology of Ulmus laevis Pallas in a transformed habitat. For. Ecol. Manag. 2014, 312, 170–178. [Google Scholar] [CrossRef]
- Santini, A.; Faccoli, M. Dutch elm disease and elm bark beetles: A century of association. iFor.-Biogeosci. For. 2015, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Anderbrant, O.; Yuvaraj, J.K.; Martin, J.A.; Gil, L.; Witzell, J. Feeding by Scolytus bark beetles to test for differently susceptible elm varieties. J. Appl. Entomol. 2017, 141, 417–420. [Google Scholar] [CrossRef]
- Liu, L.; Yang, C.; Xu, X.; Wang, X.; Liu, M.; Chen, R.; Tan, F.; Liu, Y.; Lin, T.; Liu, Y. Unlocking the Changes of Phyllosphere Fungal Communities of Fishscale Bamboo (Phyllostachys heteroclada) under Rhombic-Spot Disease Stressed Conditions. Forests 2022, 13, 185. [Google Scholar] [CrossRef]
- Martín, J.A.; Witzell, J.; Blumenstein, K.; Rozpedowska, E.; Helander, M.; Sieber, T.N.; Gil, L. Resistance to Dutch elm disease reduces presence of xylem endophytic fungi in elms (Ulmus spp.). PLoS ONE 2013, 8, e56987. [Google Scholar] [CrossRef] [Green Version]
- Witzell, J.; Martín, J.A.; Blumenstein, K. Ecological aspects of endophyte-based biocontrol of forest diseases. In Advances in Endophytic Research; Verma, V.C., Gange, A.C., Eds.; Springer: New Delhi, India, 2014; pp. 321–333. [Google Scholar]
- Busby, P.E.; Ridout, M.; Newcombe, G. Fungal endophytes: Modifiers of plant disease. Plant. Mol. Biol. 2016, 90, 645–655. [Google Scholar] [CrossRef]
- Terhonen, E.; Blumenstein, K.; Kovalchuk, A.; Asiegbu, F.O. Forest tree microbiomes and associated fungal endophytes: Functional roles and impact on forest health. Forests 2019, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Vaičys, M. Miško dirvožemių klasifikacija. In Lietuvos Dirvožemiai; Mokslas: Vilnius, Lithuania, 2001; pp. 1040–1043. (In Lithuanian) [Google Scholar]
- Karazija, S. Lietuvos Miško Tipai; Mokslas: Vilnius, Lithuania, 1988; pp. 46–48. [Google Scholar]
- Marčiulynas, A.; Marčiulynienė, D.; Mishcherikova, V.; Franić, I.; Lynikienė, J.; Gedminas, A.; Menkis, A. High Variability of Fungal Communities Associated with the Functional Tissues and Rhizosphere Soil of Picea abies in the Southern Baltics. Forests 2022, 13, 1103. [Google Scholar] [CrossRef]
- Ihrmark, K.; Bodeker, I.T.M.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J.; Strid, Y.; Stenlid, J.; Brandstrom-Durling, M.; Clemmensen, K.E.; et al. New primers to amplify the fungal ITS2—Evaluation by 454-sequencing of artificial and natural communities. FEMS Microbiol. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988; 192p. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Community Ecology Package. R Package Version 2. 2013. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 5 July 2022).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Sieber, T.N. Endophytic fungi in forest trees: Are they mutualists? Fungal Biol. Rev. 2007, 21, 75–89. [Google Scholar] [CrossRef]
- Gonthier, P.; Gennaro, M.; Nicolotti, G. Effects of water stress on the endophytic mycota of Quercus robur. Fungal Diver. 2006, 21, e80. [Google Scholar]
- Speer, J.H.; Grissino-Mayer, H.D.; Orvis, K.H.; Greenberg, C.H. Climate response of five oak species in the eastern deciduous forest of the southern Appalachian Mountains, USA. Can. J. For. Res. 2009, 39, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Pureswaran, D.S.; Roques, A.; Battisti, A. Forest insects and climate change. Curr. For. Rep. 2018, 4, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, M.H.; Shin, K.C.; Lee, J.K. Fungal Community Analyses of Endophytic Fungi from Two Oak Species, Quercus mongolica and Quercus serrata, in Korea. Mycobiology 2021, 49, 385–395. [Google Scholar] [CrossRef]
- Agan, A.; Drenkhan, R.; Adamson, K.; Tedersoo, L.; Solheim, H.; Børja, I.; Matsiakh, I.; Timmermann, V.; Nagy, N.E.; Hietala, A.M. The relationship between fungal diversity and invasibility of a foliar niche—The case of ash dieback. J. Fungus 2020, 6, 150. [Google Scholar] [CrossRef]
- Barbier, S.; Balandier, P.; Gosselin, F. Influence of several tree traits on rainfall partitioning in temperate and boreal forests: A review. Ann. For. Sci. 2009, 66, 602. [Google Scholar] [CrossRef]
- Carnol, M.; Bazgir, M. Nutrient return to the forest floor through litter and throughfall under 7 forest species after conversion from Norway spruce. For. Ecol. Manag. 2013, 309, 66–75. [Google Scholar] [CrossRef]
- Augusto, L.; Achat, D.L.; Bakker, M.R.; Bernier, F.; Bert, D.; Danjon, F.; Khlifa, R.; Meredieu, C.; Trichet, P. Biomass and nutrients in tree root systems—Sustainable harvesting of an intensively managed Pinus pinaster (Ait.) planted forest. GCB Bioenergy 2015, 7, 231–243. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Reich, P.B.; Oleksyn, J.; Ogdahl, M.; Zytkowiak, R.; Hale, C.; Karolewski, P. Tree species effects on decomposition and forest floor dynamics in a common garden. Ecology 2006, 87, 2288–2297. [Google Scholar] [CrossRef]
- Petrini, O.; Stone, J.; Carroll, F.E. Endophytic fungi in evergreen shrubs in western Oregon: A preliminary study. Can. J. Bot. 1982, 60, 789–796. [Google Scholar] [CrossRef]
- Arnold, A.E.; Herre, E.A. Canopy cover and leaf age affect colonization by tropical fungal endophytes: Ecological pattern and process in Theobroma cacao (Malvaceae). Mycologia 2003, 95, 388–398. [Google Scholar] [CrossRef]
- Menkis, A.; Marčiulynas, A.; Gedminas, A.; Lynikienė, J.; Povilaitienė, A. High-throughput sequencing reveals drastic changes in fungal communities in the phyllosphere of Norway spruce (Picea abies) following invasion of the spruce bud scale (Physokermes piceae). Microb. Ecol. 2015, 70, 904–911. [Google Scholar] [CrossRef] [Green Version]
- Leray, M.; Knowlton, N.; Ho, S.L.; Nguyen, B.N.; Machida, R.J. GenBank is a reliable resource for 21st century biodiversity research. Proc. Natl. Acad. Sci. USA 2019, 116, 22651–22656. [Google Scholar] [CrossRef] [Green Version]
- Neuhauser, S.; Huber, L.; Kirchmair, M. Is Roesleria subterranea a primary pathogen or a minor parasite of grapevines? Risk assessment and a diagnostic decision scheme. Eur. J. Plant Pathol. 2011, 130, 503–510. [Google Scholar] [CrossRef] [Green Version]
- Kowalik, M.; Muras, P. Fungi Occurring on the Fallen Leaves of Rhododendron; Roczniki Akademii Rolniczej w Poznaniu: Ogrodnictwo, Poland, 2007. [Google Scholar]
- Slinkina, N.N.; Pivkin, M.V.; Polokhin, O.V. Filamentous fungi of the submarine soils of the Sakhalin Gulf (Sea of Okhotsk). Russ. J. Mar. Biol. 2010, 36, 413–418. [Google Scholar] [CrossRef]
- Nicoletti, R.; de Stefano, M. Penicillium restrictum as an antagonist of plant pathogenic fungi. Dyn. Biochem. Process. Biotechnol. Mol. Biol. 2012, 6, 61–69. [Google Scholar]
- Andrews, J.H. Population Growth and the Landscape Ecology. Microbial Ecology of Aerial Plant Surfaces; CABI: Wallingford, UK, 2006; pp. 239–250. [Google Scholar]
- Gramisci, B.R.; Lutz, M.C.; Lopes, C.A.; Sangorrín, M.P. Enhancing the efficacy of yeast biocontrol agents against postharvest pathogens through nutrient profiling and the use of other additives. Biol. Cont. 2018, 121, 151–158. [Google Scholar] [CrossRef]
- Pawlikowska, E.; James, S.A.; Breierova, E.; Antolak, H.; Kregiel, D. Biocontrol capability of local Metschnikowia sp. isolates. Antonie Van Leeuwenhoek 2019, 112, 1425–1445. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site * | Vegetation Type a | Forest Site Type b | Stand Age | Tree Species Composition in Decreasing Order of Occurrence | Location |
|---|---|---|---|---|---|
| G1 | Šcs | ox | 90 | Pinus sylvestris, Picea abies, Quercus robur | 54.872557° N, 023.97829° E |
| G2 | Šcl | ox | 40 | Alnus incana, Populus tremula, Acer platanoides, Tilia cordata, Ulmus glabra | 54.923794° N, 023.762145° E |
| G3 | Lcl | mox | 50 | Alnus incana, Alnus glutinosa, Acer platanoides, Ulmus glabra, Betula pendula | 55.010736° N, 024.529356° E |
| G4 | Ncl | ox | 50 | Alnus incana, Ulmus glabra, Tilia cordata, Populus tremula, Pinus sylvestris | 54.598671° N, 025.23843° E |
| G5 | Šds | hox | 100 | Quercus robur, Picea abies, Tilia cordata, Fraxinus excelsior, Ulmus glabra | 54.938315° N, 023.760325° E |
| G6 | Šdp | hox | 50 | Alnus incana, Betula pendula, Fraxinus excelsior, Ulmus glabra, Acer platanoides | 55.27918° N, 024.950571° E |
| Site No. | Tree Status * | Roots | Leaves | ||||
|---|---|---|---|---|---|---|---|
| Sequences | Taxa | Shannon Index (H) | Sequences | Taxa | Shannon Index (H) | ||
| G1 | H | 3962 | 77 | 1.82 | 652 | 76 | 3.07 |
| D | - | - | - | - | - | - | |
| G2 | H | 454 | 64 | 2.81 | 577 | 61 | 2.83 |
| D | 7443 | 46 | 0.95 | - | - | - | |
| G3 | H | 783 | 37 | 0.94 | 3444 | 140 | 3.06 |
| D | 3853 | 54 | 0.32 | 1105 | 99 | 3.2 | |
| G4 | H | 270 | 40 | 2.9 | 2730 | 142 | 3.1 |
| D | 381 | 70 | 3.42 | 873 | 100 | 3.09 | |
| G5 | H | 342 | 41 | 2.54 | 210 | 46 | 2.93 |
| D | - | - | - | - | - | - | |
| G6 | H | 161 | 25 | 1.09 | 4059 | 146 | 2.89 |
| D | 305 | 60 | 3.1 | 1095 | 101 | 3.1 | |
| TOTAL | H | 5972 | 182 | 2.81 | 11,672 | 275 | 3.34 |
| D | 11,982 | 156 | 1.66 | 3073 | 191 | 3.46 | |
| ALL | 17,954 | 254 | 2.58 | 14,745 | 325 | 3.41 |
| Phylum * | Species Name | Reference Number | Similarity, % | G1 | G2 | G3 | G4 | G5 | G6 | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H, % | D, % | H, % | D, % | H, % | D, % | H, % | D, % | H, % | D, % | H, % | D, % | H, % | D, % | All, % | ||||
| A | Trichocladium griseum | MN643061 | 100 | 0.03 | - | 12.56 | 78.34 | 0.64 | 0.08 | - | 0.79 | 0.29 | - | 0.62 | 1.31 | 1.09 | 48.75 | 32.90 |
| A | Penicillium restrictum | MT090009 | 100 | 0.05 | - | 1.10 | 0.59 | 2.30 | 95.80 | 3.33 | 4.46 | 4.09 | - | 0.62 | 1.97 | 0.82 | 31.36 | 21.20 |
| B | Unidentified sp. 5238_7 | KC588795 | 99 | 57.09 | - | - | - | - | - | - | - | - | - | - | - | 37.88 | - | 12.60 |
| A | Unidentified sp. 5238_20 | KX193283 | 100 | - | - | - | 11.00 | - | - | - | - | - | - | - | - | - | 6.84 | 4.56 |
| A | Roesleria subterranea | KU666540 | 100 | - | - | - | - | 84.04 | 0.10 | 9.63 | - | - | - | - | - | 11.45 | 0.03 | 3.83 |
| B | Cyclocybe sp. 5238_22 | MW644549 | 97 | 13.55 | - | - | - | - | - | - | - | - | - | 0.62 | 1.64 | 9.01 | 0.04 | 3.02 |
| A | Unidentified sp. 5238_39 | JF519577 | 100 | 5.78 | - | 13.88 | 0.03 | 0.13 | 0.21 | - | 0.79 | - | - | - | 14.10 | 4.91 | 0.47 | 1.94 |
| B | Trechisporales sp. 5238_32 | KU973892 | 100 | 1.59 | - | 27.31 | - | 0.26 | - | - | 9.97 | - | - | 0.62 | 22.95 | 3.18 | 0.90 | 1.66 |
| A | Leptodontidium camptobactrum | MH857172 | 100 | 0.03 | - | - | 2.42 | - | - | - | - | - | - | - | - | 0.02 | 1.50 | 1.01 |
| B | Mycena leptocephala | MT644911 | 100 | 0.03 | - | - | - | - | 0.31 | - | 0.79 | - | - | 80.12 | - | 2.18 | 0.13 | 0.81 |
| A | Unidentified sp. 5238_78 | MH636731 | 99 | 3.61 | - | - | - | - | - | - | - | - | - | - | - | 2.39 | - | 0.80 |
| B | Malassezia restricta | MK336446 | 99 | 0.20 | - | 3.96 | 0.01 | 0.64 | 0.21 | 8.15 | 15.49 | 0.88 | - | - | 4.92 | 0.94 | 0.69 | 0.77 |
| B | Xerocomellus porosporus | HM190086 | 99 | 2.98 | - | - | - | - | - | - | - | - | - | - | - | 1.98 | - | 0.66 |
| M | Podila minutissima | MT366014 | 100 | 0.15 | - | 0.22 | 1.42 | - | - | 0.37 | - | 0.29 | - | - | 0.33 | 0.15 | 0.89 | 0.65 |
| B | Thyrostroma tiliae | MK751738 | 100 | 2.62 | - | - | - | - | - | - | - | - | - | - | - | 1.74 | - | 0.58 |
| A | Dothideomycetes sp. 5238_97 | LR864332 | 99 | - | - | 6.83 | 0.01 | - | 0.44 | 2.96 | 8.66 | 1.46 | - | - | - | 0.74 | 0.43 | 0.53 |
| A | Camposporium multiseptatum | NR171863 | 100 | 2.32 | - | - | - | - | - | - | - | 0.29 | - | 0.62 | 0.33 | 1.57 | 0.01 | 0.53 |
| A | Trichoderma atroviride | MT514373 | 99 | 0.03 | - | - | 1.10 | - | - | - | - | 0.29 | - | - | 0.66 | 0.03 | 0.70 | 0.48 |
| B | Entoloma strigosissimum | JF908004 | 99 | - | - | 7.27 | - | - | 0.18 | 10.00 | 4.72 | - | - | - | - | 1.00 | 0.21 | 0.47 |
| B | Conocybe nigrescens | MK217423 | 100 | - | - | - | - | - | - | - | - | 22.51 | - | - | - | 1.29 | - | 0.43 |
| A | Cadophora orchidicola | MT436755 | 100 | 0.20 | - | - | 0.13 | 0.89 | 0.10 | - | 1.57 | 3.22 | - | 3.11 | 6.89 | 0.52 | 0.34 | 0.40 |
| A | Cladosporium sp. 5238_5 | MT645945 | 100 | 0.13 | - | 1.32 | 0.01 | 1.02 | - | 6.67 | 5.77 | 0.29 | - | - | 3.28 | 0.64 | 0.28 | 0.40 |
| B | Infundibulicybe geotropa | KT122792 | 100 | - | - | - | - | - | - | - | - | 20.76 | - | - | - | 1.19 | - | 0.40 |
| Z | Absidia glauca | EU484257 | 100 | - | - | - | 0.87 | - | - | - | - | - | - | - | - | - | 0.54 | 0.36 |
| A | Unidentified sp. 5238_132 | KX222675 | 100 | 1.34 | - | - | - | - | 0.03 | - | - | - | - | 0.62 | 2.30 | 0.90 | 0.07 | 0.35 |
| B | Unidentified sp. 5238_124 | LR874952 | 100 | - | - | - | - | - | 0.03 | - | - | 16.96 | - | - | - | 0.97 | 0.01 | 0.33 |
| B | Solicoccozyma terricola | MH487580 | 100 | 0.05 | - | - | 0.74 | 0.13 | - | - | - | - | - | - | - | 0.05 | 0.46 | 0.32 |
| A | Unidentified sp. 5238_145 | MK627256 | 99 | 1.26 | - | - | - | - | 0.05 | - | - | - | - | - | 1.64 | 0.84 | 0.06 | 0.32 |
| A | Pseudogymnoascus sp. 5238_46 | MT367251 | 100 | - | - | 0.22 | 0.63 | 0.13 | 0.05 | - | 0.26 | 0.29 | - | 0.62 | 0.66 | 0.07 | 0.43 | 0.31 |
| B | Solicoccozyma aeria | MT596205 | 100 | - | - | - | 0.70 | - | - | - | - | - | - | - | - | - | 0.43 | 0.29 |
| 93.03 | 0.00 | 74.67 | 98.02 | 90.17 | 97.59 | 41.11 | 53.28 | 71.64 | 0.00 | 87.58 | 62.95 | 87.54 | 95.57 | 92.90 | ||||
| Phylum * | Species Name | Reference Number | Similarity, % | G1 | G2 | G3 | G4 | G5 | G6 | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H, % | D, % | H, % | D, % | H, % | D, % | H, % | D, % | H, % | D, % | H, % | D, % | H, % | D, % | All, % | ||||
| A | Trichomerium sp. 5238_8 | MT223865 | 94 | 5.98 | - | 14.56 | - | 10.54 | 10.23 | 20.22 | 8.59 | - | - | 9.76 | 13.52 | 12.66 | 10.93 | 12.30 |
| A | Aureobasidium pullulans | MT645930 | 100 | 3.68 | - | 10.92 | - | 9.93 | 13.12 | 7.33 | 4.93 | 5.24 | - | 17.10 | 15.98 | 12.09 | 11.81 | 12.03 |
| A | Cladosporium sp. 5238_5 | MT645945 | 100 | 5.83 | - | 11.44 | - | 5.72 | 7.87 | 7.69 | 13.29 | 27.62 | - | 18.34 | 11.87 | 11.96 | 10.84 | 11.73 |
| B | Vishniacozyma carnescens | MT595884 | 100 | 15.03 | - | 11.79 | - | 10.31 | 7.69 | 5.64 | 9.62 | 13.33 | - | 10.31 | 10.68 | 10.01 | 9.31 | 9.86 |
| A | Unidentified sp. 5238_14 | KX147989 | 90 | - | - | - | - | 24.16 | 18.55 | 2.93 | 0.11 | - | - | - | - | 7.81 | 6.70 | 7.58 |
| A | Unidentified sp. 5238_9 | MT236420 | 100 | 2.76 | - | 0.69 | - | 2.64 | 0.54 | 8.75 | 9.51 | - | - | 3.46 | 6.39 | 4.35 | 5.17 | 4.52 |
| A | Coniozyma sp. 5238_17 | MW764574 | 100 | 8.13 | - | 0.35 | - | 3.02 | 1.54 | 13.08 | 1.37 | 0.48 | - | 0.51 | 4.20 | 4.63 | 2.44 | 4.17 |
| B | Unidentified sp. 5238_30 | MN903728 | 100 | 13.50 | - | 5.55 | - | 4.33 | 4.98 | 1.61 | 2.06 | 6.19 | - | 1.84 | 2.28 | 3.50 | 3.19 | 3.44 |
| A | Didymella sp. 5238_12 | MT453298 | 100 | 2.30 | - | 8.32 | - | 1.25 | 0.54 | 4.58 | 2.52 | 4.76 | - | 2.84 | 5.11 | 3.16 | 2.73 | 3.07 |
| B | Unidentified sp. 5238_33 | KU057810 | 92 | 3.37 | - | 0.52 | - | 0.87 | 1.81 | 1.06 | 2.29 | 1.90 | - | 4.70 | 5.30 | 2.57 | 3.19 | 2.70 |
| B | Buckleyzyma aurantiaca | KX096691 | 100 | 3.37 | - | 0.52 | - | 1.54 | 0.54 | 2.75 | 1.15 | 0.48 | - | 1.53 | 1.55 | 1.91 | 1.07 | 1.74 |
| A | Lemonniera sp. 5238_6 | KX096679 | 99 | 14.42 | - | 2.43 | - | 1.10 | 1.00 | 1.47 | 0.80 | 0.95 | - | 0.42 | 0.27 | 1.77 | 0.68 | 1.55 |
| A | Unidentified sp. 5238_53 | AB476498 | 97 | 1.38 | - | - | - | 1.22 | 1.63 | 0.66 | - | - | - | 2.64 | 1.10 | 1.61 | 0.98 | 1.48 |
| B | Filobasidium wieringae | MN128850 | 100 | 0.92 | - | 0.87 | - | 1.39 | 2.44 | 2.42 | 0.92 | 0.48 | - | 0.60 | 0.27 | 1.31 | 1.24 | 1.30 |
| A | Capnodium sp. 5238_75 | AJ244240 | 96 | - | - | - | - | 0.03 | - | 0.15 | 19.70 | - | - | - | - | 0.04 | 5.60 | 1.20 |
| A | Ramularia vizellae | MK012421 | 100 | 0.46 | - | 2.25 | - | 1.74 | 3.35 | 0.44 | 0.69 | 1.43 | - | 0.69 | 0.18 | 1.05 | 1.46 | 1.13 |
| B | Tremellales sp. 5238_52 | MG827438 | 100 | 0.61 | - | 0.52 | - | 0.44 | 0.72 | 1.98 | 1.03 | 2.38 | - | 0.49 | 0.73 | 0.88 | 0.81 | 0.87 |
| B | Rhodosporidiobolus colostri | MT502792 | 100 | 0.31 | - | 0.52 | - | 2.15 | 1.81 | 0.11 | - | - | - | 0.20 | 1.00 | 0.78 | 1.01 | 0.83 |
| B | Filobasidium dingjieense | MK050343 | 100 | - | - | 1.21 | - | 2.61 | 0.81 | - | - | - | - | 0.02 | 0.37 | 0.84 | 0.42 | 0.75 |
| B | Filobasidium mucilaginum | MK050349 | 99 | - | - | 0.17 | - | 0.12 | 0.27 | 1.06 | 1.72 | 0.48 | - | 0.86 | 0.64 | 0.63 | 0.81 | 0.67 |
| B | Bannozyma yamatoana | AF444634 | 99 | - | - | - | - | 0.61 | 1.63 | 0.70 | - | - | - | 0.73 | 0.37 | 0.63 | 0.72 | 0.64 |
| B | Sporobolomyces roseus | MT502791 | 100 | 0.15 | - | 0.35 | - | 0.06 | 0.36 | 0.15 | 0.11 | 4.76 | - | 1.29 | 1.10 | 0.66 | 0.55 | 0.64 |
| A | Unidentified sp. 5238_113 | KT328793 | 90 | 1.53 | - | 14.21 | - | - | - | - | - | - | - | 0.02 | 0.09 | 0.80 | 0.03 | 0.64 |
| B | Dioszegia sp. 5238_18 | LT548261 | 100 | 1.38 | - | 1.56 | - | 0.38 | 0.27 | 0.26 | 0.23 | 0.95 | - | 0.69 | 1.00 | 0.61 | 0.52 | 0.59 |
| B | Malassezia restricta | MK336446 | 99 | 0.15 | - | - | - | 0.17 | 0.54 | 0.22 | 3.09 | 4.76 | - | 0.07 | 2.19 | 0.22 | 1.85 | 0.56 |
| A | Neosetophoma sp. 5238_42 | MN244543 | 100 | 0.31 | - | 0.35 | - | 0.29 | 0.18 | 0.18 | 1.72 | 0.48 | - | 0.64 | 0.64 | 0.42 | 0.78 | 0.50 |
| B | Curvibasidium cygneicollum | KY102972 | 100 | - | - | 0.17 | - | 0.90 | 1.18 | 0.04 | - | - | - | 0.22 | 0.46 | 0.37 | 0.59 | 0.41 |
| A | Lapidomyces aloidendricola | NR173048 | 98 | - | - | - | - | 0.03 | - | 2.01 | - | - | - | - | - | 0.48 | - | 0.38 |
| B | Cryptococcus sp. 5238_131 | KT314197 | 99 | 0.15 | - | 1.04 | - | 0.32 | 0.09 | 0.22 | 0.80 | 0.95 | - | 0.40 | 0.37 | 0.38 | 0.39 | 0.38 |
| A | Chaetothyriales sp. 5238_142 | KX402745 | 99 | - | - | - | - | 0.75 | 1.09 | 0.18 | 0.92 | - | - | - | - | 0.27 | 0.65 | 0.35 |
| All | 85.74 | 0.00 | 90.29 | 0.00 | 88.62 | 84.80 | 87.88 | 87.17 | 77.62 | 0.00 | 80.37 | 87.67 | 88.40 | 86.50 | 88.00 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marčiulynas, A.; Marčiulynienė, D.; Lynikienė, J.; Bakys, R.; Menkis, A. Fungal Communities in Leaves and Roots of Healthy-Looking and Diseased Ulmus glabra. Microorganisms 2022, 10, 2228. https://doi.org/10.3390/microorganisms10112228
Marčiulynas A, Marčiulynienė D, Lynikienė J, Bakys R, Menkis A. Fungal Communities in Leaves and Roots of Healthy-Looking and Diseased Ulmus glabra. Microorganisms. 2022; 10(11):2228. https://doi.org/10.3390/microorganisms10112228
Chicago/Turabian StyleMarčiulynas, Adas, Diana Marčiulynienė, Jūratė Lynikienė, Remigijus Bakys, and Audrius Menkis. 2022. "Fungal Communities in Leaves and Roots of Healthy-Looking and Diseased Ulmus glabra" Microorganisms 10, no. 11: 2228. https://doi.org/10.3390/microorganisms10112228
APA StyleMarčiulynas, A., Marčiulynienė, D., Lynikienė, J., Bakys, R., & Menkis, A. (2022). Fungal Communities in Leaves and Roots of Healthy-Looking and Diseased Ulmus glabra. Microorganisms, 10(11), 2228. https://doi.org/10.3390/microorganisms10112228