
Comparison of the Peel-Associated Epiphytic Bacteria of Anthocyanin-Rich “Sun Black” and Wild-Type Tomatoes under Organic and Conventional Farming

 ,
,  ,
,  ,
, 
 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Sample Collection and Processing
2.3. DNA Extraction, 16S rRNA Gene Amplicon Libraries and Sequencing
2.4. Sequence Processing and Data Analysis
2.5. Statistical Methods
3. Results
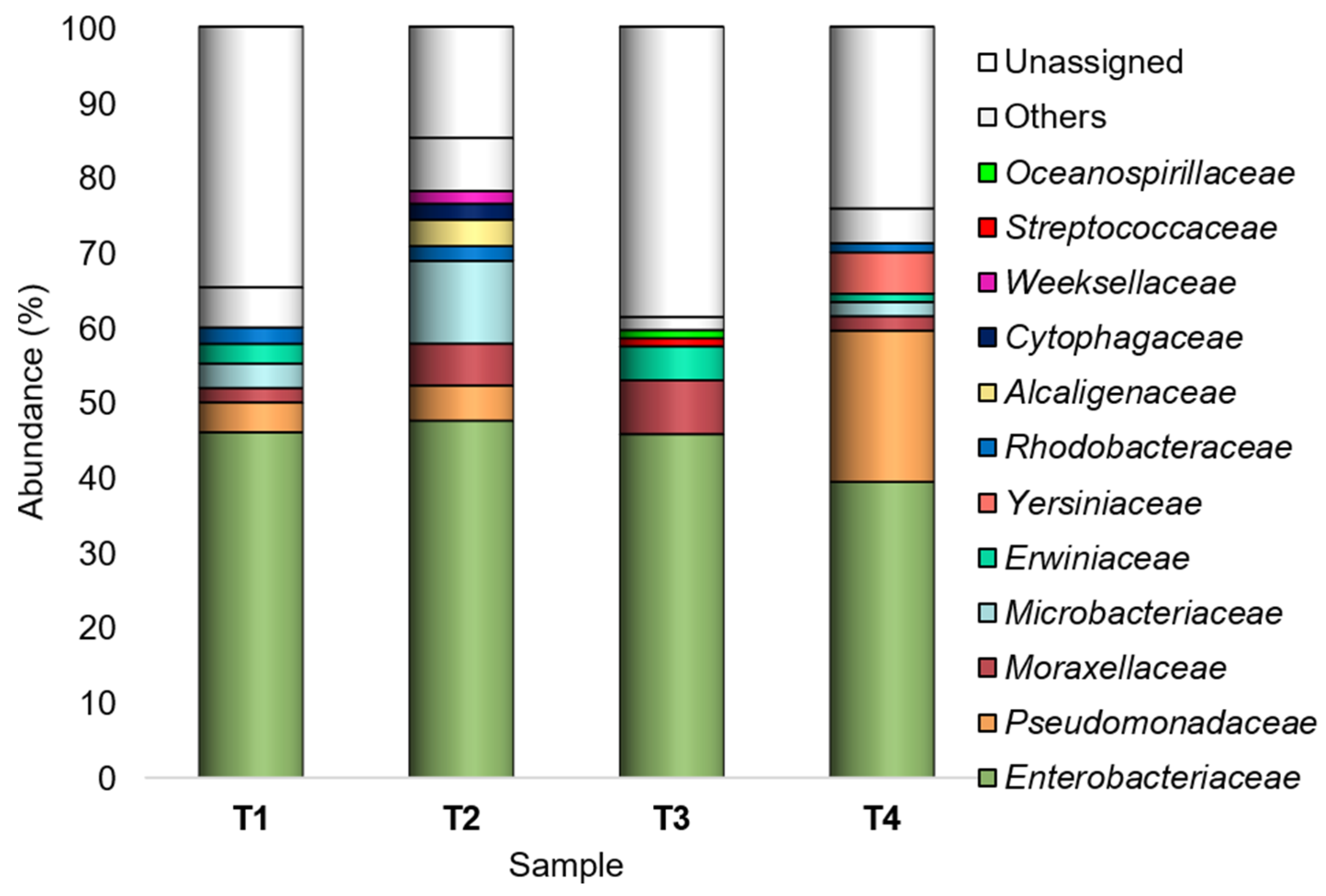
3.1. Composition of the Epiphytic Peel-Associated Bacteria
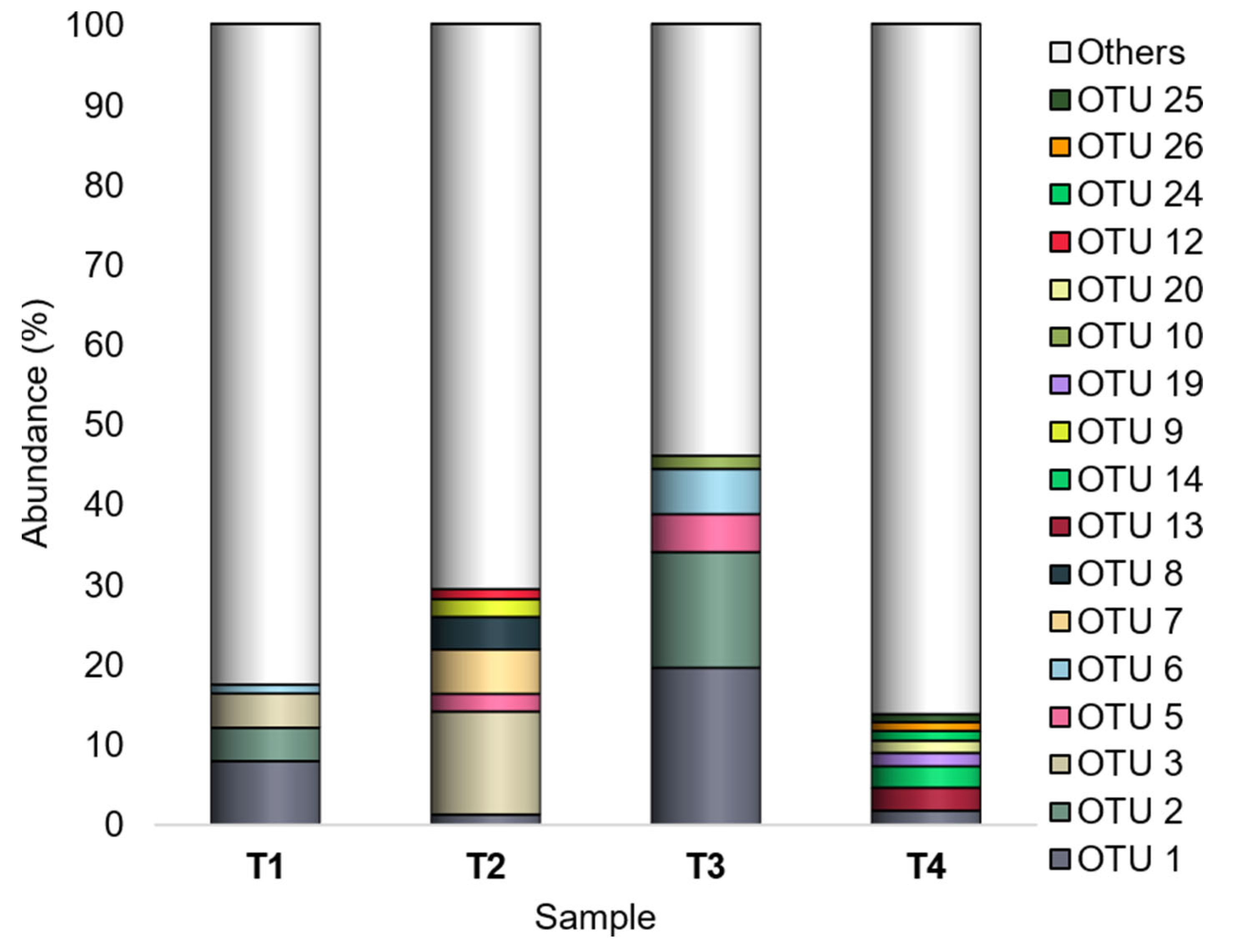
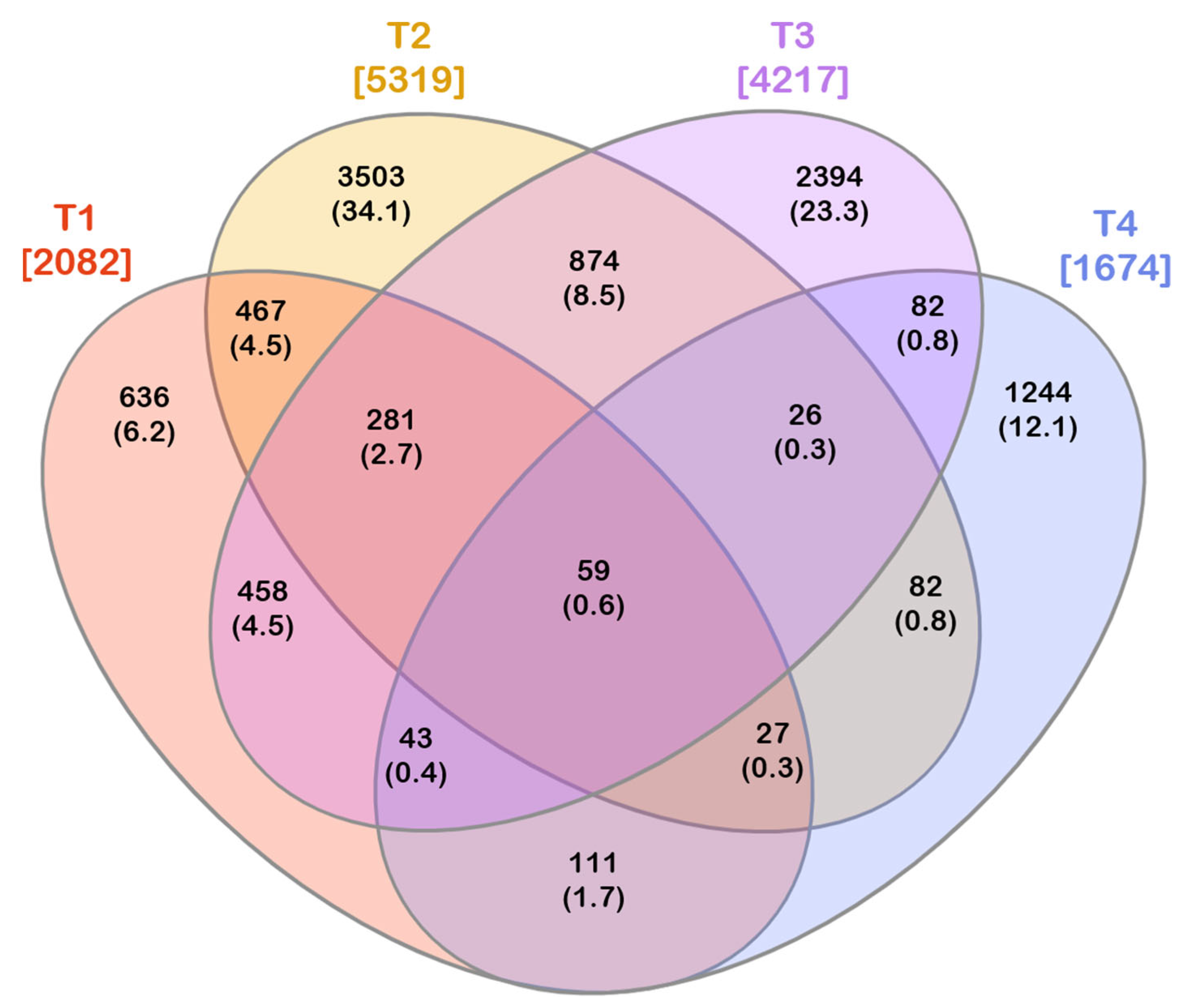
3.2. Unique and Shared Bacterial OTUs
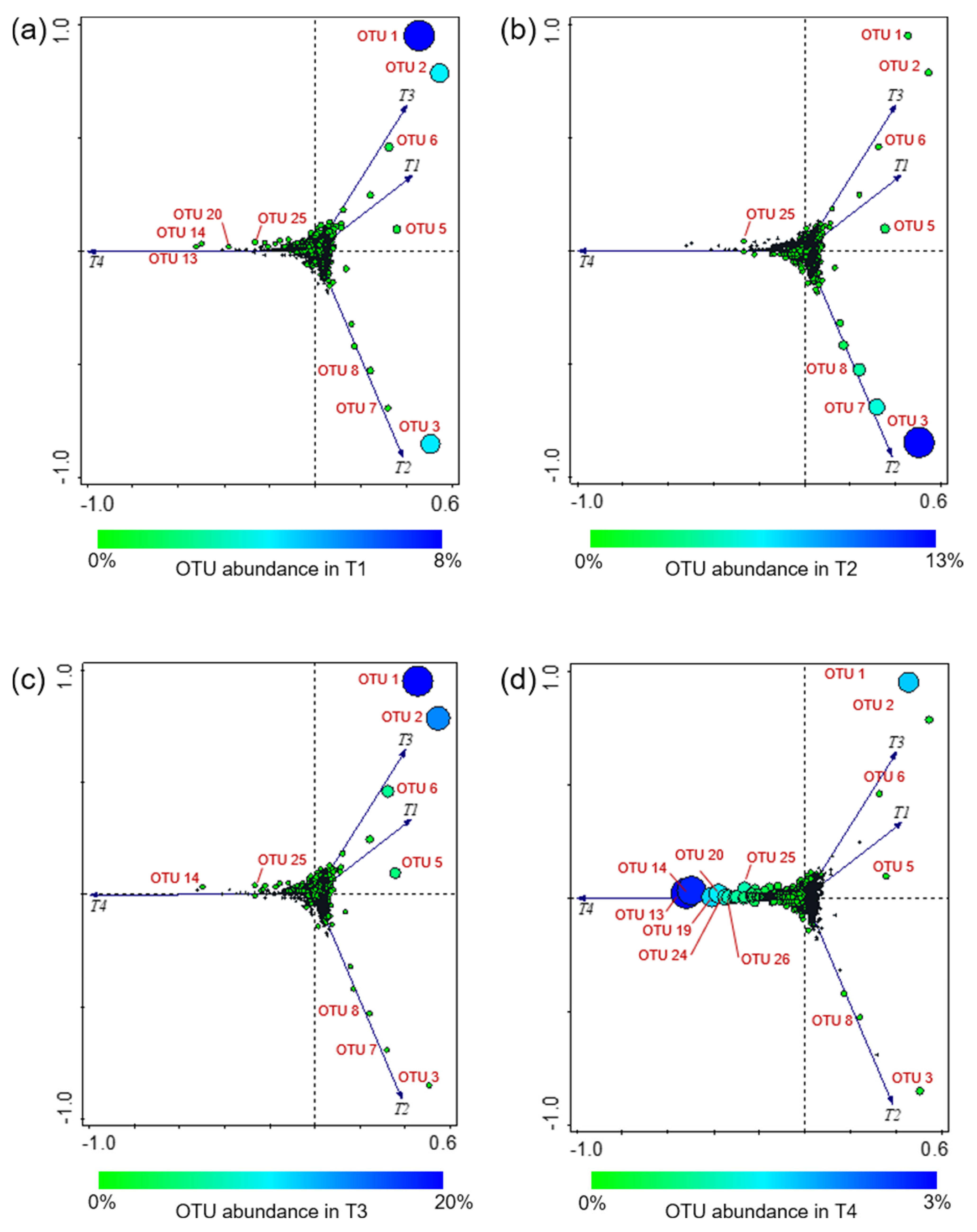
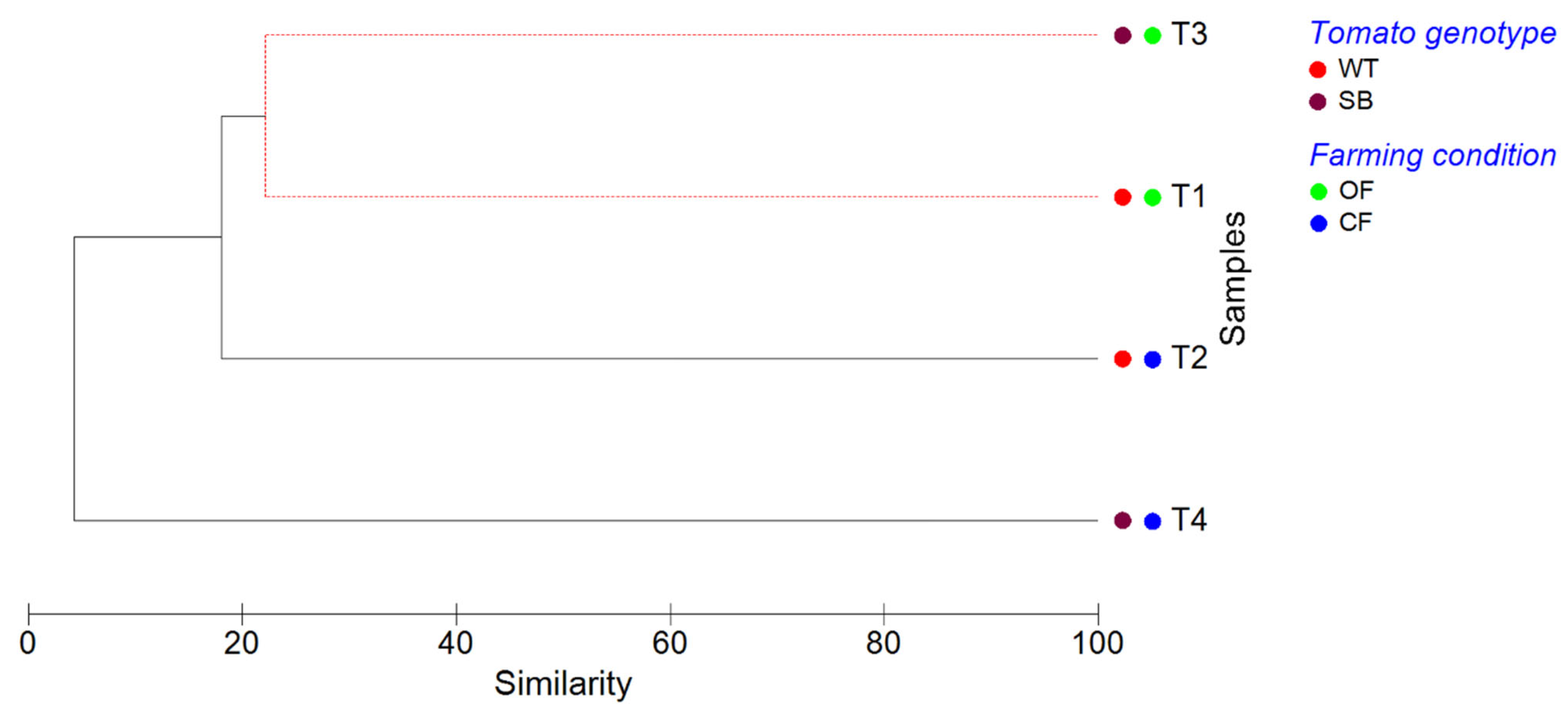
3.3. Alpha- and Beta-Diversity Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT, Crops and Livestock Products, Tomatoes. Available online: http://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 1 October 2022).
- Kandel, D.R.; Marconi, T.G.; Badillo-Vargas, I.E.; Enciso, J.; Zapata, S.D.; Lazcano, C.A.; Crosby, K.; Avila, C.A. Yield and fruit quality of high-tunnel tomato cultivars produced during the off-season in South Texas. Sci. Hortic. 2020, 272, 109582. [Google Scholar] [CrossRef]
- Borghesi, E.; González-Miret, M.L.; Escudero-Gilete, M.L.; Malorgio, F.; Heredia, F.J.; Meléndez-Martínez, A.J. Effects of salinity stress on carotenoids, anthocyanins, and color of diverse tomato genotypes. J. Agric. Food Chem. 2011, 59, 11676–11682. [Google Scholar] [CrossRef] [PubMed]
- Caseiro, M.; Ascenso, A.; Costa, A.; Creagh-Flynn, J.; Johnson, M.; Simões, S. Lycopene in human health. LWT 2020, 127, 109323. [Google Scholar] [CrossRef]
- Ronga, D.; Gallingani, T.; Zaccardelli, M.; Perrone, D.; Francia, E.; Milc, J.; Pecchioni, N. Carbon footprint and energetic analysis of tomato production in the organic vs. the conventional cropping systems in Southern Italy. J. Clean. Prod. 2019, 220, 836–845. [Google Scholar] [CrossRef] [Green Version]
- Natalini, A.; Acciarri, N.; Cardi, T. Breeding for nutritional and organoleptic quality in vegetable crops: The case of tomato and cauliflower. Agriculture 2021, 11, 606. [Google Scholar] [CrossRef]
- Roberts, D.P.; Mattoo, A.K. Sustainable crop production systems and human nutrition. Front. Sustain. Food Syst. 2019, 3, 72. [Google Scholar] [CrossRef]
- Bona, E.; Cantamessa, S.; Massa, N.; Manassero, P.; Marsano, F.; Copetta, A.; Lingua, G.; D’Agostino, G.; Gamalero, E.; Berta, G. Arbuscular mycorrhizal fungi and plant growth-promoting pseudomonads improve yield, quality and nutritional value of tomato: A field study. Mycorrhiza 2017, 27, 1–11. [Google Scholar] [CrossRef]
- Bona, E.; Todeschini, V.; Cantamessa, S.; Cesaro, P.; Copetta, A.; Lingua, G.; Gamalero, E.; Berta, G.; Massa, N. Combined bacterial and mycorrhizal inocula improve tomato quality at reduced fertilization. Sci. Hortic. 2018, 234, 160–165. [Google Scholar] [CrossRef]
- Roberts, D.; Mattoo, A. Sustainable agriculture—Enhancing environmental benefits, food nutritional quality and building crop resilience to abiotic and biotic stresses. Agriculture 2018, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Gonzali, S.; Perata, P. Anthocyanins from Purple Tomatoes as Novel Antioxidants to Promote Human Health. Antioxidants 2020, 9, 1017. [Google Scholar] [CrossRef]
- Luo, D.; Xiong, C.; Lin, A.; Zhang, C.; Sun, W.; Zhang, J.; Yang, C.; Lu, Y.; Li, H.; Ye, Z.; et al. SlBBX20 interacts with the COP9 signalosome subunit SlCSN5-2 to regulate anthocyanin biosynthesis by activating SlDFR expression in tomato. Hortic. Res. 2021, 8, 163. [Google Scholar] [CrossRef]
- Blando, F.; Berland, H.; Maiorano, G.; Durante, M.; Mazzucato, A.; Picarella, M.E.; Nicoletti, I.; Gerardi, C.; Mita, G.; Andersen, Ø.M. Nutraceutical Characterization of Anthocyanin-Rich Fruits Produced by “Sun Black” Tomato Line. Front. Nutr. 2019, 6, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Butelli, E.; De Stefano, R.; Schoonbeek, H.J.; Magusin, A.; Pagliarani, C.; Wellner, N.; Hill, L.; Orzaez, D.; Granell, A.; et al. Anthocyanins double the shelf life of tomatoes by delaying overripening and reducing susceptibility to gray mold. Curr. Biol. 2013, 23, 1094–1100. [Google Scholar] [CrossRef] [Green Version]
- Mazzucato, A.; Willems, D.; Bernini, R.; Picarella, M.E.; Santangelo, E.; Ruiu, F.; Tilesi, F.; Soressi, G.P. Novel phenotypes related to the breeding of purple-fruited tomatoes and effect of peel extracts on human cancer cell proliferation. Plant Phys. Biochem. 2013, 72, 125–133. [Google Scholar] [CrossRef]
- Tilesi, F.; Lombardi, A.; Mazzucato, A. Scientometric and Methodological Analysis of the Recent Literature on the Health-Related Effects of Tomato and Tomato Products. Foods 2021, 10, 1905. [Google Scholar] [CrossRef]
- Gupta, R.; Patil, R. Phyllospheric microbes: Diversity, functions, interaction, and applications in agriculture. In Current Trends in Microbial Biotechnology for Sustainable Agriculture; Yadav, A.N., Singh, J., Singh, C., Yadav, N., Eds.; Springer: Singapore, 2021; pp. 301–323. [Google Scholar] [CrossRef]
- Mamphogoro, T.P.; Babalola, O.O.; Aiyegoro, O.A. Exploitation of epiphytic bacterial antagonists for the management of post-harvest diseases of sweet pepper and other fresh produce—A viable option. Biocontrol Sci. Technol. 2020, 30, 741–761. [Google Scholar] [CrossRef]
- Berger, C.N.; Sodha, S.V.; Shaw, R.K.; Griffin, P.M.; Pink, D.; Hand, P.; Frankel, G. Fresh fruit and vegetables as vehicles for the transmission of human pathogens. Environ. Microbiol. 2010, 12, 2385–2397. [Google Scholar] [CrossRef]
- de São José, J.F.B.; Faria-Silva, L.; Lepaus, B.M. Decontamination of Vegetables. In Microbial Decontamination of Food; Shah, M.A., Mir, S.A., Eds.; Springer: Singapore, 2022; pp. 71–92. [Google Scholar] [CrossRef]
- Vandenkoornhuyse, P.; Quaiser, A.; Le Van Duhamel, M.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Abadi, V.A.J.M.; Sepehri, M.; Rahmani, H.A.; Zarei, M.; Ronaghi, A.; Taghavi, S.M.; Shamshiripour, M. Role of dominant phyllosphere bacteria with plant growth–promoting characteristics on growth and nutrition of maize (Zea mays L.). J. Soil Sci. Plant Nutr. 2020, 20, 2348–2363. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Plant endophytes and epiphytes: Burgeoning sources of known and “unknown” cytotoxic and antibiotic agents? Planta Med. 2020, 86, 891–905. [Google Scholar] [CrossRef] [Green Version]
- Al-Maawali, S.S.; Al-Sadi, A.M.; Alsheriqi, S.A.K.; Al-Sabahi, J.N.; Velazhahan, R. The potential of antagonistic yeasts and bacteria from tomato phyllosphere and fructoplane in the control of Alternaria fruit rot of tomato. All Life 2021, 14, 34–48. [Google Scholar] [CrossRef]
- Hammami, R.; Oueslati, M.; Smiri, M.; Nefzi, S.; Ruissi, M.; Comitini, F.; Romanazzi, G.; Cacciola, S.O.; Sadfi Zouaoui, N. Epiphytic Yeasts and Bacteria as Candidate Biocontrol Agents of Green and Blue Molds of Citrus Fruits. J. Fungi 2022, 8, 818. [Google Scholar] [CrossRef] [PubMed]
- Campiglia, E.; Mancinelli, R.; Radicetti, E. Influence of no-tillage and organic mulching on tomato (Solanum lycopersicum L.) production and nitrogen use in the mediterranean environment of central Italy. Sci. Hortic. 2011, 130, 588–598. [Google Scholar] [CrossRef]
- Campiglia, E.; Radicetti, E.; Mancinelli, R. Cover crops and mulches influence weed management and weed flora composition in strip-tilled tomato (Solanum lycopersicum). Weed Res. 2015, 55, 416–425. [Google Scholar] [CrossRef] [Green Version]
- Ottesen, A.R.; Peña, A.G.; White, J.R.; Pettengill, J.B.; Li, C.; Allard, S.; Rideout, S.; Allard, M.; Hill, T.; Evans, P.; et al. Baseline survey of the anatomical microbial ecology of an important food plant: Solanum lycopersicum (tomato). BMC Microbiol. 2013, 13, 114. [Google Scholar] [CrossRef] [Green Version]
- Janisiewicz, W.J.; Ii, W.M.J.; Peter, K.A.; Kurtzman, C.P.; Buyer, J. Yeasts associated with plums and their potential for controlling brown rot after harvest. Yeast 2014, 31, 207–218. [Google Scholar] [CrossRef]
- Fernández-Suárez, R.; Ramírez-Villatoro, G.; Díaz-Ruiz, G.; Eslava, C.; Calderón, M.; Navarro-Ocaña, A.; Trejo-Márquez, A.; Wacher, C. Effect of postharvest UV-C treatment on the bacterial diversity of Ataulfo mangoes by PCR-DGGE, survival of E. coli and antimicrobial activity. Front. Microbiol. 2013, 4, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, J.A.; Welch, D.B.M.; Morrison, H.G.; Huse, S.M.; Neal, P.R.; Butterfield, D.A.; Sogin, M.L. Microbial Population Structures in the Deep Marine Biosphere. Science 2007, 318, 97–100. [Google Scholar] [CrossRef]
- Wang, Y.; Qian, P.Y. Conservative Fragments in Bacterial 16S rRNA Genes and Primer Design for 16S Ribosomal DNA Amplicons in Metagenomic Studies. PLoS ONE 2009, 4, e7401. [Google Scholar] [CrossRef] [Green Version]
- Gorrasi, S.; Franzetti, A.; Ambrosini, R.; Pittino, F.; Pasqualetti, M.; Fenice, M. Spatio-temporal variation of the bacterial communities along a salinity gradient within a thalassohaline environment (Saline di Tarquinia salterns, Italy). Molecules 2021, 26, 1338. [Google Scholar] [CrossRef]
- Gorrasi, S.; Pasqualetti, M.; Braconcini, M.; Muñoz-Palazon, B.; Fenice, M. Could Pontimonas Harbour Halophilic Members Able to Withstand Very Broad Salinity Variations? Microorganisms 2022, 10, 790. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Palazon, B.; Rosa-Masegosa, A.; Vilchez-Vargas, R.; Link, A.; Gorrasi, S.; Gonzalez-Lopez, J.; Gonzalez-Martinez, A. Biological Removal Processes in Aerobic Granular Sludge for Treating Synthetic Hospital Wastewater: Effect of Temperature. J. Water Process Eng. 2022, 47, 102691. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claesson, M.J.; O’Sullivan, O.; Wang, Q.; Nikkilä, J.; Marchesi, J.R.; Smidt, H.; de Vos, W.M.; Ross, R.P.; O’Toole, P.W. Comparative analysis of pyrosequencing and a phylogenetic microarray for exploring microbial community structures in the human distal intestine. PLoS ONE 2009, 4, e6669. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E. A Mathematical Theory of Communication. Bell. Sys. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- Solow, A.R. A simple test for change in community structure. J. Anim. Ecol. 1993, 2, 191–193. [Google Scholar] [CrossRef]
- Seaby, R.M.; Henderson, P.A. Species Diversity and Richness, Version 4; Pisces Conservation Ltd.: Lymington, UK, 2006. [Google Scholar]
- Clarke, K.R.; Somerfield, P.J.; Gorley, R.N. Testing of null hypotheses in exploratory community analyses: Similarity profiles and biota-environment linkage. J. Exp. Mar. Biol. Ecol. 2008, 366, 56–69. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [Green Version]
- Leff, J.W.; Fierer, N. Bacterial Communities Associated with the Surfaces of Fresh Fruits and Vegetables. PLoS ONE 2013, 8, e59310. [Google Scholar] [CrossRef] [Green Version]
- Constancias, F.; Saby, N.P.A.; Terrat, S.; Dequiedt, S.; Horrigue, W.; Nowak, V.; Guillemin, J.-P.; Biju-Duval, L.; Prévost-Bouré, N.C.; Ranjard, L. Contrasting Spatial Patterns and Ecological Attributes of Soil Bacterial and Archaeal Taxa across a Landscape. MicrobiologyOpen 2015, 4, 518–531. [Google Scholar] [CrossRef]
- Dong, C.-J.; Wang, L.-L.; Li, Q.; Shang, Q.-M. Bacterial communities in the rhizosphere, phyllosphere and endosphere of tomato plants. PLoS ONE 2019, 14, e022384. [Google Scholar] [CrossRef] [PubMed]
- Bill, M.; Chidamba, L.; Gokul, J.K.; Korsten, L. Mango endophyte and epiphyte microbiome composition during fruit development and post-harvest stages. Horticulturae 2021, 7, 495. [Google Scholar] [CrossRef]
- Kumar, A.; Zhimo, Y.; Biasi, A.; Salim, S.; Feygenberg, O.; Wisniewski, M.; Droby, S. Endophytic Microbiome in the Carposphere and Its Importance in Fruit Physiology and Pathology. In Postharvest Pathology: Plant Pathology in the 21st Century; Spadaro, D., Droby, S., Gullino, M.L., Eds.; Springer: Cham, Switzerland, 2021; Volume 11, pp. 73–88. [Google Scholar] [CrossRef]
- Berg, G.; Erlacher, A.; Grube, M. The Edible Plant Microbiome: Importance and Health Issues. In Principles of Plant-Microbe Interactions: Microbes for Sustainable Agriculture; Lugtenberg, B., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 419–426. [Google Scholar] [CrossRef]
- Ottesen, A.; Ramachandran, P.; Reed, E.; Gu, G.; Gorham, S.; Ducharme, D.; Newell, M.; Rideout, S.; Turini, T.; Hill, T.; et al. Metagenome tracking biogeographic agroecology: Phytobiota of tomatoes from virginia, maryland, north carolina and california. Food Microbiol. 2019, 79, 132–136. [Google Scholar] [CrossRef]
- Kämpfer, P. Buttiauxella. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.E.B., Ed.; John Wiley & Sons: New York, NY, USA, 2015; pp. 1–16. [Google Scholar] [CrossRef]
- Gorrasi, S.; Pasqualetti, M.; Franzetti, A.; Gonzalez-Martinez, A.; Gonzalez-Lopez, J.; Muñoz-Palazon, B.; Fenice, M. Persistence of Enterobacteriaceae drawn into a marine saltern (Saline di Tarquinia, Italy) from the adjacent coastal zone. Water 2021, 13, 1443. [Google Scholar] [CrossRef]
- Wu, K.; Luo, J.; Li, J.; An, Q.; Yang, X.; Liang, Y.; Li, T. Endophytic bacterium Buttiauxella sp. SaSR13 improves plant growth and cadmium accumulation of hyperaccumulator Sedum alfredii. Environ. Sci. Pollut. Res. 2018, 25, 21844–21854. [Google Scholar] [CrossRef]
- Almasia, R.; Henríquez, M.; Levican, A.; Poblete-Morales, M. Genome sequence of a potentially new Buttiauxella species, Strain B2, isolated from rhizosphere of olivillo trees (Aextoxicon punctatum). Microbiol. Resour. Announc. 2020, 9, e01351-19. [Google Scholar] [CrossRef] [Green Version]
- Xin, Y.-Y.; Rahman, A.; Li, H.-X.; Xu, T.; Ding, G.-C.; Li, J. Modification of total and phosphorus mineralizing bacterial communities associated with Zea mays L. through plant development and fertilization regimes. J. Integr. Agric. 2021, 20, 3026–3038. [Google Scholar] [CrossRef]
- Sahu, K.P.; Patel, A.; Kumar, M.; Sheoran, N.; Mehta, S.; Reddy, B.; Eke, P.; Prabhakaran, N.; Kumar, A. Integrated metabarcoding and culturomic-based microbiome profiling of rice phyllosphere reveal diverse and functional bacterial communities for blast disease suppression. Front. Microbiol. 2021, 12, 780458. [Google Scholar] [CrossRef] [PubMed]
- Ottesen, A.R.; Gorham, S.; Pettengill, J.B.; Rideout, S.; Evans, P.; Brown, E. The impact of systemic and copper pesticide applications on the phyllosphere microflora of tomatoes. J. Sci. Food Agric. 2015, 95, 1116–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechan Llontop, M.E.; Tian, L.; Sharma, P.; Heflin, L.; Bernal-Galeano, V.; Haak, D.C.; Clarke, C.R.; Vinatzer, B.A. Experimental evidence pointing to rain as a reservoir of tomato phyllosphere microbiota. Phytobiomes J. 2021, 5, 382–399. [Google Scholar] [CrossRef]
- Hata, H.; Natori, T.; Mizuno, T.; Kanazawa, I.; Eldesouky, I.; Hayashi, M.; Miyata, M.; Fukunaga, H.; Ohji, S.; Hosoyama, A.; et al. Phylogenetics of family Enterobacteriaceae and proposal to reclassify Escherichia hermannii and Salmonella subterranea as Atlantibacter hermannii and Atlantibacter subterranea gen. Nov., Comb. Nov. Microbiol. Immunol. 2016, 60, 303–311. [Google Scholar] [CrossRef]
- Janda, J.M.; Abbott, S.L. The Changing Face of the Family Enterobacteriaceae (Order: “Enterobacterales”): New Members, Taxonomic Issues, Geographic Expansion, and New Diseases and Disease Syndromes. Clin. Microbiol. Rev. 2021, 34, e00174-20. [Google Scholar] [CrossRef]
- Shelobolina, E.S.; Sullivan, S.A.; O’Neill, K.R.; Nevin, K.P.; Lovley, D.R. Isolation, characterization, and U (VI)-reducing potential of a facultatively anaerobic, acid-resistant bacterium from low-pH, nitrate- and U (VI)-contaminated subsurface sediment and description of Salmonella subterranea sp. nov. Appl. Environ. Microbiol. 2004, 70, 2959–2965. [Google Scholar] [CrossRef] [Green Version]
- Watanapokasin, R.Y.; Boonyakamol, A.; Sukseree, S.; Krajarng, A.; Sophonnithiprasert, T.; Kanso, S.; Imai, T. Hydrogen production and anaerobic decolorization of wastewater containing Reactive Blue 4 by a bacterial consortium of Salmonella subterranea and Paenibacillus polymyxa. Biodegradation 2009, 20, 411–418. [Google Scholar] [CrossRef]
- Higdon, S.M.; Pozzo, T.; Kong, N.; Huang, B.C.; Yang, M.L.; Jeannotte, R.; Brown, C.T.; Bennett, A.B.; Weimer, B.C. Genomic Characterization of a Diazotrophic Microbiota Associated with Maize Aerial Root Mucilage. PLoS ONE 2020, 15, e0239677. [Google Scholar] [CrossRef]
- Besaury, L.; Rémond, C. Culturable and metagenomic approaches of wheat bran and wheat straw phyllosphere’s highlight new lignocellulolytic microorganisms. Lett. Appl. Microbiol. 2022, 74, 840–850. [Google Scholar] [CrossRef]
- Hu, H.; Liu, Y.; Huang, Y.; Zhang, Z.; Tang, H. The Leaf Microbiome of Tobacco Plants across Eight Chinese Provinces. Microorganisms 2022, 10, 450. [Google Scholar] [CrossRef]
- PLaBAse-PLant-Associated BActeria Web Resource, Atlantibacter. Available online: https://plabase.informatik.uni-tuebingen.de/pb/plaba_db_q.php (accessed on 1 October 2022).
- Pati, B.R.; Chandra, A.K. Diazotrophic bacterial population and other associated organisms on the phyllosphere of some crop plants. Zentralbl. Mikrobiol. 1993, 148, 392–402. [Google Scholar] [CrossRef]
- Pande, B.N.; Krysinska-Traczyk, E.; Prazmo, Z.; Skórska, C.; Sitkowska, J.; Dutkiewicz, J. Occupational biohazards in agricultural dusts from India. Ann. Agric. Environ. Med. 2000, 7, 133–139. [Google Scholar] [PubMed]
- Stephan, R.; Van Trappen, S.; Cleenwerck, I.; Vancanneyt, M.; De Vos, P.; Lehner, A. Enterobacter turicensis sp. nov. and Enterobacter helveticus sp. nov., isolated from fruit powder. Int. J. Syst. Evol. Microbiol. 2007, 57, 820–826. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.X.; Peng, G.X.; Wang, E.T.; Yan, H.; Yuan, Q.H.; Zhang, W.; Lou, X.; Wu, H.; Tan, Z.Y. Diverse endophytic nitrogen-fixing bacteria isolated from wild rice Oryza rufipogon and description of Phytobacter diazotrophicus gen. nov. sp. nov. Arch. Microbiol. 2007, 189, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Osdaghi, E.; Rahimi, T.; Taghavi, S.M.; Ansari, M.; Zarei, S.; Portier, P.; Briand, M.; Jacques, M.-A. Comparative genomics and phylogenetic analyses suggest several novel species within the genus Clavibacter, including nonpathogenic tomato-associated strains. Appl. Environ. Microbiol. 2020, 86, e02873-19. [Google Scholar] [CrossRef] [Green Version]
- Laroute, V.; Tormo, H.; Couderc, C.; Mercier-Bonin, M.; Le Bourgeois, P.; Cocaign-Bousquet, M.; Daveran-Mingot, M.-L. From Genome to Phenotype: An Integrative Approach to Evaluate the Biodiversity of Lactococcus lactis. Microorganisms 2017, 5, 27. [Google Scholar] [CrossRef]
- Latt, Z.K.; San, S.Y.; Kyaw, E.P.; Lynn, T.M.; Nwe, M.T.; Mon, W.W.; Aye, K.N. Using Cellulolytic Nitrogen Fixing Bacterium, Azomonas agilis for effective degradation of agricultural residues. Open Microbiol. J. 2018, 12, 154–162. [Google Scholar] [CrossRef]
- Compant, S.; Samad, A.; Faist, H.; Sessitsch, A. A review on the plant microbiome: Ecology, functions, and emerging trends in microbial application. J. Adv. Res. 2019, 19, 29–37. [Google Scholar] [CrossRef]
- Madhaiyan, M.; Saravanan, V.S.; Blom, J.; Smits, T.H.; Rezzonico, F.; Kim, S.J.; Weon, H.Y.; Kwon, S.W.; Whitman, W.B.; Ji, L. Phytobacter palmae sp. nov., a novel endophytic, N2 fixing, plant growth promoting Gammaproteobacterium isolated from oil palm (Elaeis guineensis Jacq.). Int. J. Syst. Evol. Microbiol. 2020, 70, 841–848. [Google Scholar] [CrossRef]
- Trias, R.; Bañeras, L.; Montesinos, E.; Badosa, E. Lactic acid bacteria from fresh fruit and vegetables as biocontrol agents of phytopathogenic bacteria and fungi. Int. J. Microbiol. 2008, 11, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Trotel-Aziz, P.; Couderchet, M.; Biagianti, S.; Aziz, A. Characterization of new bacterial biocontrol agents Acinetobacter, Bacillus, Pantoea and Pseudomonas spp. mediating grapevine resistance against Botrytis cinerea. Environ. Exp. Bot. 2008, 64, 21–32. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [Green Version]
- Thapa, S.P.; Davis, E.W.; Lyu, Q.; Weisberg, A.J.; Stevens, D.M.; Clarke, C.R.; Coaker, G.; Chang, J.H. The Evolution, Ecology, and Mechanisms of Infection by Gram-Positive, Plant-Associated Bacteria. Annu. Rev. Phytopathol. 2019, 57, 341–365. [Google Scholar] [CrossRef] [PubMed]
- Telias, A.; White, J.R.; Pahl, D.M.; Ottesen, A.R.; Walsh, C.S. Bacterial community diversity and variation in spray water sources and the tomato fruit surface. BMC Microbiol. 2011, 11, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enya, J.; Shinohara, H.; Yoshida, S.; Tsukiboshi, T.; Negishi, H.; Suyama, K.; Tsushima, S. Culturable leaf-associated bacteria on tomato plants and their potential as biological control agents. Microb. Ecol. 2007, 53, 524–536. [Google Scholar] [CrossRef]
- Mohammed, A.F.; Oloyede, A.R.; Odeseye, A.O. Biological control of bacterial wilt of tomato caused by Ralstonia solanacearum using Pseudomonas species isolated from the rhizosphere of tomato plants. Arch. Phytopathol. Plant Prot. 2020, 53, 1–16. [Google Scholar] [CrossRef]
- Abo-Elyousr, K.A.M.; Hassan, S.A. Biological control of Ralstonia solanacearum (Smith), the causal pathogen of bacterial wilt disease by using Pantoea spp. Egypt. J. Biol. Pest. Control 2021, 31, 113. [Google Scholar] [CrossRef]
- Ghadamgahi, F.; Tarighi, S.; Taheri, P.; Saripella, G.V.; Anzalone, A.; Kalyandurg, P.B.; Catara, V.; Ortiz, R.; Vetukuri, R.R. Plant Growth-Promoting Activity of Pseudomonas aeruginosa FG106 and Its Ability to Act as a Biocontrol Agent against Potato, Tomato and Taro Pathogens. Biology 2022, 11, 140. [Google Scholar] [CrossRef]
- Ashmawy, N.A.; Shoeib, A.A.; Youssef, H.F.B.; Mahmoud, S.M. Pathological, biochemical and molecular characterization of the seed-borne bacteria “Pantoea spp., Xanthomonas spp. and Pseudomonas spp.” from solanaceous plants in Egypt. J. Microbiol. Biotech. Food Sci. 2020, 10, 289–295. [Google Scholar] [CrossRef]
- Tian, Q.; Chuan, J.; Sun, X.; Zhou, A.; Wang, L.; Zou, J.; Zhao, W.; Li, X. Description of Clavibacter zhangzhiyongii sp. nov., a phytopathogenic actinobacterium isolated from barley seeds, causing leaf brown spot and decline. Int. J. Syst. Evol. Microbiol. 2021, 71, 004786. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorrasi, S.; Pasqualetti, M.; Muñoz-Palazon, B.; Novello, G.; Mazzucato, A.; Campiglia, E.; Fenice, M. Comparison of the Peel-Associated Epiphytic Bacteria of Anthocyanin-Rich “Sun Black” and Wild-Type Tomatoes under Organic and Conventional Farming. Microorganisms 2022, 10, 2240. https://doi.org/10.3390/microorganisms10112240
Gorrasi S, Pasqualetti M, Muñoz-Palazon B, Novello G, Mazzucato A, Campiglia E, Fenice M. Comparison of the Peel-Associated Epiphytic Bacteria of Anthocyanin-Rich “Sun Black” and Wild-Type Tomatoes under Organic and Conventional Farming. Microorganisms. 2022; 10(11):2240. https://doi.org/10.3390/microorganisms10112240
Chicago/Turabian StyleGorrasi, Susanna, Marcella Pasqualetti, Barbara Muñoz-Palazon, Giorgia Novello, Andrea Mazzucato, Enio Campiglia, and Massimiliano Fenice. 2022. "Comparison of the Peel-Associated Epiphytic Bacteria of Anthocyanin-Rich “Sun Black” and Wild-Type Tomatoes under Organic and Conventional Farming" Microorganisms 10, no. 11: 2240. https://doi.org/10.3390/microorganisms10112240
APA StyleGorrasi, S., Pasqualetti, M., Muñoz-Palazon, B., Novello, G., Mazzucato, A., Campiglia, E., & Fenice, M. (2022). Comparison of the Peel-Associated Epiphytic Bacteria of Anthocyanin-Rich “Sun Black” and Wild-Type Tomatoes under Organic and Conventional Farming. Microorganisms, 10(11), 2240. https://doi.org/10.3390/microorganisms10112240