


Euterpe oleracea Mart (Açaizeiro) from the Brazilian Amazon: A Novel Font of Fungi for Lipase Production

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Solvents
2.2. Isolation and Identification of Endophytic Fungi from the Fruits of E. oleracea Mart
2.3. Identification and Conservation of Endophytic Fungi
2.4. Determining the Enzymatic Activity
2.5. Enzymatic Test for Lipase in a Solid Environment
2.6. Lipase Production in a Liquid Environment
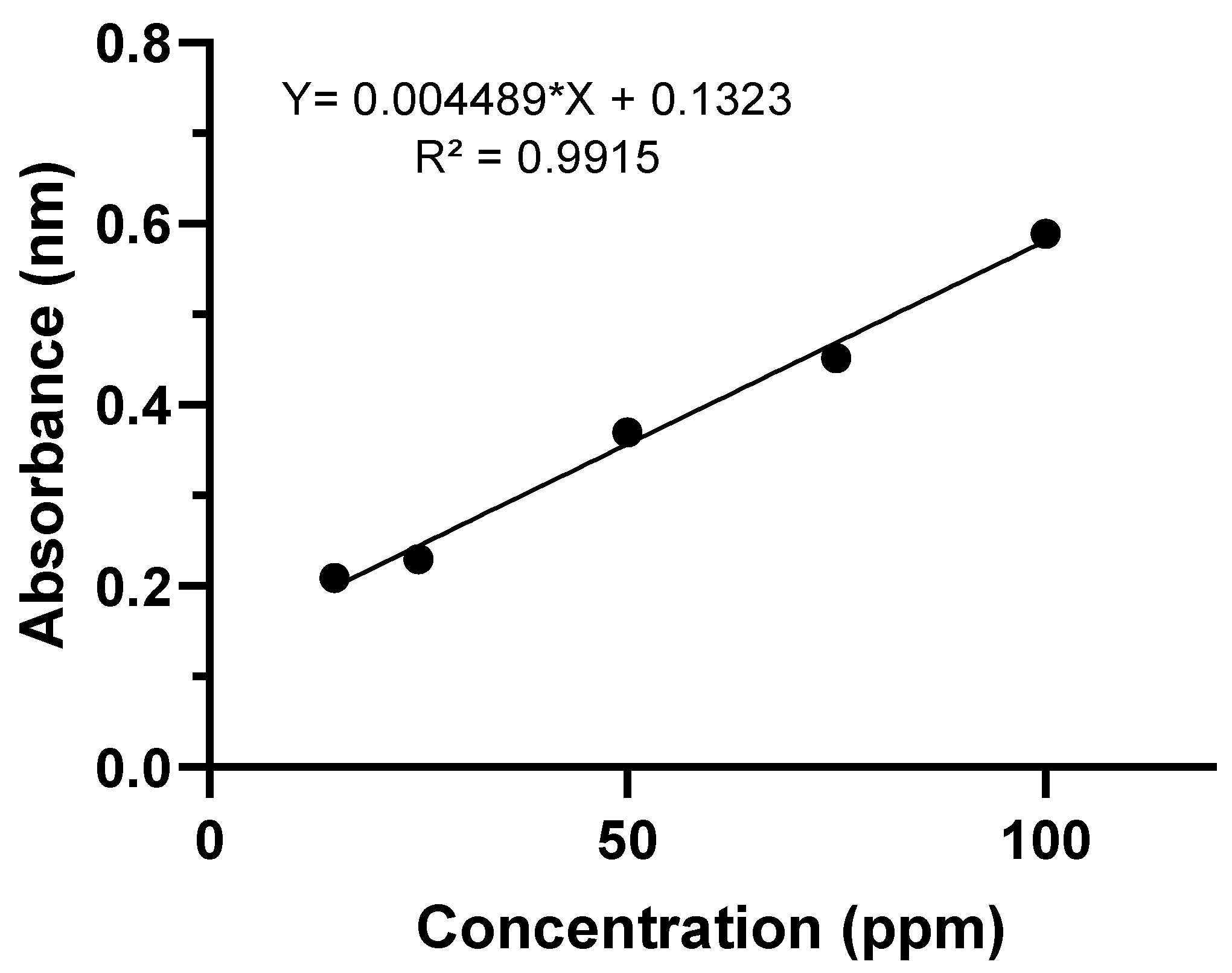
2.7. Quantification of Lipolytic Activity in a Liquid Medium
2.8. Statistical Analysis
Experimental Design and Statistical Model
3. Results and Discussion
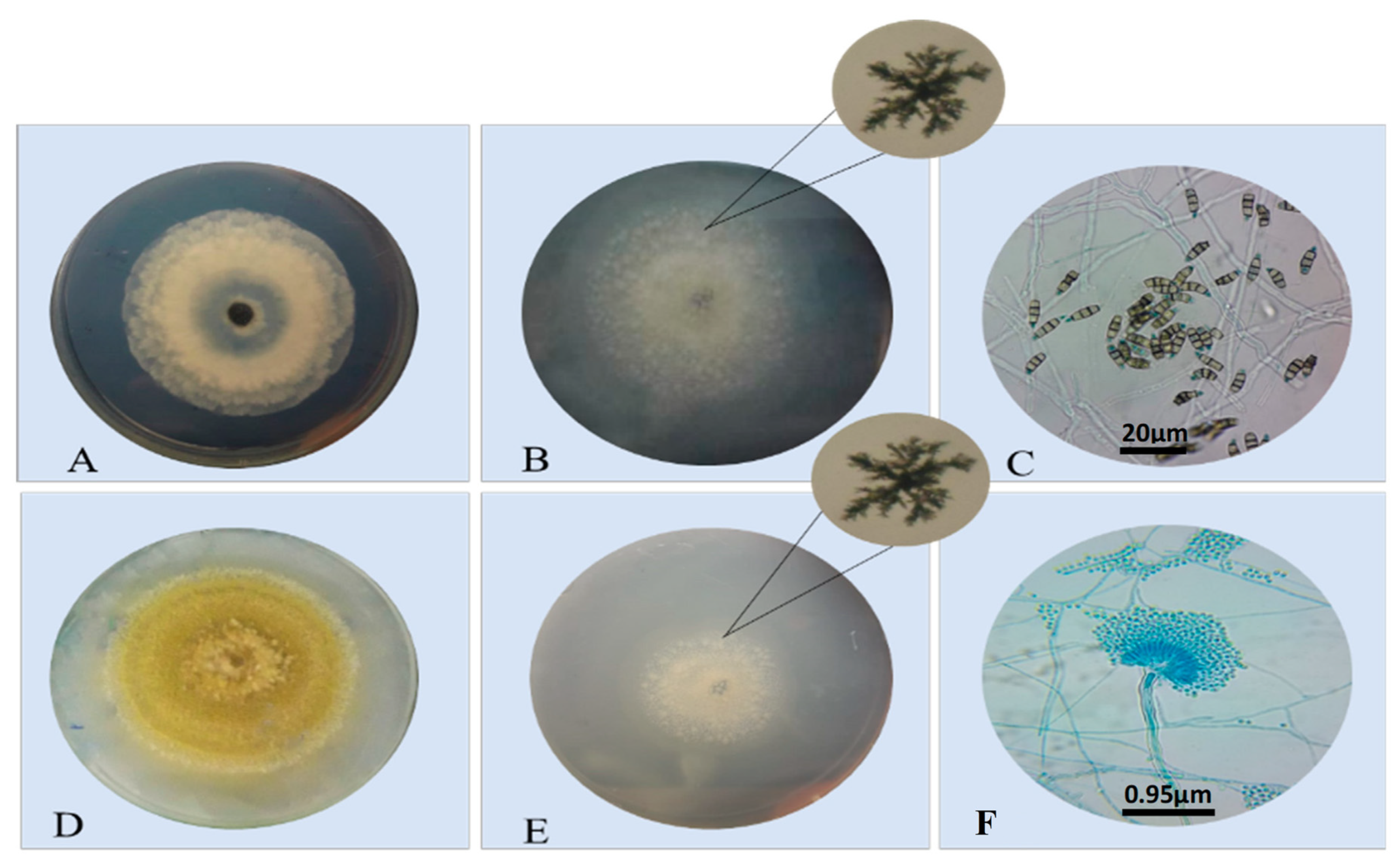
3.1. Isolating and Purifying Endophytic Fungi
3.2. Screening of Lipase-Producing Endophytic Fungi
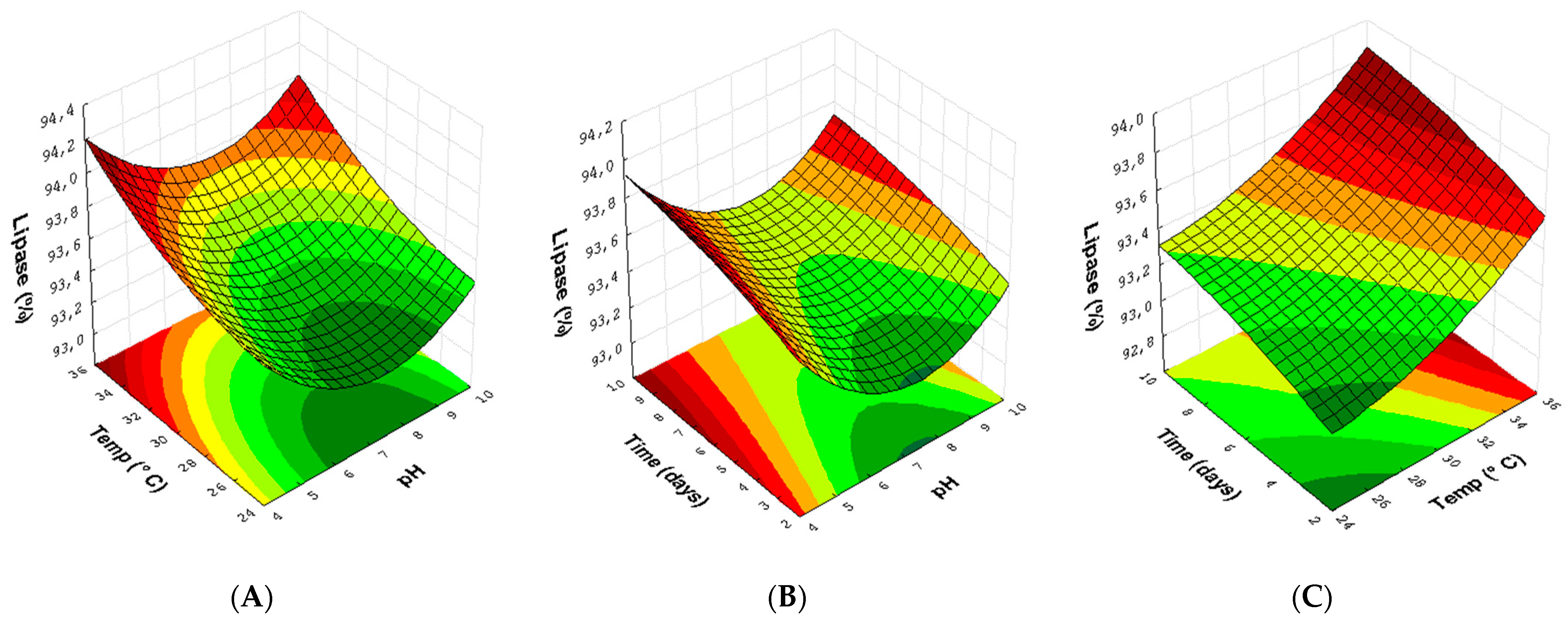
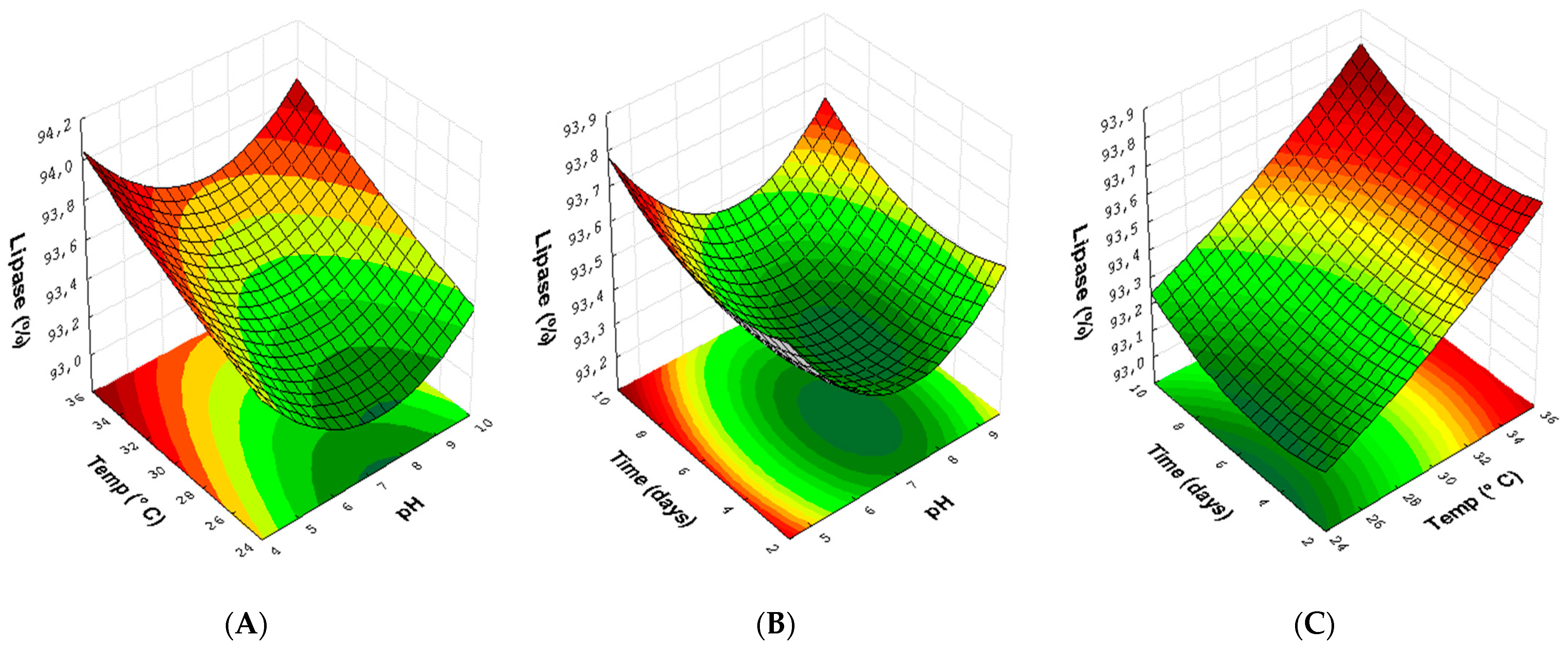
3.3. Experimental Design for Lipase with Endophytic Fungi Pestalotiopsis sp. (30) and Aspergillus sp. (24)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lewinsohn, T.M.; Prado, P.I. How Many Species Are There in Brazil? Conserv. Biol. 2005, 19, 619–624. [Google Scholar] [CrossRef]
- Rocha, K.S.C.; Queiroz, M.S.R.; Gomes, B.S.; Dallago, R.; de Souza, R.O.M.A.; Guimarães, D.O.; Itabaiana, I.; Leal, I.C.R. Lipases of Endophytic Fungi Stemphylium lycopersici and Sordaria Sp.: Application in the Synthesis of Solketal Derived Monoacylglycerols. Enzym. Microb. Technol. 2020, 142, 109664. [Google Scholar] [CrossRef] [PubMed]
- Rovere-Querini, P.; De Lorenzo, R.; Conte, C.; Brioni, E.; Lanzani, C.; Yacoub, M.R.; Chionna, R.; Martinenghi, S.; Vitali, G.; Tresoldi, M.; et al. Post-COVID-19 follow-up clinic: Depicting chronicity of a new disease. Acta Biomed. 2020, 91, 22–28. [Google Scholar] [PubMed]
- Lopes, E.; Soares-Filho, B.; Souza, F.; Rajão, R.; Merry, F.; Carvalho Ribeiro, S. Mapping the Socio-Ecology of Non Timber Forest Products (NTFP) Extraction in the Brazilian Amazon: The Case of Açaí (Euterpe precatoria Mart) in Acre. Landsc. Urban Plan. 2019, 188, 110–117. [Google Scholar] [CrossRef]
- De Souza, A.Q.L.; de Souza, A.D.L.; Filho, S.A.; Pinheiro, M.L.B.; Sarquis, M.I.D.M.; Pereira, J.O. Atividade Antimicrobiana de Fungos Endofíticos Isolados de Plantas Tóxicas Da Amazônia: Palicourea longiflora (Aubl.) Rich e Strychnos Cogens Bentham. Acta Amaz. 2004, 34, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, J.; Fishbane, P.M.; Gasiorowicz, S.G. Analysis of Physical Systems with Paraconsistent Annotated Logic: Introducing the Paraquantum Gamma Factor. J. Mod. Phys. 2000, 12, 624. [Google Scholar]
- Martins, G.R.; do Amaral, F.R.L.; Brum, F.L.; Mohana-Borges, R.; de Moura, S.S.T.; Ferreira, F.A.; Sangenito, L.S.; Santos, A.L.S.; Figueiredo, N.G.; Silva, A.S.A. da Chemical Characterization, Antioxidant and Antimicrobial Activities of Açaí Seed (Euterpe oleracea Mart.) Extracts Containing A- and B-Type Procyanidins. LWT 2020, 132, 109830. [Google Scholar] [CrossRef]
- de Souza, F.G.; de Araújo, F.F.; de Paulo Farias, D.; Zanotto, A.W.; Neri-Numa, I.A.; Pastore, G.M. Brazilian Fruits of Arecaceae Family: An Overview of Some Representatives with Promising Food, Therapeutic and Industrial Applications. Food Res. Int. 2020, 138, 109690. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; El Sayed, M.T.; Rady, A.; Zein, N.; Enan, G.; Shindia, A.; El-Hefnawy, S.; Sitohy, M.; Sitohy, B. Exploiting the Biosynthetic Potency of Taxol from Fungal Endophytes of Conifers Plants; Genome Mining and Metabolic Manipulation. Molecules 2020, 25, 3000. [Google Scholar] [CrossRef]
- Jagannath, S.; Konappa, N.; Lokesh, A.; Bhuvaneshwari; Dasegowda, T.; Udayashankar, A.C.; Chowdappa, S.; Cheluviah, M.; Satapute, P.; Jogaiah, S. Bioactive Compounds Guided Diversity of Endophytic Fungi from Baliospermum montanum and Their Potential Extracellular Enzymes. Anal. Biochem. 2021, 614, 114024. [Google Scholar] [CrossRef]
- Khan, A.L.; Halo, B.A.; Elyassi, A.; Ali, S.; Al-Hosni, K.; Hussain, J.; Al-Harrasi, A.; Lee, I.J. Indole Acetic Acid and ACC Deaminase from Endophytic Bacteria Improves the Growth of Solanum lycopersicum. Electron. J. Biotechnol. 2016, 21, 58–64. [Google Scholar] [CrossRef]
- Rana, K.L.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A.N.; Yadav, N.; Dhaliwal, H.S.; Saxena, A.K. Endophytic Microbes: Biodiversity, Plant Growth-Promoting Mechanisms and Potential Applications for Agricultural Sustainability; Springer International Publishing: Cham, Switzerland, 2020; Volume 113, ISBN 0123456789. [Google Scholar]
- Rodrigues, K.F. Endophytic Fungi in the Tropical Palm Euterpe oleracea Mart; University Microfilms International: Ann Arbor, MI, USA, 1992; p. 24. [Google Scholar]
- Rodrigues, K.F. The Foliar Fungal Endophytes of the Amazonian Palm Euterpe oleracea. Mycologia 1994, 86, 376–385. [Google Scholar] [CrossRef]
- Batista, B.N.; Raposo, N.V.D.M.; Da Silva, I.R. Isolamento e Avaliação Da Atividade Antimicrobiana de Fungos Endofíticos de Açaizeiro. Rev. Fitos 2018, 12, 161–174. [Google Scholar] [CrossRef]
- Peters, L.P.; Prado, L.S.; Silva, F.I.N.; Souza, F.S.C.; Carvalho, C.M. Selection of Endophytes as Antagonists of Colletotrichum gloeosporioides in Açaí Palm. Biol. Control 2020, 150, 104350. [Google Scholar] [CrossRef]
- McCulloch, J.A.; de Oliveira, V.M.; Pina, A.V.D.A.; Pérez-Chaparro, P.J.; de Almeida, L.M.; de Vasconcelos, J.M.; de Oliveira, L.F.; da Silva, D.E.A.; Rogez, H.; Cretenet, M.; et al. Complete Genome Sequence of Lactococcus lactis Strain AI06, an Endophyte of the Amazonian Açaí Palm. Genome Announc. 2014, 2, 2649–2650. [Google Scholar] [CrossRef] [Green Version]
- Colla, L.M.; Primaz, A.L.; Benedetti, S.; Loss, R.A.; de Lima, M.; Reinehr, C.O.; Bertolin, T.E.; Costa, J.A.V. Surface Response Methodology for the Optimization of Lipase Production under Submerged Fermentation by Filamentous Fungi. Brazilian J. Microbiol. 2016, 47, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Tacin, M.V.; Massi, F.P.; Fungaro, M.H.P.; Teixeira, M.F.S.; de Paula, A.V.; de Carvalho Santos-Ebinuma, V. Biotechnological Valorization of Oils from Agro-Industrial Wastes to Produce Lipase Using Aspergillus sp. from Amazon. Biocatal. Agric. Biotechnol. 2019, 17, 369–378. [Google Scholar] [CrossRef]
- Messias, J.M.; Da Costa, B.Z.; De Lima, V.M.G.; Giese, E.C.; Dekker, R.F.H.; Barbosa, A.D.M. Lipases Microbianas: Produção, Propriedades e Aplicações Biotecnológicas. Semin. Ciências Exatas E Tecnológicas 2011, 32, 213–234. [Google Scholar] [CrossRef] [Green Version]
- Kourist, R.; Guterl, J.K.; Miyamoto, K.; Sieber, V. Enzymatic Decarboxylation-An Emerging Reaction for Chemicals Production from Renewable Resources. ChemCatChem 2014, 6, 689–701. [Google Scholar] [CrossRef]
- Salihu, A.; Abbas, O.; Sallau, A.B.; Alam, M.Z. Agricultural Residues for Cellulolytic Enzyme Production by Aspergillus niger: Effects of Pretreatment. 3 Biotech 2015, 5, 1101–1106. [Google Scholar] [CrossRef] [Green Version]
- Filho, A.B.C.; Machado, B.Q.; Alves, A.U.; Pereira, B.d.J.; Guerra, N.M.; Neto, F.B. Bio-Agronomic Efficiency Indices of Eggplant and Tomato Intercropping. Hortic. Bras. 2022, 40, 181–189. [Google Scholar] [CrossRef]
- de Melo, I.S.; Silva, C.M.M.S.; Fay, E.Y.; Monteiro, P.R.T.; Dentzien, A.F.M. Degradation of atrazine by filamentous fungi. In Proceedings of the International Mycological Congress, Jerusalem, Israel, 1998. [Google Scholar]
- Mayordomo, I.; Randez-Gil, F.; Prieto, J.A. Isolation, Purification, and Characterization of a Cold-Active Lipase from Aspergillus nidulans. J. Agric. Food Chem. 2000, 48, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Southcott, K.A.; Johnson, J.A. Isolation of Endophytes from Two Species of Palm, from Bermuda. Can. J. Microbiol. 1997, 43, 789–792. [Google Scholar] [CrossRef]
- Jeewon, R.; Hyde, K.D. Establishing Species Boundaries and New Taxa among Fungi: Recommendations to Resolve Taxonomic Ambiguities. Mycosphere 2016, 7, 1669–1677. [Google Scholar] [CrossRef]
- Karakaya, A. First Report of Infection of Kiwifruit by Pestalotiopsis sp. in Turkey. Plant Dis. 2001, 85, 3082–3091. [Google Scholar] [CrossRef]
- Samson, R.A.; Noonim, P.; Meijer, M.; Houbraken, J.; Frisvad, J.C.; Varga, J. Diagnostic Tools to Identify Black Aspergillus. Stud. Mycol. 2007, 59, 129–145. [Google Scholar] [CrossRef]
- Klich, M.A. Identification of Common Aspergillus Species; Centraal Bureau voor Schim: Utrecht, The Netherlands, 2002; p. 116. [Google Scholar]
- Zhang, H.W.; Song, Y.C.; Tan, R.X. Biology and Chemistry of Endophytes. Nat. Prod. Rep. 2006, 23, 753–771. [Google Scholar] [CrossRef]
- Griebeler, N.; Polloni, A.E.; Remonatto, D.; Arbter, F.; Vardanega, R.; Cechet, J.L.; Di Luccio, M.; de Oliveira, D.; Treichel, H.; Cansian, R.L.; et al. Isolation and Screening of Lipase-Producing Fungi with Hydrolytic Activity. Food Bioprocess Technol. 2011, 4, 578–586. [Google Scholar] [CrossRef]
- Turki, S. Towards the Development of Systems for High-Yield Production of Microbial Lipases. Biotechnol. Lett. 2013, 35, 1551–1560. [Google Scholar] [CrossRef]
- Sandoval, G.; Marty, A. Screening Methods for Synthetic Activity of Lipases. Enzyme Microb. Technol. 2007, 40, 390–393. [Google Scholar] [CrossRef]
- Sarmah, N.; Rani, K.Y. Recent Advances on Sources and Industrial Applications of Lipases. Biotechnol Prog. 2018, 34, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Fabrício, H.; Birolli, W.G.; Morais, S.; Sena, I.S.; Ferreira, A.M.; Maria, S.; Faustino, M.; Grace, L.; Solon, S.; Porto, A.L.M.; et al. Biocatalysis and Agricultural Biotechnology Study of Biodegradation of Chloramphenicol by Endophytic Fungi Isolated from Bertholletia excelsa (Brazil Nuts). Biocatal. Agric. Biotechnol. 2019, 20, 101200. [Google Scholar] [CrossRef]
- Asghar, A.; Aziz, A.; Raman, A.; Mohd, W.; Wan, A. A Comparison of Central Composite Design and Taguchi Method for Optimizing Fenton Process. Sci. World J. 2014, 2014, 869120. [Google Scholar] [CrossRef] [Green Version]
- Beg, S.; Ii, V.; Case, P. Design of Experiments for Pharmaceutical Product Development; Springer: Berlin/Heidelberg, Germany, 2021; Volume II, ISBN 9789813343504. [Google Scholar]
- Reinehr, C.O.; Treichel, H. Production with Hydrolytic Activity by Aspergillus Using Agroindustrial Byproducts, Soy Oil and Glycerol. RECEN-Rev. Ciências Exatas E Nat. 2016, 18, 97–115. [Google Scholar]
- Souza, I.M.; Bassi, G.J.; Luiz, J.H.H.; Hirata, D.B. Isolation and Screening of Extracellular Lipase- Producing Endophytic Fungi from Handroanthus Impetiginosus. Asian J. Biotechnol. Bioresour. Technol. 2018, 4, 1–10. [Google Scholar] [CrossRef]
- Elango, D.; Manikandan, V.; Jayanthi, P.; Velmurugan, P.; Balamuralikrishnan, B.; Veera, A.; Subramaniam, M. Current Plant Biology Selection and Characterization of Extracellular Enzyme Production by an Endophytic Fungi Aspergillus Sojae and Its Bio-e Ffi Cacy Analysis against Cotton Leaf Worm, Spodoptera Litura. Curr. Plant Biol. 2020, 23, 100153. [Google Scholar] [CrossRef]
- Nema, A.; Patnala, S.H.; Mandari, V.; Kota, S.; Devarai, S.K. Production and Optimization of Lipase Using Aspergillus Niger MTCC 872 by Solid-State Fermentation. Bull. Natl. Res. Cent. 2019, 43, 82. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, A.; Vargas, J.; Rodrigues, M.; Mariano, A.B. Utilização de Resíduos da Agroindústria para a Produção de Enzimas Lipolíticas por Fermentação Submersa. Rev. Bras. de Prod. Agroindustriais 2013, 15, 19–26. [Google Scholar] [CrossRef]
- Ben Mefteh, F.; Frikha, F.; Daoud, A.; Bouket, A.C.; Luptakova, L.; Alenezi, F.N.; Al-Anzi, B.S.; Oszako, T.; Gharsallah, N.; Belbahri, L. Response Surface Methodology Optimization of an Acidic Protease Produced by Penicillium bilaiae Isolate TDPEF30, a Newly Recovered Endophytic Fungus from Healthy Roots of Date Palm Trees (Phoenix dactylifera L.). Microorganisms 2019, 7, 74. [Google Scholar] [CrossRef] [Green Version]
- Abd-elaziz, A.M. Enhanced Production, Partial Purification, and Characterization of Alkaline Thermophilic Protease from the Endophytic Fungus Aspergillus Ochraceus BT21. Egypt. Pharm. J. 2021, 19, 338–349. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Name | Levels | ||
|---|---|---|---|---|
| −1 | 0 | +1 | ||
| x11 | pH | 5 | 7 | 9 |
| x22 | Time (days) | 3 | 6 | 9 |
| x33 | Temperature (°C) | 25 | 30 | 35 |
| N° | Genus (Codig) | PZ * | N° | Genus (Codig) | PZ |
|---|---|---|---|---|---|
| 1 | Botryodiplodia sp. (30) | ++++ | 17 | Aspergillus sp. (26) | - |
| 2 | Fusarium sp. (2) | ++++ | 18 | Aspergillus sp. (11) | - |
| 3 | Penicillium sp. (5) | +++ | 19 | Penicillium sp. (12) | - |
| 4 | Penicillium sp. (7) | ++++ | 20 | Fusarium sp. (13) | - |
| 5 | Neocosmopora sp. (9) | ++++ | 21 | Mucor sp. (14) | - |
| 6 | Aspergillus sp. (21) | ++++ | 22 | Fusarium sp(15) | - |
| 7 | Aspergillus sp. (24) | ++++ | 23 | Penicillum sp. (16) | - |
| 8 | Fusarium sp. (27) | ++++ | 24 | Trichoderma sp. (17) | ++++ |
| 9 | Colletotrichum sp. (3) | ++++ | 25 | Verticillium sp. (18) | - |
| 10 | Pestalotiopsis sp. (31) | ++++ | 26 | Penicillium sp. (19) | - |
| 11 | Chaetomium sp. (35) | - | 27 | Penicillium sp. (20) | - |
| 12 | Aspergillus sp. (1) | - | 28 | Penicillium sp. (22) | - |
| 13 | Xylaria sp. (4) | - | 29 | Penicillium sp. (25) | - |
| 14 | Penicillium sp. (8) | - | 30 | Penicillium sp. (25) | - |
| 15 | Penicillium sp. (10) | - | 31 | Aspergillus sp. (29) | - |
| 16 | Aspergillus sp. (23) | - | 32 | Fusarium sp. (34) | - |
| Run | Coded and Uncoded Levels and Variables | Lipase (%) Pestalotiopsis sp. (30) | Lipase (%) Aspergilllus sp. (21) | |||||
|---|---|---|---|---|---|---|---|---|
| x1 (pH) | x2 (Temp.) | x3 (Time) | ||||||
| 1 | 5 | −1 | 25 | −1 | 6 | 0 | 93.40 | 93.23 |
| 2 | 5 | −1 | 30 | 0 | 3 | −1 | 93.23 | 93.66 |
| 3 | 5 | −1 | 30 | 0 | 9 | 1 | 93.84 | 93.49 |
| 4 | 5 | −1 | 35 | 1 | 6 | 0 | 94.00 | 93.87 |
| 5 | 7 | 0 | 25 | −1 | 3 | −1 | 93.18 | 93.12 |
| 6 | 7 | 0 | 25 | −1 | 9 | 1 | 93.22 | 93.28 |
| 7 | 7 | 0 | 35 | 1 | 3 | −1 | 93.61 | 93.50 |
| 8 | 7 | 0 | 35 | 1 | 9 | 1 | 93.68 | 93.70 |
| 9 | 9 | 1 | 25 | −1 | 6 | 0 | 93.34 | 93.29 |
| 10 | 9 | 1 | 30 | 0 | 3 | −1 | 93.41 | 93.40 |
| 11 | 9 | 1 | 30 | 0 | 9 | 1 | 93.54 | 93.56 |
| 12 | 9 | 1 | 35 | 1 | 6 | 0 | 93.67 | 93.61 |
| 13 | 7 | 0 | 30 | 0 | 6 | 0 | 93.36 | 93.40 |
| 14 | 7 | 0 | 30 | 0 | 6 | 0 | 93.37 | 93.31 |
| 15 | 7 | 0 | 30 | 0 | 6 | 0 | 92.38 | 93.24 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sena, I.S.; Ferreira, A.M.; Marinho, V.H.; e Holanda, F.H.; Borges, S.F.; de Souza, A.A.; de Carvalho R. Koga, R.; Lima, A.L.; Florentino, A.C.; Ferreira, I.M. Euterpe oleracea Mart (Açaizeiro) from the Brazilian Amazon: A Novel Font of Fungi for Lipase Production. Microorganisms 2022, 10, 2394. https://doi.org/10.3390/microorganisms10122394
Sena IS, Ferreira AM, Marinho VH, e Holanda FH, Borges SF, de Souza AA, de Carvalho R. Koga R, Lima AL, Florentino AC, Ferreira IM. Euterpe oleracea Mart (Açaizeiro) from the Brazilian Amazon: A Novel Font of Fungi for Lipase Production. Microorganisms. 2022; 10(12):2394. https://doi.org/10.3390/microorganisms10122394
Chicago/Turabian StyleSena, Iracirema S., Adriana M. Ferreira, Victor H. Marinho, Fabrício H. e Holanda, Swanny F. Borges, Agerdanio A. de Souza, Rosemary de Carvalho R. Koga, Adilson L. Lima, Alexandro C. Florentino, and Irlon M. Ferreira. 2022. "Euterpe oleracea Mart (Açaizeiro) from the Brazilian Amazon: A Novel Font of Fungi for Lipase Production" Microorganisms 10, no. 12: 2394. https://doi.org/10.3390/microorganisms10122394
APA StyleSena, I. S., Ferreira, A. M., Marinho, V. H., e Holanda, F. H., Borges, S. F., de Souza, A. A., de Carvalho R. Koga, R., Lima, A. L., Florentino, A. C., & Ferreira, I. M. (2022). Euterpe oleracea Mart (Açaizeiro) from the Brazilian Amazon: A Novel Font of Fungi for Lipase Production. Microorganisms, 10(12), 2394. https://doi.org/10.3390/microorganisms10122394