
Identification of a Green Algal Strain Collected from the Sarno River Mouth (Gulf of Naples, Italy) and Its Exploitation for Heavy Metal Remediation


 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sediment Sampling, Microalgae Isolation, and Culturing
2.2. Taxonomical Identification through 18S rRNA and rbcL Genes
2.3. Impact of HMs on Cell Growth and Morphology
2.4. Determination of HM Concentration in Microalgal Biomass
3. Results
3.1. Taxonomical Identification
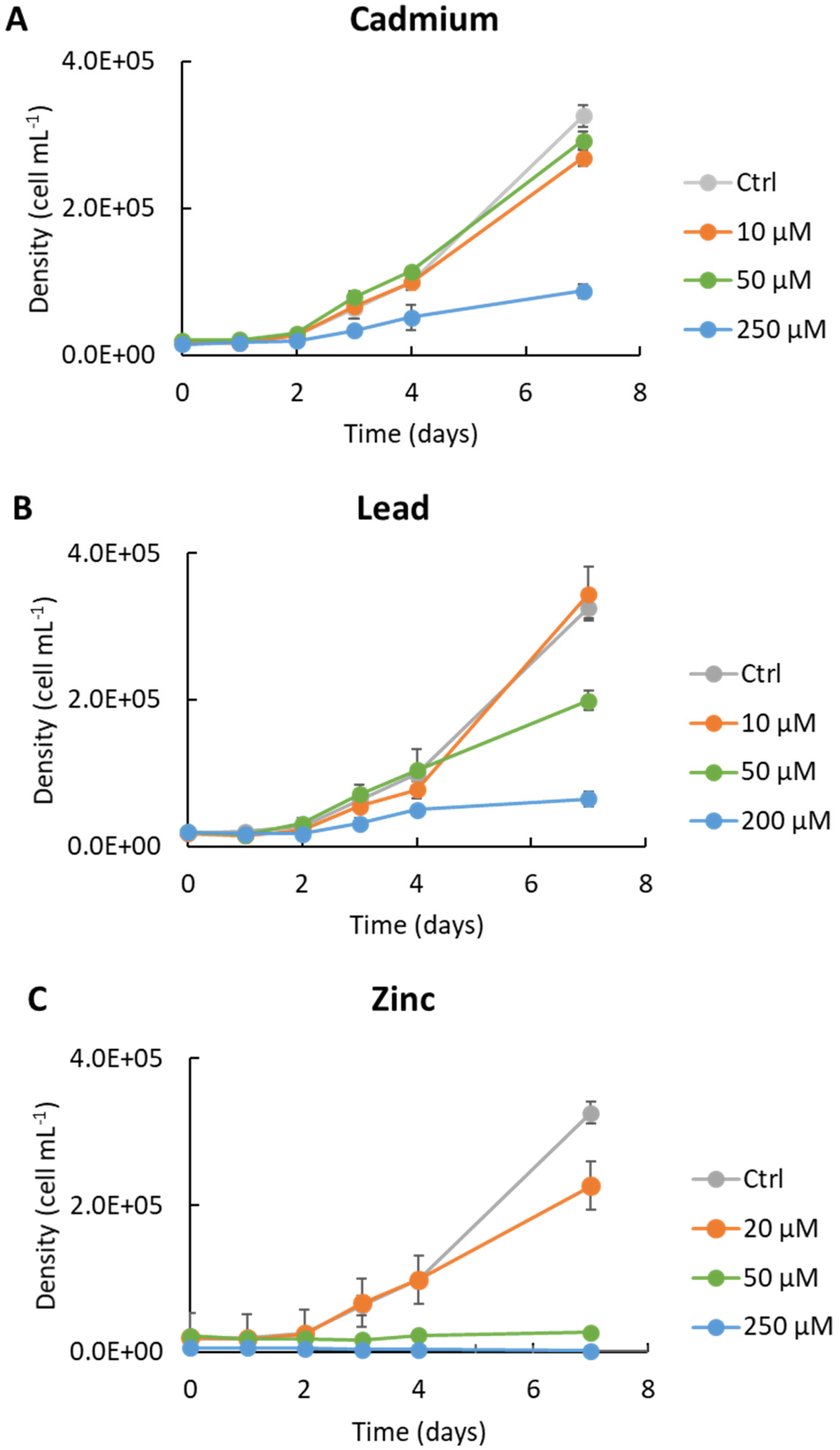
3.2. Response to Heavy Metal Stress
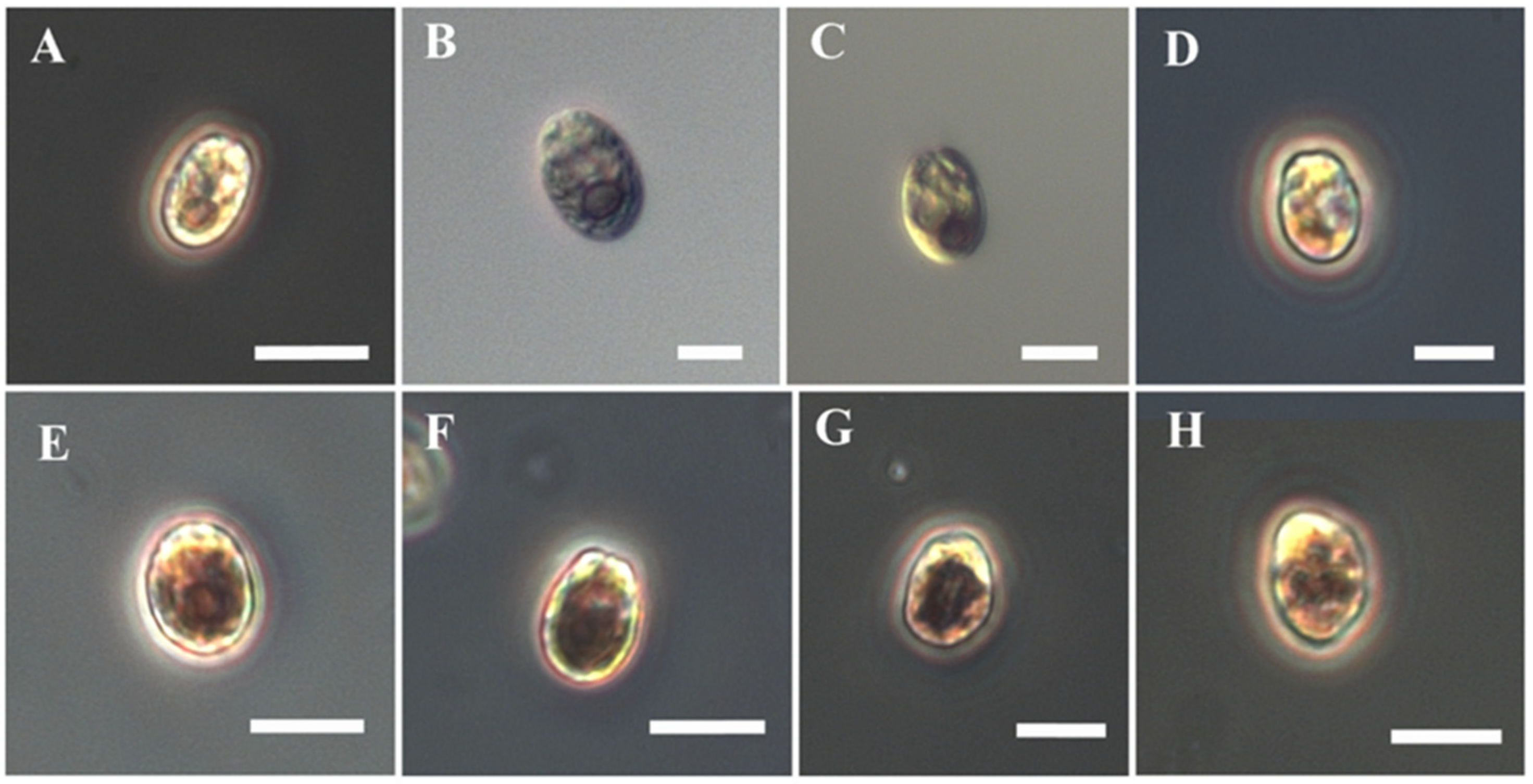
3.3. Effect on Cell Morphology
3.4. HM Sorption in Algal Biomass
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- García, E.M.; Cruz-Motta, J.J.; Farina, O.; Bastidas, C. Anthropogenic influences on heavy metals across marine habitats in the western coast of Venezuela. Cont. Shelf Res. 2008, 28, 2757–2766. [Google Scholar] [CrossRef]
- Sarker, S.; Vashistha, D.; Sarker, M.S.; Sarkar, A. DNA damage in marine rock oyster (Saccostrea Cucullata) exposed to environmentally available PAHs and heavy metals along the Arabian Sea coast. Ecotoxicol. Environ. Saf. 2018, 151, 132–143. [Google Scholar] [CrossRef]
- Yuan, W.K.; Zhou, Y.F.; Chen, Y.L.; Liu, X.N.; Wang, J. Toxicological effects of microplastics and heavy metals on the Daphnia magna. Sci. Total. Environ. 2020, 746, 141254. [Google Scholar] [CrossRef]
- Dirilgen, N. Accumulation of heavy metals in freshwater organisms: Assessment of toxic interactions. Turk. J. Chem. 2001, 25, 173–179. [Google Scholar]
- Yabanli, M.; Coskun, Y.; Oz, B. Detection of heavy metal concentrations in three freshwater fish species in lake Marmara (Western Anatolia) and heavy metal human health risk assessment. Fresenius Environ. Bull. 2015, 24, 4273–4278. [Google Scholar]
- Chon, H.T.; Lee, J.S.; Lee, J.U. Heavy Metal Contamination of Soil, Its Risk Assessment and Bioremediation. Geosystem Eng. 2011, 14, 191–206. [Google Scholar] [CrossRef]
- Armiento, G.; Caprioli, R.; Cerbone, A.; Chiavarini, S.; Crovato, C.; De Cassan, M.; De Rosa, L.; Montereali, M.R.; Nardi, E.; Nardi, L.; et al. Current status of coastal sediments contamination in the former industrial area of Bagnoli-Coroglio (Naples, Italy). Chem. Ecol. 2020, 36, 579–597. [Google Scholar] [CrossRef]
- Montuori, P.; Lama, P.; Aurino, S.; Naviglio, D.; Triassi, M. Metals loads into the Mediterranean Sea: Estimate of Sarno River inputs and ecological risk. Ecotoxicology 2013, 22, 295–307. [Google Scholar] [CrossRef] [Green Version]
- Tangherlini, M.; Corinaldesi, C.; Rastelli, E.; Musco, L.; Armiento, G.; Danovaro, R.; Dell’Anno, A. Chemical contamination can promote turnover diversity of benthic prokaryotic assemblages: The case study of the Bagnoli-Coroglio bay (southern Tyrrhenian Sea). Mar. Environ. Res. 2020, 160, 105040. [Google Scholar] [CrossRef]
- Das, A.; Osborne, J.W. Bioremediation of Heavy Metals. In Nanotechnology, Food Security and Water Treatment; Gothandam, K.M., Ranjan, S., Dasgupta, N., Ramalingam, C., Lichtfouse, E., Eds.; Environmental Chemistry for a Sustainable World; Springer: Berlin/Heidelberg, Germany, 2018; Volume 11, pp. 277–311. [Google Scholar]
- Leong, Y.K.; Chang, J.-S. Bioremediation of heavy metals using microalgae: Recent advances and mechanisms. Bioresour. Technol. 2020, 303, 122886. [Google Scholar] [CrossRef]
- Manikandan, A.; Babu, P.S.; Shyamalagowri, S.; Kamaraj, M.; Muthukumaran, P.; Aravind, J. Emerging role of microalgae in heavy metal bioremediation. J. Basic Microbiol. 2021, 62, 330–347. [Google Scholar] [CrossRef]
- Paul, O.; Jasu, A.; Lahiri, D.; Nag, M.; Ray, R.R. In Situ and ex situ bioremediation of heavy metals: The present scenario. J. Environ. Eng. Landsc. Manag. 2021, 29, 454–469. [Google Scholar] [CrossRef]
- Verma, S.; Kuila, A. Bioremediation of heavy metals by microbial process. Environ. Technol. Innov. 2019, 14, 100369. [Google Scholar] [CrossRef]
- Zeraatkar, A.K.; Ahmadzadeh, H.; Talebi, A.F.; Moheimani, N.R.; McHenry, M.P. Potential use of algae for heavy metal bioremediation, a critical review. J. Environ. Manag. 2016, 181, 817–831. [Google Scholar] [CrossRef]
- Choi, H.-J. Biosorption of Heavy Metals from Acid Mine Drainage by Modified Sericite and Microalgae Hybrid System. Water Air Soil Pollut. 2015, 226, 185. [Google Scholar] [CrossRef]
- Danouche, M.; El Ghachtouli, N.; El Arroussi, H. Phycoremediation mechanisms of heavy metals using living green microalgae: Physicochemical and molecular approaches for enhancing selectivity and removal capacity. Heliyon 2021, 7. [Google Scholar] [CrossRef]
- Goswami, R.K.; Agrawal, K.; Shah, M.P.; Verma, P. Bioremediation of heavy metals from wastewater: A current perspective on microalgae-based future. Lett. Appl. Microbiol. 2021, 75, 701–717. [Google Scholar] [CrossRef]
- Kumar, K.S.; Dahms, H.-U.; Won, E.-J.; Lee, J.-S.; Shin, K.-H. Microalgae – A promising tool for heavy metal remediation. Ecotoxicol. Environ. Saf. 2015, 113, 329–352. [Google Scholar] [CrossRef]
- Kwon, H.K.; Jeon, J.Y.; Oh, S.J. Potential for heavy metal (copper and zinc) removal from contaminated marine sediments using microalgae and light emitting diodes. Ocean Sci. J. 2017, 52, 57–66. [Google Scholar] [CrossRef]
- Rawat, I.; Gupta, S.K.; Shriwastav, A.; Singh, P.; Kumari, S.; Bux, F. Microalgae Applications in Wastewater Treatment. In Algae Biotechnology: Products and Processes; Bux, F., Chisti, Y., Eds.; Green Energy and Technology: Taiapei, Taiwan, 2016; pp. 249–268. [Google Scholar]
- Spain, O.; Plöhn, M.; Funk, C. The cell wall of green microalgae and its role in heavy metal removal. Physiol. Plant. 2021, 173, 526–535. [Google Scholar] [CrossRef]
- Ubando, A.T.; Africa, A.D.M.; Maniquiz-Redillas, M.C.; Culaba, A.B.; Chen, W.-H.; Chang, J.-S. Microalgal biosorption of heavy metals: A comprehensive bibliometric review. J. Hazard. Mater. 2021, 402, 123431. [Google Scholar] [CrossRef]
- Wilde, E.W.; Benemann, J.R. Bioremoval of heavy metals by the use of microalgae. Biotechnol. Adv. 1993, 11, 781–812. [Google Scholar] [CrossRef]
- Fu, Z.S.; Xi, S.H. The effects of heavy metals on human metabolism. Toxicol. Mech. Methods 2019, 30, 167–176. [Google Scholar] [CrossRef]
- Li, B.M.; Xia, M.S.; Zorec, R.; Parpura, V.; Verkhratsky, A. Astrocytes in heavy metal neurotoxicity and neurodegeneration. Brain Res. 2021, 1752, 147234. [Google Scholar] [CrossRef]
- Cao, D.-J.; Xie, P.-P.; Deng, J.-W.; Zhang, H.-M.; Ma, R.-X.; Liu, C.; Liu, R.-J.; Liang, Y.-G.; Li, H.; Shi, X.-D. Effects of Cu(2+) and Zn(2+) on growth and physiological characteristics of green algae, Cladophora. Environ. Sci. Pollut. Res. 2015, 22, 16535–16541. [Google Scholar] [CrossRef]
- Ling, N.; Li, W.L.; Xu, G.G.; Qi, Z.; Ji, C.F.; Liu, X.R.; Cui, D.; Sun, Y. Transcriptomic sequencing reveals the response of Dunaliella salina to copper stress via the increased photosynthesis and carbon mechanism. Mol. Omics 2021, 17, 769–782. [Google Scholar] [CrossRef]
- Atay, S.; Ozkoc, H.B. Effect of sediment on the bioavailability and toxicity of copper and zinc to a green alga. Fresenius Environ. Bull. 2010, 19, 3018–3027. [Google Scholar]
- Zhou, G.J.; Peng, F.-Q.; Zhang, L.-J.; Ying, G.-G. Biosorption of zinc and copper from aqueous solutions by two freshwater green microalgae Chlorella pyrenoidosa and Scenedesmus obliquus. Environ. Sci. Pollut. Res. 2012, 19, 2918–2929. [Google Scholar] [CrossRef]
- Cavalletti, E.; Romano, G.; Palma Esposito, F.; Barra, L.; Chiaiese, P.; Balzano, S.; Sardo, A. Copper Effect on Microalgae: Toxicity and Bioremediation Strategies. Toxics 2022, 10, 527. [Google Scholar] [CrossRef]
- García-García, J.D.; Sánchez-Thomas, R.; Moreno-Sánchez, R. Bio-recovery of non-essential heavy metals by intra- and extracellular mechanisms in free-living microorganisms. Biotechnol. Adv. 2016, 34, 859–873. [Google Scholar] [CrossRef]
- Lane, T.W.; Morel, F.M.M. A biological function for cadmium in marine diatoms. Proc. Natl. Acad. Sci. USA 2000, 97, 4627–4631. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Feng, L.; Jeffrey, P.D.; Shi, Y.; Morel, F.M. Structure and metal exchange in the cadmium carbonic anhydrase of marine diatoms. Nature 2008, 452, 56–61. [Google Scholar] [CrossRef]
- Abdel-Ghani, N.T.; El-Chaghaby, G. Biosorption for metal ions removal from aqueous solutions: A review of recent studies. Int. J. Latest Res. Sci. Technol. 2014, 3, 24–42. [Google Scholar]
- Ali, H.; Khan, E. Trophic transfer, bioaccumulation, and biomagnification of non-essential hazardous heavy metals and metalloids in food chains/webs—Concepts and implications for wildlife and human health. Hum. Ecol. Risk Assessment 2019, 25, 1353–1376. [Google Scholar] [CrossRef]
- Amqam, H.; Thalib, D.; Anwar, D.; Sirajuddin, S.; Mallongi, A. Human health risk assessment of heavy metals via consumption of fish from Kao Bay. Rev. Environ. Heal. 2020, 35, 257–263. [Google Scholar] [CrossRef]
- Hossain, M.B.; Ahmed, A.S.S.; Sarker, M.S.I. Human health risks of Hg, As, Mn, and Cr through consumption of fish, Ticto barb (Puntius ticto) from a tropical river, Bangladesh. Environ. Sci. Pollut. Res. 2018, 25, 31727–31736. [Google Scholar] [CrossRef]
- Albanese, S.; Iavazzo, P.; Adamo, P.; Lima, A.; De Vivo, B. Assessment of the environmental conditions of the Sarno river basin (south Italy): A stream sediment approach. Environ. Geochem. Health 2013, 35, 283–297. [Google Scholar] [CrossRef] [Green Version]
- Cicchella, D.; Giaccio, L.; Lima, A.; Albanese, S.; Cosenza, A.; Civitillo, D.; De Vivo, B. Assessment of the topsoil heavy metals pollution in the Sarno River basin, south Italy. Environ. Earth Sci. 2013, 71, 5129–5143. [Google Scholar] [CrossRef]
- De Pippo, T.; Donadio, C.; Guida, M.; Petrosino, C. The Case of Sarno River (Southern Italy). Effects of geomorphology on the environmental impacts. Environ. Sci. Pollut. Res. 2006, 13, 184–191. [Google Scholar] [CrossRef]
- Balzano, S.; Sardo, A.; Blasio, M.; Chahine, T.B.; Dell’Anno, F.; Sansone, C.; Brunet, C. Microalgal Metallothioneins and Phytochelatins and Their Potential Use in Bioremediation. Front. Microbiol. 2020, 11, 517. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Forján, E.; Navarro, F.; Cuaresma, M.; Vaquero, I.; Ruiz-Domínguez, M.C.; Gojkovic, Ž.; Vázquez, M.; Márquez, M.; Mogedas, B.; Bermejo, E.; et al. Microalgae: Fast-Growth Sustainable Green Factories. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1705–1755. [Google Scholar] [CrossRef]
- Huang, Z.Q.; Chen, B.Y.; Zhang, J.; Yang, C.L.; Wang, J.; Song, F.; Li, S.G. Absorption and speciation of arsenic by microalgae under arsenic-copper Co-exposure. Ecotoxicol. Environ. Saf. 2021, 213, 112024. [Google Scholar] [CrossRef]
- Cameron, H.; Mata, M.T.; Riquelme, C. The effect of heavy metals on the viability of Tetraselmis marina AC16-MESO and an evaluation of the potential use of this microalga in bioremediation. PeerJ 2018, 6, e5295. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum Book Publication Corporation: New York, NY, USA, 1975; pp. 29–60. [Google Scholar]
- Kooistra, W.; De Stefano, M.; Mann, D.G.; Salma, N.; Medlin, L.K. Phylogenetic position of Toxarium, a pennate-like lineage within centric diatoms (Bacillariophyceae). J. Phycol. 2003, 39, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Balzano, S.; Marie, D.; Gourvil, P.; Vaulot, D. Composition of the summer photosynthetic pico and nanoplankton communities in the Beaufort Sea assessed by T-RFLP and sequences of the 18S rRNA gene from flow cytometry sorted samples. ISME J. 2012, 6, 1480–1498. [Google Scholar] [CrossRef] [Green Version]
- Lepere, C.; Demura, M.; Kawachi, M.; Romac, S.; Probert, I.; Vaulot, D. Whole-genome amplification (WGA) of marine photosynthetic eukaryote populations. FEMS Microbiol. Ecol. 2011, 76, 513–523. [Google Scholar] [CrossRef] [Green Version]
- Pichard, S.L.; Campbell, L.; Paul, J.H. Diversity of the ribulose bisphosphate carboxylase/oxygenase form I gene (rbcL) in natural phytoplankton communities. Appl. Environ. Microbiol. 1997, 63, 3600–3606. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Balzano, S.; Sarno, D.; Kooistra, W.H.C.F. Effects of salinity on the growth rate and morphology of ten Skeletonema strains. J. Plankton Res. 2011, 33, 937–945. [Google Scholar] [CrossRef] [Green Version]
- Sunda, W.G.; Huntsman, S.A. Interactions among Cu2+, Zn2+, and Mn2+ in controlling cellular Mn, Zn, and growth rate in the coastal alga Chlamydomonas. Limnol. Oceanogr. 1998, 43, 1055–1064. [Google Scholar] [CrossRef]
- Palomba, E.; Chiaiese, P.; Termolino, P.; Paparo, R.; Filippone, E.; Mazzoleni, S.; Chiusano, M.L. Effects of Extracellular Self- and Nonself-DNA on the Freshwater Microalga Chlamydomonas reinhardtii and on the Marine Microalga Nannochloropsis gaditana. Plants 2022, 11, 1436. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Domozych, D.S. The crystalline cell wall of Tetraselmis convolutae (Chlorophyta), A freeze fracture analyses. J. Phycol. 1984, 20, 415–418. [Google Scholar] [CrossRef]
- Becker, B.; Melkonian, M.; Kamerling, J.P. The cell wall (theca) of Tetraselmis striata (Chlorophyta): Macromolecular composition and structural elements of the complex polysaccharides. J. Phycol. 1998, 34, 779–787. [Google Scholar] [CrossRef]
- Odyakov, V.F.; Zhizhina, E.G. A potentiometric study of precipitation in the (Pb2+-H2PO4--Cl-) system. Russ. J. Inorg. Chem. 2005, 50, 969–973. [Google Scholar]
- Schulze, P.S.C.; Carvalho, C.F.M.; Pereira, H.; Gangadhar, K.N.; Schüler, L.M.; Santos, T.F.; Varela, J.C.S.; Barreira, L. Urban wastewater treatment by Tetraselmis sp. CTP4 (Chlorophyta). Bioresour. Technol. 2017, 223, 175–183. [Google Scholar] [CrossRef]
- Pena, A.D.C.; Schaumloffel, L.D.; Trierweiler, L.F.; Gutterres, M. Tetraselmis sp. Isolated from a Microalgae Consortium for Tannery Wastewater Treatment. J. Soc. Leather Technol. Chem. 2018, 102, 258–261. [Google Scholar]
- Pérez-Rama, M.; Abalde, J.; Herrero, C.; Vaamonde, E.T. Cadmium removal by living cells of the marine microalga Tetraselmis suecica. Bioresour. Technol. 2002, 84, 265–270. [Google Scholar] [CrossRef]
- Da Costa, A.C.A.; De França, F.P. The behaviour of the microalgae Tetraselmis chuii in cadmium-contaminated solutions. Aquac. Int. 1998, 6, 57–66. [Google Scholar] [CrossRef]
- Perrein-Ettajani, H.; Amiard, J.C.; Haure, J.; Renaud, C. Effects of metals (Ag, Cd, Cu) on the biochemical composition and compartmentalization of these metals in two microalgae Skeletonema costatum and Tetraselmis suecica. Can. J. Fish. Aquat. Sci. 1999, 56, 1757–1765. [Google Scholar] [CrossRef]
- Perez-Rama, M.; Herrero, C.; Abalde, J.; Vaamonde, E.T. Class III metallothioneins in response to cadmium toxicity in the marine microalga Tetraselmis suecica (Kylin) Butch. Environ. Toxicol. Chem. 2001, 20, 2061–2066. [Google Scholar] [CrossRef]
- Kutlu, B.; Dorucu, M. Effects of heavy metal on Tetraselmis chuii. Fresenius Environ. Bull. 2021, 30, 13109–13117. [Google Scholar]
- Debelius, B.; Forja, J.M.; DelValls, A.; Lubián, L.M. Toxicity and bioaccumulation of copper and lead in five marine microalgae. Ecotoxicol. Environ. Saf. 2009, 72, 1503–1513. [Google Scholar] [CrossRef]
- Soto-Jiménez, M.F.; Arellano-Fiore, C.; Rocha-Velarde, R.; Jara-Marini, M.E.; Ruelas-Inzunza, J.; Voltolina, D.; Frías-Espericueta, M.G.; Quintero-Alvarez, J.M.; Páez-Osuna, F. Biological responses of a simulated marine food chain to lead addition. Environ. Toxicol. Chem. 2011, 30, 1611–1617. [Google Scholar] [CrossRef]
- Twining, B.S.; Baines, S.B. The Trace Metal Composition of Marine Phytoplankton. In Annual Review of Marine Science; Carlson, C.A., Giovannoni, S.J., Eds.; Annual Reviews: Palo Alto, CA, USA, 2013; Volume 5, pp. 191–215. [Google Scholar]
- Keller, M.D.; Selvin, R.C.; Claus, W.; Guillard, R.R.L. Media for the culture of oceanic ultraphytoplankton. J. Phycol. 2007, 23, 633–638. [Google Scholar] [CrossRef]
- Godinho, R.M.; Cabrita, M.T.; Alves, L.C.; Pinheiro, T. Imaging of intracellular metal partitioning in marine diatoms exposed to metal pollution: Consequences to cellular toxicity and metal fate in the environment. Metallomics 2014, 6, 1626–1631. [Google Scholar] [CrossRef]
- Santomauro, G.; Sun, W.-L.; Brümmer, F.; Bill, J. Incorporation of zinc into the coccoliths of the microalga Emiliania huxleyi. BioMetals 2016, 29, 225–234. [Google Scholar] [CrossRef]
- Li, J.; Schiavo, S.; Rametta, G.; Miglietta, M.L.; La Ferrara, V.; Wu, C.; Manzo, S. Comparative toxicity of nano ZnO and bulk ZnO towards marine algae Tetraselmis suecica and Phaeodactylum tricornutum. Environ. Sci. Pollut. Res. 2017, 24, 6543–6553. [Google Scholar] [CrossRef]
- Kumar, D.S.; Santhanam, P.; Ananth, S.; Shenbaga Devi, A.; Nandakumar, R.; Balaji Prasath, B.; Jeyanthi, S.; Jayalakshmi, T.; Ananthi, P. Effect of different dosages of zinc on the growth and biomass in five marine microalgae authors. Int. J. Fish. Aquac. 2014, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Levy, J.L.; Stauber, J.L.; Jolley, D.F. Sensitivity of marine microalgae to copper: The effect of biotic factors on copper adsorption and toxicity. Sci. Total. Environ. 2007, 387, 141–154. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Raouf, N.; Sholkamy, E.N.; Bukhari, N.; Al-Enazi, N.M.; Alsamhary, K.I.; Al-Khiat, S.H.A.; Ibraheem, I.B.M. Bioremoval capacity of Co+2 using Phormidium tenue and Chlorella vulgaris as biosorbents. Environ. Res. 2022, 204, 111630. [Google Scholar] [CrossRef]
- Merola, F.; Memmolo, P.; Miccio, L.; Savoia, R.; Mugnano, M.; Fontana, A.; D’Ippolito, G.; Sardo, A.; Iolascon, A.; Gambale, A.; et al. Tomographic flow cytometry by digital holography. Light. Sci. Appl. 2017, 6, e16241. [Google Scholar] [CrossRef] [Green Version]
- Das, P.; Thaher, M.I.; Hakim, M.; Al-Jabri, H.; Alghasal, G. A comparative study of the growth of Tetraselmis sp. in large scale fixed depth and decreasing depth raceway ponds. Bioresour. Technol. 2016, 216, 114–120. [Google Scholar] [CrossRef]
- Fon-Sing, S.; Borowitzka, M.A. Isolation and screening of euryhaline Tetraselmis spp. suitable for large-scale outdoor culture in hypersaline media for biofuels. J. Appl. Phycol. 2016, 28, 1–14. [Google Scholar] [CrossRef]
- Gojkovic, Z.; Guidi, F.; Bustamante, B.; Venuleo, M.; de Assunçao, P.; Portillo, E. Scaling-Up and Semi-Continuous Cultivation of Locally Isolated Marine Microalgae Tetraselmis striata in the Subtropical Island of Gran Canaria (Canary Islands, Spain). Processes 2021, 9, 1326. [Google Scholar] [CrossRef]
- Patrinou, V.; Daskalaki, A.; Kampantais, D.; Kanakis, D.C.; Economou, C.N.; Bokas, D.; Kotzamanis, Y.; Aggelis, G.; Vayenas, D.V.; Tekerlekopoulou, A.G. Optimization of Cultivation Conditions for Tetraselmis striata and Biomass Quality Evaluation for Fish Feed Production. Water 2022, 14, 3162. [Google Scholar] [CrossRef]
- Di Lena, G.; Casini, I.; Lucarini, M.; Lombardi-Boccia, G. Carotenoid profiling of five microalgae species from large-scale production. Food Res. Int. 2019, 120, 810–818. [Google Scholar] [CrossRef]
- Pereira, H.; Páramo, J.; Silva, J.; Marques, A.; Barros, A.; Maurício, D.; Santos, T.; Schulze, P.; Barros, R.; Gouveia, L.; et al. Scale-up and large-scale production of Tetraselmis sp. CTP4 (Chlorophyta) for CO2 mitigation: From an agar plate to 100-m3 industrial photobioreactors. Sci. Rep. 2018, 8, 5112. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment * | Metal | Concentration (mg/L) |
|---|---|---|
| Ctrl | - | - |
| Cd-10 | Cadmium (as CdCl2) | 1.1 |
| Cd-50 | 5.6 | |
| Cd-250 | 28.1 | |
| Pb-10 | Lead (as PbNO3) | 2.1 |
| Pb-50 | 10.4 | |
| Pb-200 | 22.4 | |
| Zn-20 | Zinc (as ZnSO4·7H2O) | 1.3 |
| Zn-50 | 3.3 | |
| Zn-250 | 16.4 |
| HM | Growth Rate Inhibition (% of Control) | Cell Surface (µm2) |
|---|---|---|
| Ctrl | - | 55 ± 7.9 |
| Cd-10 | 7.6 ± 2.9 | n.c. |
| Cd-50 | 6.7 ± 1.5 | n.c. |
| Cd-250 | 39 ± 3 | 66 ± 8 |
| Pb-10 | 0 | n.c. |
| Pb-50 | 20 ± 1.8 | n.c. |
| Pb-200 | 59 ± 4.7 | 64 ± 9.4 |
| Zn-20 | 16 ± 2.4 | n.c. |
| Zn-100 | 93 ± 1.3 | n.c. |
| Zn-250 | 100 ± 3.7 | 65 ± 9.4 |
| HM Typology and Molar Concentration (µM) (Theoretical Concentration) | Real HM Concentration Measured by ICP-MS (µM) | µg/mg | pg/cell |
|---|---|---|---|
| Pb 10 µM | 11.58 | 1.19 ± 0.06 | 2.04 ± 0.30 |
| Pb 50 µM | 47.78 | 9.77 ± 0.91 | 28.26 ± 2.00 |
| Cd 50 µM | 47.15 | 0.3 ± 0.00 | 0.67 ± 0.07 |
| Cd 250 µM | 226.85 | 0.31 ± 0.10 | 1.97 ± 0.11 |
| Zn 20 µM | 23.71 | 0.83 ± 0.03 | 2.56 ± 0.49 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barra, L.; Sardo, A.; Moros, M.; Smerilli, A.; Chiaiese, P.; Percopo, I.; Cavalletti, E.; Castro-Hinojosa, C.; Balzano, S. Identification of a Green Algal Strain Collected from the Sarno River Mouth (Gulf of Naples, Italy) and Its Exploitation for Heavy Metal Remediation. Microorganisms 2022, 10, 2445. https://doi.org/10.3390/microorganisms10122445
Barra L, Sardo A, Moros M, Smerilli A, Chiaiese P, Percopo I, Cavalletti E, Castro-Hinojosa C, Balzano S. Identification of a Green Algal Strain Collected from the Sarno River Mouth (Gulf of Naples, Italy) and Its Exploitation for Heavy Metal Remediation. Microorganisms. 2022; 10(12):2445. https://doi.org/10.3390/microorganisms10122445
Chicago/Turabian StyleBarra, Lucia, Angela Sardo, Maria Moros, Arianna Smerilli, Pasquale Chiaiese, Isabella Percopo, Elena Cavalletti, Christian Castro-Hinojosa, and Sergio Balzano. 2022. "Identification of a Green Algal Strain Collected from the Sarno River Mouth (Gulf of Naples, Italy) and Its Exploitation for Heavy Metal Remediation" Microorganisms 10, no. 12: 2445. https://doi.org/10.3390/microorganisms10122445
APA StyleBarra, L., Sardo, A., Moros, M., Smerilli, A., Chiaiese, P., Percopo, I., Cavalletti, E., Castro-Hinojosa, C., & Balzano, S. (2022). Identification of a Green Algal Strain Collected from the Sarno River Mouth (Gulf of Naples, Italy) and Its Exploitation for Heavy Metal Remediation. Microorganisms, 10(12), 2445. https://doi.org/10.3390/microorganisms10122445