
Endophytic Diaporthe as Promising Leads for the Development of Biopesticides and Biofertilizers for a Sustainable Agriculture



Abstract
:1. Introduction
2. Material and Methods
2.1. Criteria Used for Considering Studies
2.2. Search Strategy for Identification of Studies
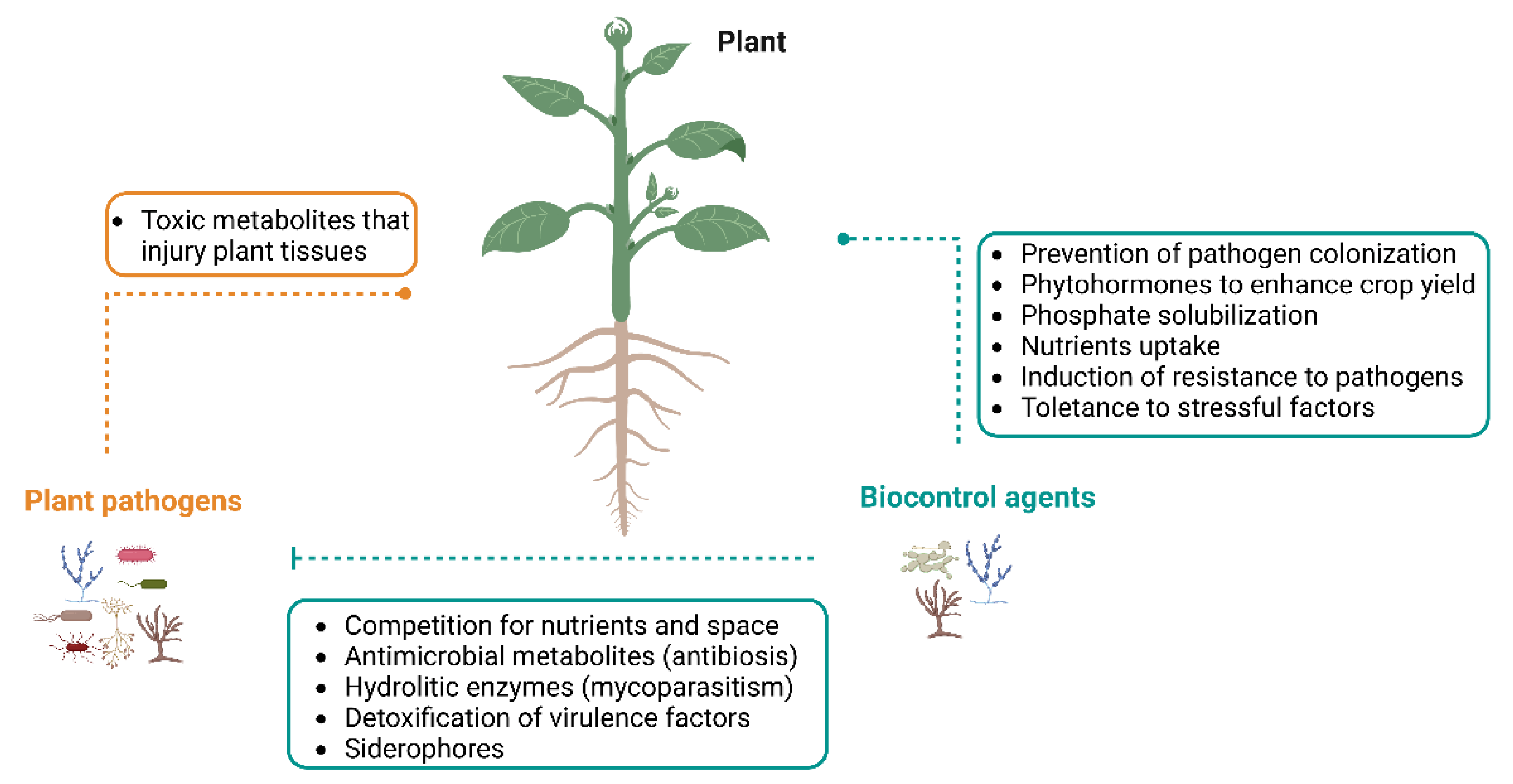
3. Fungal Endophytes and Their Benefits for Plants
4. Species of Diaporthe as Benefit Microorganisms to Agriculture
4.1. Production of Antimicrobial Compounds

{kind=link}
{kind=link}
| Host Plant | Endophyte | Antimicrobial Activity | References |
|---|---|---|---|
| Aconitum carmichaelii | Diaporthe sp. | Antifungal activity against the rice blast fungus Magnaporthe oryzae | [75] |
| Aconitum carmichaeli | D. amygdali | Antifungal activity against plant pathogenic fungi: Fusarium graminearum, Verticillium albo-atrum and Microdochium nivale | [76] |
| Actinidia chinensis | D. phragmatis (syn. D. eres) | Inhibitory activity against Pseudomonas syringae pv. actinidiae, the causal agent of kiwi canker disease | [73] |
| Balanophora polyandra | D. foeniculina | Antibacterial potential against plant pathogenic bacteria: Ralstonia solanacearum, Pseudomonas lachrymans and Xanthomonas vesicatoria | [77] |
| Cistus salvifolius | Diaporthe sp. | Antifungal activity against pathogens of agricultural importance: Phytophthora infestans, Botrytis cinerea and Septoria tritici | [78] |
| Copaifera pubiflora and Melocactus ernestii | D. miriciae | Antifungal activity against the grapevine pathogen D. rudis | [74] |
| Endodesmia calophylloides | Diaporthe sp. | Inhibition of zoospores of grapevine pathogen Plasmopara viticola | [79] |
| Espeletia sp. | D. phaseolorum | Antifungal activity against the plant pathogen Phytophthora infestans | [80] |
| Gossypium hirsutum and G. arboreum | D. longicolla and D. melonis | Antifungal activity against P. citricarpa | [72] |
| Gossypium hirsutum | Phomopsis-like species | Antifungal activity against Sclerotinia sclerotiorum, F. oxysporum, B. cinerea, Bipolaris sorokiniana, Gaeumannomyces graminis var. tritici, and Rhizoctonia cerealis | [81] |
| Maytenus ilicifolia | D. endophytica | Antifungal activity against the citrus pathogen Phyllosticta citricarpa | [82,83] |
| Mikania glomerata | D. citri | Antifungal activity against F. solani and Didymella bryoniae | [71] |
| Pachystachys lutea | Diaporthe sp. | Antifungal activity against the pathogenic fungi F. oxysporum and Colletotrichum sp. | [34] |
| Prunus domestica | D. eres | Antifungal activity against plant pathogenic fungi: Trichothecium roseum, F. avenaceum and Alternaria alternata | [17] |
| Rhizophora mucronata | Diaporthe sp. | Antifungal activity against pathogens of agricultural importance: Verticillium dahlia, Botrytis cinerea and Sclerotinia sclerotiorum | [84] |
| Schinus terebinthifolius | D. terebinthifolii | Antifungal activity against the citrus pathogen Phyllosticta citricarpa | [82,83] |
| Solanum lycopersicum | D. phaseolorum | Inhibitory activity against bacterial spot of tomato (Xanthomonas vesicatoria) | [85] |
| Vochysia divergens and Stryphnodendron adstringens | Diaporthe cf. heveae | Antifungal activity against P. citricarpa and Colletotrichum abscissum | [86] |
4.2. Phytotoxins as Potential Mycoherbicides
4.3. Plant-Growth Promoters
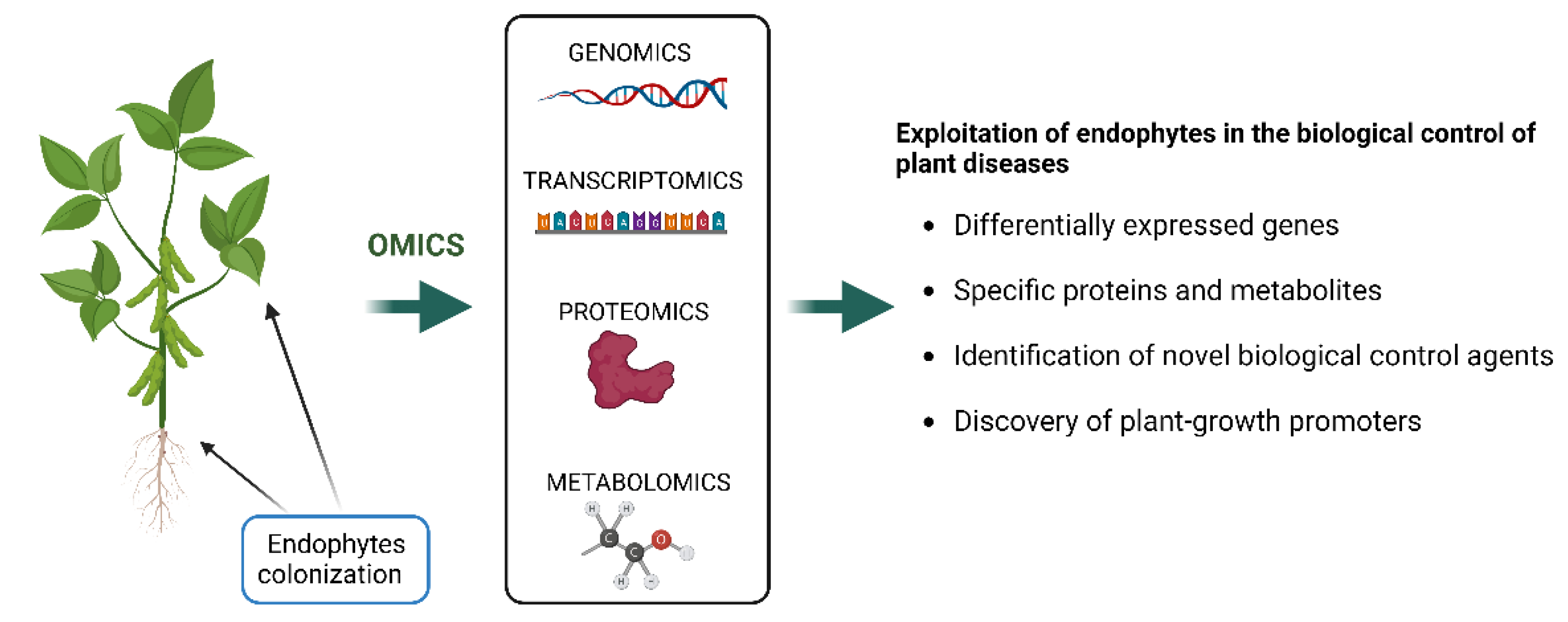
5. Omics to Explore the Secondary Metabolism of Diaporthe
5.1. Genomics
| Compound | Compound Nature | Biological Function | Species | References |
|---|---|---|---|---|
| ACR-Toxin I | Polyketide | Phytotoxin produced by the plant pathogenic fungus A. alternata, causing lemon leaf spot disease. | D. ampelina D. helianthi | [153,154] |
| ACT-Toxin II | Polyketide | Toxin causing brown spot disease on tangerine, produced by A. alternata | D. eres D. capsici D. citrisiana D. vaccinii (syn. D. eres) | [155] |
| Alternariol | Polyketide | Metabolite produced by Alternaria spp. that exhibits both phytotoxic and antifungal activity (e.g., zoosporicidal potential) | D. amygdali D. destruens D. capsici D. citri D. citrichinensis D. phragmitis (syn. D. eres) D. vaccinii (syn. D. eres) | [141] |
| Asperlactone | Polyketide | Methylsalicylic acid produced by Aspergillus westerdijkiae, antibacterial, antifungal and insecticidal activities | D. longicolla | [145,146] |
| Enniatin | Non-ribosomal peptide | Toxin produced by Fusarium spp., as a contaminant in cereals. It is also known as antibacterial, antifungal, and herbicidal. | D. citrichinensis | [156,157] |
| Fusarin | Polyketide | Mycotoxin produced mainly by fungi of the genus Fusarium, which can infect agriculturally important crops. | D. amygdali D. aspalathi D. helianthi | [158,159] |
| Fusicoccin | Terpene | Phytotoxin produced by D. amygdali, with plant-growth promoting potential | D. amygdali | [131] |
| (-)-Mellein | Phenolic compound | Metabolite firstly isolated from Alternaria melleus, with phytotoxic and antimicrobial activity | D. capsici D. citri D. citrichinensis D. destruens D. phragmitis (syn. D. eres) D. vaccinii (syn. D. eres) D. longicolla | [142] |
5.2. Transcriptomics
5.3. Proteomics
5.4. Metabolomics
6. Conclusions and Future Prospects
- (1)
- Reliable tools to enhance plant health and growth;
- (2)
- Novel strategies for mitigating the impacts of climate changes;
- (3)
- An ecological and sustainable agriculture.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Large, E.C. Control of Potato Blight (Phytophthora infestans) by Spraying with Suspensions of Metallic Copper. Nature 1943, 151, 80–81. [Google Scholar] [CrossRef]
- Brauer, V.S.; Rezende, C.P.; Pessoni, A.M.; De Paula, R.G.; Rangappa, K.S.; Nayaka, S.C.; Gupta, V.K.; Almeida, F. Antifungal agents in agriculture: Friends and foes of public health. Biomolecules 2019, 9, 521. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Li, X.Q.; Zhao, D.L.; Zhang, P. Antifungal secondary metabolites produced by the fungal endophytes: Chemical diversity and potential use in the development of biopesticides. Front. Microbiol. 2021, 12, 689527. [Google Scholar] [CrossRef]
- De Silva, N.I.; Brooks, S.; Lumyong, S.; Hyde, K.D. Use of endophytes as biocontrol agents. Fungal Biol. Rev. 2019, 33, 133–148. [Google Scholar] [CrossRef]
- Talibi, I.; Boubaker, H.; Boudyach, E.H.; Ait Ben Aoumar, A. Alternative methods for the control of postharvest citrus diseases. J. Appl. Microbiol. 2014, 117, 1–17. [Google Scholar] [CrossRef]
- Wang, Z.; Li, C.; White, J. Effects of Epichloë endophyte infection on growth, physiological properties and seed germination of wild barley under saline conditions. J. Agron. Crop Sci. 2019, 206, 43–51. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, C.; Xiang, X.; Zhang, Q.; Zhao, W.; Wei, G.; Hu, A. Uptake and transport of antibiotic kasugamycin in castor bean (Ricinus communis L.) seedlings. Front. Microbiol. 2022, 13, 948171. [Google Scholar] [CrossRef]
- Osada, H. Discovery and applications of nucleoside antibiotics beyond polyoxin. J. Antibiot. 2019, 72, 855–864. [Google Scholar] [CrossRef]
- El-Naggar, N.E.A. Streptomyces-based cell factories for production of biomolecules and bioactive metabolites. In Microbial Cell Factories Engineering for Production of Biomolecules; Academic Press: Cambridge, MA, USA, 2021; pp. 183–234. [Google Scholar]
- Chakraborty, A.; Ray, P. Mycoherbicides for the Noxious Meddlesome: Can Colletotrichum be a Budding Candidate? Front. Microbiol. 2021, 12, 754048. [Google Scholar] [CrossRef]
- Suebrasri, T.; Harada, H.; Jogloy, S.; Ekprasert, J.; Boonlue, S. Auxin-producing fungal endophytes promote growth of sunchoke. Rhizosphere 2020, 16, 100271. [Google Scholar] [CrossRef]
- Domka, A.M.; Rozpaadek, P.; Turnau, K. Are fungal endophytes merely mycorrhizal copycats? The role of fungal endophytes in the adaptation of plants to metal toxicity. Front. Microbiol. 2019, 10, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, Y.; Luan, P.; Wang, K.; Li, G.; Yin, Y.; Yang, Y.; Zhang, Q.; Liu, Y. Antifungal Activity and Plant Growth-Promoting Properties of Bacillus mojovensis B1302 against Rhizoctonia cerealis. Microorganisms 2022, 10, 1682. [Google Scholar] [CrossRef] [PubMed]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.; Kannangara, S.D.; Promputtha, I. Fungi vs. fungi in biocontrol: An overview of fungal antagonists applied against fungal plant pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Singh, H.; Bhardwaj, N.; Bhardwaj, S.K.; Khatri, M.; Kim, K.H.; Peng, W. An exploration on the toxicity mechanisms of phytotoxins and their potential utilities. Crit. Rev. Environ. Sci. Technol. 2022, 52, 395–435. [Google Scholar] [CrossRef]
- Xu, T.C.; Lu, Y.H.; Wang, J.F.; Song, Z.Q.; Hou, Y.G.; Liu, S.S.; Liu, C.S.; Wu, S.H. Bioactive secondary metabolites of the genus Diaporthe and anamorph Phomopsis from terrestrial and marine habitats and endophytes: 2010–2019. Microorganisms 2021, 9, 217. [Google Scholar] [CrossRef]
- Abramczyk, B.; Marzec-Grządziel, A.; Grządziel, J.; Król, E.; Gałązka, A.; Oleszek, W. Biocontrol Potential and Catabolic Profile of Endophytic Diaporthe eres Strain 1420S from Prunus domestica L. in Poland—A Preliminary Study. Agronomy 2022, 12, 165. [Google Scholar] [CrossRef]
- Hyde, K.D.; Soytong, K. The fungal endophyte dilemma. Fungal Divers. 2008, 33, e173. [Google Scholar]
- Chitnis, V.R.; Suryanarayanan, T.S.; Nataraja, K.N.; Prasad, S.R.; Oelmüller, R.; Shaanker, R.U. Fungal endophyte-mediated crop improvement: The way ahead. Front. Plant Sci. 2020, 11, 1588. [Google Scholar] [CrossRef]
- Baron, N.C.; Rigobelo, E.C. Endophytic fungi: A tool for plant growth promotion and sustainable agriculture. Mycology 2022, 13, 39–55. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kim, Y.-H.; Kang, S.-M.; Lee, J.-H.; Lee, I.-J. Gibberellins producing endophytic Aspergillus fumigatus sp. LH02 influenced endogenous phytohormonal levels, isoflavonoids production and plant growth in salinity stress. Proc. Biochem. 2011, 46, 440–447. [Google Scholar] [CrossRef]
- Khan, A.L.; Hamayun, M.; Kang, S.-M.; Kim, Y.-H.; Jung, H.-Y.; Lee, J.-H.; Lee, I.-J. Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: An example of Paecilomyces formosus LHL10. BMC Microbiol. 2012, 12, 3. [Google Scholar] [CrossRef] [PubMed]
- García-Latorre, C.; Rodrigo, S.; Santamaria, O. Effect of fungal endophytes on plant growth and nutrient uptake in Trifolium subterraneum and Poa pratensis as affected by plant host specificity. Mycol. Prog. 2021, 20, 1217–1231. [Google Scholar] [CrossRef]
- Baron, N.C.; Costa, N.T.A.; Mochi, D.A.; Rigobelo, E.C. First report of Aspergillus sydowii and Aspergillus brasiliensis as phosphorus solubilizers in maize. Ann Microbiol. 2018, 68, 863–870. [Google Scholar] [CrossRef]
- Christian, N.; Herre, E.A.; Clay, K. Foliar endophytic fungi alter patterns of nitrogen uptake and distribution in Theobroma cacao. New Phytol. 2019, 222, 1573–1583. [Google Scholar] [CrossRef] [PubMed]
- Rajini, S.B.; Nandhini, M.; Udayashankar, A.C.; Niranjana, S.R.; Lund, O.S.; Prakash, H.S. Diversity, plant growth-promoting traits, and biocontrol potential of fungal endophytes of Sorghum bicolor. Plant Pathol. 2020, 69, 642–654. [Google Scholar] [CrossRef]
- Khan, S.A.; Hamayun, M.; Yoon, H.; Kim, H.Y.; Suh, S.J.; Hwang, S.K.; Kim, J.M.; Lee, I.J.; Choo, Y.S.; Yoon, U.H.; et al. Plant growth promotion and Penicillium citrinum. BMC Microbiol. 2008, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Baron, N.C.; de Souza Pollo, A.; Rigobelo, E.C. Purpureocillium lilacinum and Metarhizium marquandii as plant growth-promoting fungi. PeerJ 2020, 8, e9005. [Google Scholar] [CrossRef]
- Ismail, H.M.; Hussain, A.; Iqbal, A.; Khan, S.A.; Lee, I.J. Aspergillus niger boosted heat stress tolerance in sunflower and soybean via regulating their metabolic and antioxidant system. J. Plant Interact. 2020, 15, 223–232. [Google Scholar] [CrossRef]
- Fenibo, E.O.; Ijoma, G.N.; Matambo, T. Biopesticides in sustainable agriculture: A critical sustainable development driver governed by green chemistry principles. Front. Sustain. Food Syst. 2021, 5, 619058. [Google Scholar] [CrossRef]
- Adhikari, K.; Bhandari, S.; Niraula, D.; Shrestha, J. Use of neem (Azadirachta indica A. Juss) as a biopesticide in agriculture: A review. J. Agric. Appl. Biol. 2021, 1, 100–117. [Google Scholar] [CrossRef]
- Kalpana, T.; Anil, K. A review of biopesticides and their plant phytochemicals information. Ann. Romanian Soc. Cell Biol. 2021, 25, 3576–3588. [Google Scholar]
- Santos, C.M.; da Silva, R.A.; Garcia, A.; Polli, A.D.; Polonio, J.C.; Azevedo, J.L.; Pamphile, J.A. Enzymatic and antagonist activity of endophytic fungi from Sapindus saponaria L. (Sapindaceae). Acta Biol. Colomb. 2019, 24, 322. [Google Scholar] [CrossRef]
- Ribeiro, A.D.S.; Polonio, J.C.; Costa, A.T.; Dos Santos, C.M.; Rhoden, S.A.; Azevedo, J.L.; Pamphile, J.A. Bioprospection of culturable endophytic fungi associated with the ornamental plant Pachystachys lutea. Curr. Microbiol. 2018, 75, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Landum, M.; Felix, M.; Alho, J.; Garcia, R.; Cabrita, M.J.; Rei, F.; Varanda, C.M. Antagonistic activity of fungi of Olea europaea L. against Colletotrichum acutatum. Microbiol. Res. 2016, 183, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Rakshith, D.; Santosh, P.; Satish, S. Isolation and characterization of antimicrobial metabolite producing endophytic Phomopsis sp. from Ficus pumila Linn. (Moraceae). Int. J. Chem. Anal. Sci. 2013, 4, 156–160. [Google Scholar] [CrossRef]
- Deketelaere, S.; Tyvaert, L.; França, S.C.; Höfte, M. Desirable traits of a good biocontrol agent against Verticillium wilt. Front. Microbiol. 2017, 8, 1186. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K.; Sharma, M. Approaches and Trends in Plant Disease Management; Scientific Publishers: Jodhpur, India, 2014; p. 429. [Google Scholar]
- Roberts, W. Studies on biogenesis. Philos. Trans. R. Soc. 1874, 164, 466. [Google Scholar]
- Baker, K.F. Evolving concepts of biological control of plant pathogens. Annu. Rev. Phytopathol. 1987, 25, 67–85. [Google Scholar] [CrossRef]
- Weindling, R. Experimental consideration of the mold toxins of Gliocladium and Trichoderma. Phytopathology 1941, 31, 991. [Google Scholar]
- Howell, C.R. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, D.C.; He, M.H.; Amalin, D.M.; Liu, W.; Alvindia, D.G.; Zhan, J. Biological control of plant diseases: An evolutionary and eco-economic consideration. Pathogens 2021, 10, 1311. [Google Scholar] [CrossRef] [PubMed]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.S.; Patra, J.K. Endophytes: A treasure house of bioactive compounds of medicinal importance. Front. Microbiol. 2016, 7, 1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmukh, S.K.; Gupta, M.K.; Prakash, V.; Saxena, S. Endophytic Fungi: A Source of Potential Antifungal Compounds. J. Fungi 2018, 4, 77. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-H.; He, J.; Li, X.-N.; Huang, R.; Song, F.; Chen, Y.-W.; Miao, C.-P. Guaiane sesquiterpenes and isopimarane diterpenes from an endophytic fungus Xylaria sp. Phytochemistry 2014, 105, 197–204. [Google Scholar] [CrossRef]
- Sangeetha, G.; Usharani, S.; Muthukumar, A. Biocontrol with Trichoderma species for the management of postharvest crown rot of banana. Phytopathol. Mediterr. 2009, 48, 214–225. [Google Scholar]
- Bai, Y.B.; Gao, Y.Q.; Nie, X.D.; Tuong, T.M.; Li, D.; Gao, J.M. Antifungal activity of griseofulvin derivatives against phytopathogenic fungi in vitro and in vivo and three-dimensional quantitative structure-activity relationship analysis. J. Agric. Food Chem. 2019, 67, 6125–6132. [Google Scholar] [CrossRef]
- Segaran, G.; Sathiavelu, M. Fungal endophytes: A potent biocontrol agent and a bioactive metabolites reservoir. Biocatal. Agric. Biotechnol. 2019, 21, 101284. [Google Scholar] [CrossRef]
- Sessa, L.; Abreo, E.; Lupo, S. Diversity of fungal latent pathogens and true endophytes associated with fruit trees in Uruguay. J. Phytopathol. 2018, 166, 633–647. [Google Scholar] [CrossRef]
- Udayanga, D.; Liu, X.; McKenzie, E.H.C.; Chukeatirote, E.; Bahkali, A.H.A.; Hyde, K.D. The genus Phomopsis: Biology, applications, species concepts and names of common phytopathogens. Fungal Divers. 2011, 50, 189–225. [Google Scholar] [CrossRef]
- Arciuolo, R.; Camardo Leggieri, M.; Chiusa, G.; Castello, G.; Genova, G.; Spigolon, N.; Battilani, P. Ecology of Diaporthe eres, the causal agent of hazelnut defects. PLoS ONE 2021, 16, e0247563. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Renderos, W.; Bevis, E.; Hebb, J.; Abbasi, P.A. Diaporthe eres causes stem cankers and death of young apple rootstocks in Canada. Can. J. Plant Pathol. 2020, 42, 218–227. [Google Scholar] [CrossRef]
- Udayanga, D.; Castlebury, L.A.; Rossman, A.Y.; Chukeatirote, E.; Hyde, K.D. Insights into the genus Diaporthe: Phylogenetic species delimitation in the D. eres species complex. Fungal Divers. 2014, 67, 203–229. [Google Scholar] [CrossRef] [Green Version]
- Gomes, R.R.; Glienke, C.; Videira, S.I.R.; Lombard, L.; Groenewald, J.Z.; Crous, P.W. Diaporthe: A genus of endophytic, saprobic and plant pathogenic fungi. Persoonia 2013, 31, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Manawasinghe, I.S.; Huang, Y.; Shu, Y.; Phillips, A.J.L.; Dissanayake, A.J.; Hyde, K.D.; Xiang, M.; Luo, M. Endophytic Diaporthe associated with Citrus grandis cv. Tomentosa in China. Front. Microbiol. 2021, 11, 609387. [Google Scholar] [CrossRef]
- Guarnaccia, V.; Crous, P.W. Emerging citrus diseases in Europe caused by Diaporthe spp. IMA Fungus 2017, 8, 317–334. [Google Scholar] [CrossRef] [Green Version]
- Chepkirui, C.; Stadler, M. The genus Diaporthe: A rich source of diverse and bioactive metabolites. Mycol. Prog. 2017, 16, 477–494. [Google Scholar] [CrossRef]
- Chen, H.; Huang, M.; Li, X.; Liu, L.; Chen, B.; Wang, J.; Lin, Y. Phochrodines A-D, first naturally occurring new chromenopyridines from mangrove entophytic fungus Phomopsis sp. 33#. Fitoterapia 2018, 124, 103–107. [Google Scholar]
- Sharma, V.; Singamaneni, V.; Sharma, N.; Kumar, A.; Arora, D.; Kushwaha, M.; Bhushan, S.; Jaglan, S.; Gupta, P. Valproic acid induces three novel cytotoxic secondary metabolites in Diaporthe sp., an endophytic fungus from Datura inoxia Mill. Bioorganic Med. Chem. Lett. 2018, 28, 2217–2221. [Google Scholar] [CrossRef]
- Song, H.C.; Qin, D.; Han, M.J.; Wang, L.; Zhang, K.; Dong, J.Y. Bioactive 2-pyrone metabolites from an endophytic Phomopsis asparagi SWUKJ5.2020 of Kadsura angustifolia. Phytochem. Lett. 2017, 22, 235–240. [Google Scholar] [CrossRef]
- Zhang, W.; Xu, L.; Yang, L.; Huang, Y.; Li, S.; Shen, Y. Phomopsidone A, a novel depsidone metabolite from the mangrove endophytic fungus Phomopsis sp. A123. Fitoterapia 2014, 96, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Krohn, K.; Farooq, U.; Hussain, H.; Ahmed, I.; Rheinheimer, J.; Draeger, S.; Schulz, B.; van Ree, T. Phomosines H-J, novel highly substituted biaryl ethers, isolated from the endophytic fungus Phomopsis sp. from Ligustrum vulgare. Nat. Prod. Commun. 2011, 6, 1907–1912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebastianes, F.L.S.; Cabedo, N.; El Aouad, N.; Valente, A.M.M.P.; Lacava, P.T.; Azevedo, J.L.; Pizzirani-Kleiner, A.A.; Cortes, D. 3-Hydroxypropionic acid as an antibacterial agent from endophytic fungi Diaporthe phaseolorum. Curr. Microbiol. 2012, 65, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Goddard, M.L.; Mottier, N.; Jeanneret-Gris, J.; Christen, D.; Tabacchi, R.; Abou-Mansour, E. Differential production of phytotoxins from Phomopsis sp. from grapevine plants showing esca symptoms. J. Agric. Food Chem. 2014, 62, 8602–8607. [Google Scholar] [CrossRef]
- Tanney, J.B.; McMullin, D.R.; Green, B.D.; Miller, J.D.; Seifert, K.A. Production of antifungal and antiinsectan metabolites by the Picea endophyte Diaporthe maritima sp. nov. Fungal Biol. 2016, 120, 1448–1457. [Google Scholar] [CrossRef]
- Wulansari, D.; Julistiono, H.; Nurkanto, A.; Agusta, A. Antifungal activity of (+)-2,20-epicytoskyrin A and its membrane-disruptive action. Makara J. Sci. 2016, 20, 160–166. [Google Scholar] [CrossRef]
- Ferreira, M.C.; Cantrell, C.L.; Wedge, D.E.; Gonçalves, V.N.; Jacob, M.R.; Khan, S.; Rosa, C.A.; Rosa, L.H. Antimycobacterial and antimalarial activities of endophytic fungi associated with the ancient and narrowly endemic neotropical plant Vellozia gigantea from Brazil. Mem. Inst. Oswaldo Cruz 2017, 112, 692–697. [Google Scholar] [CrossRef]
- Liu, Y.; Ruan, Q.; Jiang, S.; Qu, Y.; Chen, J.; Zhao, M.; Yang, B.; Liu, Y.; Zhao, Z.; Cui, H. Cytochalasins and polyketides from the fungus Diaporthe sp. GZU-1021 and their anti-inflammatory activity. Fitoterapia 2019, 137, 104187. [Google Scholar] [CrossRef]
- Polonio, J.C.; Almeida, T.; Garcia, A.; Mariucci, G.; Azevedo, J.; Rhoden, S.; Pamphile, J.A. Biotechnological prospecting of foliar endophytic fungi of guaco (Mikania glomerata Spreng.) with antibacterial and antagonistic activity against phytopathogens. Genet. Mol. Res. 2015, 14, 7297–7309. [Google Scholar] [CrossRef]
- Verma, P.; Hiremani, N.S.; Gawande, S.P.; Sain, S.K.; Nagrale, D.T.; Narkhedkar, N.G.; Prasad, Y.G. Modulation of plant growth and antioxidative defense system through endophyte biopriming in cotton (Gossypium spp.) and non-host crops. Heliyon 2022, 8, e09487. [Google Scholar] [CrossRef]
- Yu, J.J.; Yang, H.X.; Zhang, F.L.; He, J.; Li, Z.H.; Liu, J.K.; Feng, T. Secondary metabolites from cultures of the kiwi-associated fungus Diaporthe phragmitis and their antibacterial activity assessment. Phytochem. Lett. 2021, 46, 143–148. [Google Scholar] [CrossRef]
- Carvalho, C.D.; Ferreira-D’Silva, A.; Wedge, D.E.; Cantrell, C.L.; Rosa, L.H. Antifungal activities of cytochalasins produced by Diaporthe miriciae, an endophytic fungus associated with tropical medicinal plants. Can. J. Microbiol. 2018, 64, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Ma, K.X.; Xie, X.S.; Wang, T.; Wu, S.H. Secondary metabolites of an endophytic fungus Phomopsis sp. Chem. Nat. Compd. 2015, 51, 392–394. [Google Scholar] [CrossRef]
- Ma, K.X.; Shen, X.T.; Huang, R.; Wang, T.; Xie, X.S.; Liu, S.W.; Wu, S.H.; He, J. Bioactive metabolites produced by the endophytic fungus Phomopsis sp. YM355364. Nat. Prod. Commun. 2014, 9, 669–670. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Wang, W.; Wang, X.; Shahid, H.; Yang, Y.; Wang, Y.; Wang, S.; Shan, T. Diversity and communities of culturable endophytic fungi from the root holoparasite Balanophora polyandra Griff. and their antibacterial and antioxidant activities. Ann. Microbiol. 2022, 72, 19. [Google Scholar] [CrossRef]
- Hussain, H.; Ahmed, I.; Schulz, B.; Draeger, S.; Krohn, K. Pyrenocines J-M: Four new pyrenocines from the endophytic fungus, Phomopsis sp. Fitoterapia 2012, 83, 523–526. [Google Scholar] [CrossRef]
- Talontsi, F.M.; Islam, M.T.; Facey, P.; Douanla-Meli, C.; von Tiedemann, A.; Laatsch, H. Depsidones and other constituents from Phomopsis sp. CAFT69 and its host plant Endodesmia calophylloides with potent inhibitory effect on motility of zoospores of grapevine pathogen Plasmopara viticola. Phytochem. Lett. 2012, 3, 657–664. [Google Scholar] [CrossRef]
- Prada, H.; Avila, L.; Sierra, R.; Bernal, A.; Restrepo, S. Morphological and molecular characterization of the antagonistic interaction between the endophyte Diaporthe sp. isolated from frailejon (Espeletia sp.) and the plant pathogen Phytophthora infestans. Rev. Iberoam. Micol. 2009, 26, 198–201. [Google Scholar] [CrossRef]
- Fu, J.; Zhou, Y.; Li, H.F.; Ye, Y.H.; Guo, J.H. Antifungal metabolites from Phomopsis sp. By254, an endophytic fungus in Gossypium hirsutum. Afr. J. Microbiol. Res. 2011, 5, 1231–1236. [Google Scholar]
- Santos, P.J.; Savi, D.C.; Gomes, R.R.; Goulin, E.H.; Da Costa Senkiv, C.; Tanaka, F.A.; Almeida, Á.M.; Galli-Terasawa, L.; Kava, V.; Glienke, C. Diaporthe endophytica and D. terebinthifolii from medicinal plants for biological control of Phyllosticta citricarpa. Microbiol. Res. 2016, 186–187, 153–160. [Google Scholar] [CrossRef]
- Tonial, F.; Maia, B.H.; Sobottka, A.M.; Savi, D.C.; Vicente, V.A.; Gomes, R.R.; Glienke, C. Biological activity of Diaporthe terebinthifolii extracts against Phyllosticta citricarpa. FEMS Microbiol. Lett. 2017, 364, fnx026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiono, Y.; Sasaki, T.; Shibuya, F.; Yasuda, Y.; Koseki, T.; Supratman, U. Isolation of a phomoxanthone A derivative, a new metabolite of tetrahydroxanthone, from a Phomopsis sp. isolated from the mangrove, Rhizhopora mucronata. Nat. Prod. Commun. 2013, 8, 1735–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, T.S. Bioactive metabolites from tomato endophytic fungi with antibacterial activity against tomato bacterial spot disease. Rhizosphere 2021, 17, 100292. [Google Scholar] [CrossRef]
- Noriler, S.A.; Savi, D.C.; Aluizio, R.; Palacio-Cortes, A.M.; Possiede, Y.M.; Glienke, C. Bioprospecting and structure of fungal endophyte communities found in the Brazilian biomes, Pantanal, and Cerrado. Front. Microbiol. 2018, 9, 1526. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, A.; Santos, S. Sustainable approach to weed management: The role of precision weed management. Agronomy 2022, 12, 118. [Google Scholar] [CrossRef]
- Esposito, M.; Crimaldi, M.; Cirillo, V. Drone and sensor technology for sustainable weed management: A review. Chem. Biol. Technol. Agric. 2021, 8, 18. [Google Scholar] [CrossRef]
- Bjawa, A.A.; Mahajan, G.; Chauhan, B.S. Nonconventional weed management strategies for modern agriculture. Weed Sci. 2015, 63, 723–747. [Google Scholar] [CrossRef]
- Mia, M.J.; Massetani, F.; Murri, G.; Neri, D. Sustainable alternatives to chemicals for weed control in the orchard—A Review. Hortic. Sci. 2020, 47, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Bastos, B.D.O.; Deobald, G.A.; Brun, T.; Dal Prá, V.; Junges, E.; Kuhn, R.C.; Pinto, A.K.; Mazutti, M.A. Solid-state fermentation for production of a bioherbicide from Diaporthe sp. and its formulation to enhance the efficacy. Biotech 2017, 7, 135. [Google Scholar] [CrossRef] [Green Version]
- Brun, T.; Rabuske, J.E.; Luft, L.; Confortin, T.C.; Todero, I.; Aita, B.C.; Zabot, G.L.; Mazutti, M.A. Powder containing biomolecules from Diaporthe schini for weed control. Environ. Technol. 2022, 43, 2135–2144. [Google Scholar] [CrossRef]
- Cimmino, A.; Cinelli, T.; Masi, M.; Reveglia, P.; da Silva, M.A.; Mugnai, L.; Michereff, S.J.; Surico, G.; Evidente, A. Phytotoxic lipophilic metabolites produced by grapevine strains of Lasiodiplodia species in Brazil. J. Agric. Food Chem. 2017, 65, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Macías-Rubalcava, M.L.; García-Méndez, M.C.; King-Díaz, B.; Macías-Ruvalcaba, N.A. Effect of phytotoxic secondary metabolites and semisynthetic compounds from endophytic fungus Xylaria feejeensis strain SM3e-1b on spinach chloroplast photosynthesis. J. Photochem. Photobiol. B 2017, 166, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Andolfi, A.; Boari, A.; Evidente, M.; Cimmino, A.; Vurro, M.; Ash, G.; Evidente, A. Gulypyrones A and B and phomentrioloxins B and C Produced by Diaporthe gulyae, a potential mycoherbicide for saffron thistle (Carthamus lanatus). J. Nat. Prod. 2015, 78, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.R.C.; Baldoni, D.B.; Lima, J.; Porto, V.; Marcuz, C.; Ferraz, R.C.; Kuhn, R.C.; Jacques, R.J.S.; Guedes, J.V.C.; Mazutti, M.A. Bioherbicide production by Diaporthe sp. isolated from the Brazilian Pampa biome. Biocatal. Agric. Biotechnol. 2015, 4, 575–578. [Google Scholar] [CrossRef]
- Souza, A.R.C.; Baldoni, D.B.; Lima, J.; Porto, V.; Marcuz, C.; Machado, C.; Ferraz, R.C.; Kuhn, R.C.; Jacques, R.J.S.; Guedes, J.V.C.; et al. Selection, isolation, and identification of fungi for bioherbicide production. Braz. J. Microbiol. 2016, 48, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Pes, M.P.; Mazutti, M.A.; Almeida, T.C.; Curioletti, L.E.; Melo, A.A.; Guedes, J.V.C.; Kuhn, R.C. Bioherbicide based on Diaporthe sp. secondary metabolites in the control of three tough weeds. Afr. J. Agric. Res. 2016, 11, 4242–4249. [Google Scholar]
- Briscoe, W.E. Identification of a Phytotoxic Fungus and an Investigation into the Isolation of Its Phytotoxic Constituents. Ph.D. Tesis, University of Mississippi, Oxford, MS, USA, 2014. [Google Scholar]
- Cimmino, A.; Andolfi, A.; Zonno, M.C.; Boari, A.; Troise, C.; Motta, A.; Vurro, M.; Ash, C.; Evidente, A. Phomentrioloxin, a fungal phytotoxin with potential herbicidal activity, and its derivatives: A structure–activity relationship study. J. Agric. Food Chem. 2013, 61, 9645–9649. [Google Scholar] [CrossRef]
- Evidente, M.; Boari, A.; Vergura, S.; Cimmino, A.; Vurro, M.; Ash, G.; Superchi, S.; Evidente, A. Structure and absolute configuration of kongiidiazadione, a new phytotoxic 3-substituted-5-diazenylcyclopentendione produced by Diaporthe Kongii. Chirality 2015, 27, 557–562. [Google Scholar] [CrossRef]
- Yang, Z.; Ge, M.; Yin, Y.; Chen, Y.; Luo, M.; Chen, D. A novel phytotoxic nonenolide from Phomopsis sp. HCCB03520. Chem. Biodivers. 2012, 9, 403–408. [Google Scholar] [CrossRef]
- Meepagala, K.M.; Briscoe, W.E.; Techen, N.; Johnson, R.D.; Clausen, B.M.; Duke, S.O. Isolation of a phytotoxic isocoumarin from Diaporthe eres-infected Hedera helix (English ivy) and synthesis of its phytotoxic analogs. Pest Manag. Sci. 2018, 74, 37–45. [Google Scholar] [CrossRef]
- Avantaggiato, G.; Solfrizzo, M.; Tosi, L.; Zazzerini, A.; Fanizzi, F.P.; Visconti, A. Isolation and characterization of phytotoxic compounds produced by Phomopsis helianthi. Nat. Toxins 1999, 7, 119–127. [Google Scholar] [CrossRef]
- Reveglia, P.; Pacetti, A.; Masi, M.; Cimmino, A.; Carella, G.; Marchi, G.; Mugnai, L.; Evidente, A. Phytotoxic metabolites produced by Diaporthe eres involved in cane blight of grapevine in Italy. Nat. Prod. Res. 2021, 35, 2872–2880. [Google Scholar] [CrossRef] [PubMed]
- Bunyapaiboonsri, T.; Yoiprommarat, S.; Srikitikulchai, P.; Srichomthong, K.; Lumyong, S. Oblongolides from the endophytic fungus Phomopsis sp. BCC 9789. J. Nat. Prod. 2010, 73, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Hussain, H.; Schulz, B.; Draeger, S.; Padula, D.; Pescitelli, G.; Van Ree, T.; Krohn, K. Three new antimicrobial metabolites from the endophytic fungus Phomopsis sp. Eur. J. Chem. 2011, 15, 2867–2873. [Google Scholar] [CrossRef]
- Evidente, A.; Rodeva, R.; Andolfi, A.; Stoyanova, Z.; Perrone, C.; Motta, A. Phytotoxic polyketides produced by Phomopsis foeniculi, a strain isolated from diseased Bulgarian fennel. Eur. J. Plant Pathol. 2011, 130, 173–182. [Google Scholar] [CrossRef]
- Tsantrizos, Y.S.; Ogilvie, K.K.; Watson, A.K. Phytotoxic metabolites of Phomopsis convolvulus, a host-specific pathogen of field bindweed. Can. J. Chem. 1992, 70, 2276–2284. [Google Scholar] [CrossRef] [Green Version]
- Ballio, A.; Bottalico, A.; Framondino, M.; Graniti, A.; Randazzo, G. Fusicoccin: Structure-phytotoxicity relationships. Phytopathol. Mediterr. 1971, 10, 26–32. [Google Scholar]
- Evidente, A.; Lanzetta, R.; Abouzeid, M.A.; Corsaro, M.M.; Mugnai, L.; Surico, G. Foeniculoxin, a new phytotoxic geranylhydroquinone from Phomopsis foeniculi. Tetrahedron 1994, 50, 10371–10378. [Google Scholar] [CrossRef]
- Turner, N.C.; Graniti, A. Fusicoccin: A fungal toxin that opens stomata. Nature 1969, 223, 1070–1071. [Google Scholar] [CrossRef]
- Culvenor, C.C.J.; Edgar, J.A.; Mackay, M.F.; Gorst-Allman, C.P.; Marasas, W.F.O.; Steyn, P.S.; Vleggaar, R.; Wessels, P.L. Structure elucidation and absolute configuration of phomopsin A, a hexapeptide mycotoxin produced by Phomopsis leptostromiformis. Tetrahedron 1989, 45, 2351–2372. [Google Scholar] [CrossRef]
- Mazars, C.; Rossignol, M.; Auriol, P.; Klaebe, A. Phomozin, a phytotoxin from Phomopsis helianthi, the causal agent of stem canker of sunflower. Phytochemistry 1990, 29, 3441–3444. [Google Scholar] [CrossRef]
- Lopes, M.J.D.S.; Dias-Filho, M.B.; Gurgel, E.S.C. Successful plant growth-promoting microbes: Inoculation methods and abiotic factors. Front. Sustain. Food Syst. 2021, 5, 606454. [Google Scholar] [CrossRef]
- Asghari, B.; Khademian, R.; Sedaghati, B. Plant growth promoting rhizobacteria (PGPR) confer drought resistance and stimulate biosynthesis of secondary metabolites in pennyroyal (Mentha pulegium L.) under water shortage condition. Sci. Hort. 2020, 263, 1–10. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Rahman, M.A.; Guo, J.; Kang, Z.; Ali Babar, M. Comparative physiological and metabolic analysis reveals a complex mechanism involved in drought tolerance in chickpea (Cicer arietinum L.) induced by PGPR and PGRs. Sci. Rep. 2019, 9, 2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poveda, J.; Eugui, D.; Abril-Urías, P.; Velasco, P. Endophytic fungi as direct plant growth promoters for sustainable agricultural production. Symbiosis 2021, 85, 1–19. [Google Scholar] [CrossRef]
- Lugtenberg, B.J.; Caradus, J.R.; Johnson, L.J. Fungal endophytes for sustainable crop production. FEMS Microbiol. Ecol. 2016, 92, fiw194. [Google Scholar] [CrossRef]
- Du, Y.C.; Kong, L.J.; Cao, L.S.; Zhang, W.; Zhu, Q.; Ma, C.Y.; Sun, K.; Dai, C.C. Endophytic Fungus Phomopsis liquidambaris Enhances Fe Absorption in Peanuts by Reducing Hydrogen Peroxide. Front. Plant Sci. 2022, 13, 872242. [Google Scholar] [CrossRef]
- Yuan, Z.L.; Dai, C.C.; Li, X.; Tian, L.S.; Wang, X.X. Extensive host range of an endophytic fungus affects the growth and physiological functions in rice (Oryza sativa L.). Symbiosis 2007, 43, 21–28. [Google Scholar]
- Zhou, J.; Huang, P.W.; Li, X.; Vaistij, F.E.; Dai, C.C. Generalist endophyte Phomopsis liquidambaris colonization of Oryza sativa L. promotes plant growth under nitrogen starvation. Plant Mol. Biol. 2022, 6, 703–715. [Google Scholar] [CrossRef]
- Aldana, B.R.V.; Arellano, J.B.; Cuesta, M.J.; Mellado-Ortega, E.; González-Blanco, V.; Zabalgogeazcoa, I. Screening fungal endophytes from a wild grass for growth promotion in tritordeum, an agricultural cereal. Plant Sci. 2021, 303, 110762. [Google Scholar] [CrossRef]
- Toghueo, R.M.; Zabalgogeazcoa, I.; Pereira, E.C.; de Aldana, B.R.V. A Diaporthe fungal endophyte from a wild grass improves growth and salinity tolerance of tritordeum and perennial ryegrass. Front. Plant Sci. 2022, 13, 896755. [Google Scholar] [CrossRef] [PubMed]
- Ważny, R.; Rozpądek, P.; Jędrzejczyk, R.J.; Domka, A.; Nosek, M.; Kidd, P.; Turnau, K. Phytohormone based biostimulant combined with plant growth promoting endophytic fungus enhances Ni phytoextraction of Noccaea goesingensis. Sci. Total Environ. 2021, 789, 147950. [Google Scholar] [CrossRef] [PubMed]
- Deepthi, A.S.; Ray, J.G. Applications of endophytic-fungal-isolates from velamen root of wild orchids in floriculture. Braz. J. Biol. 2019, 6, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Lacerda, J.W.; Siqueira, K.A.; Vasconcelos, L.G.; Bellete, B.S.; Dall’Oglio, E.L.; Sousa Junior, P.T.; Faraggi, T.M.; Vieira, L.C.; Soares, M.A.; Sampaio, O.M. Metabolomic Analysis of Combretum lanceolatum Plants Interaction with Diaporthe phaseolorum and Trichoderma spirale Endophytic Fungi through 1H-NMR. Chem. Biodivers. 2021, 18, 2100350. [Google Scholar] [CrossRef] [PubMed]
- Serrano, L.; Lacerda, J.W.; Moura, M.S.; Ali, A.; de Vasconcelos, L.G.; Junior, P.T.S.; Bellete, B.S.; Soares, M.A.; Vieira, L.C.; Sampaio, O.M. Metabolomics Analysis of C. lanceolatum Roots in the Presence of Its Endophytic Fungi. J. Braz. Chem. Soc. 2022, 18, e2100350. [Google Scholar] [CrossRef]
- da Silva Santos, S.D.S.; da Silva, A.A.D.; Polonio, J.C.; Polli, A.D.; Orlandelli, R.C.; dos Santos Oliveira, J.A.D.S.; Brandão Filho, J.U.T.; Azevedo, J.L.; Pamphile, J.A. Influence of plant growth-promoting endophytes Colletotrichum siamense and Diaporthe masirevici on tomato plants (Lycopersicon esculentum Mill.). Mycology 2022, 13, 257–270. [Google Scholar] [CrossRef]
- Ballio, A.; Michelis, M.I.; Lado, P.; Randazzo, G. Fusicoccin structure-activity relationships: Stimulation of growth by cell enlargement and promotion of seed germination. Physiol. Plant. 1981, 52, 471–475. [Google Scholar] [CrossRef]
- Marra, A.; Camoni, A.L.; Visconti, A.; Fiorillo, A.; Evident, A. The Surprising story of Fusicoccin: A wilt-inducing phytotoxin, a tool in plant physiology and a 14-3-3-targeted drug. Biomolecules 2021, 11, 1393. [Google Scholar] [CrossRef]
- Chithra, S.; Mathew, J.; Radhakrishnan, E.K. Endophytic Phomopsis sp. colonization in Oryza sativa was found to result in plant growth promotion and piperine production. Physiol. Plant. 2017, 160, 437–446. [Google Scholar]
- Bhattacharjee, S.; Roy Das, A.; Saha, A.K.; Das, P. Fungal endophytes from medicinal plants: Growth promotion in Oryza sativa L. and Cicer arietinum L. Vegetos 2019, 32, 381–386. [Google Scholar] [CrossRef]
- Yang, D.; Du, X.; Yang, Z.; Liang, Z.; Guo, Z.; Liu, Y. Transcriptomics, proteomics, and metabolomics to reveal mechanisms underlying plant secondary metabolism. Eng. Life Sci. 2014, 14, 456–466. [Google Scholar] [CrossRef]
- Xia, Y.; Liu, J.; Chen, C.; Mo, X.; Tan, Q.; He, Y.; Wang, Z.; Yin, J.; Zhou, G. The Multifunctions and Future Prospects of Endophytes and Their Metabolites in Plant Disease Management. Microorganisms 2022, 10, 1072. [Google Scholar] [CrossRef] [PubMed]
- Lubna, L.; Asaf, S.; Jan, R.; Khan, A.L.; Bilal, S.; Asif, S.; Al-Harrasi, A.; Kim, K.M. Unraveling the Genome Sequence of Plant Growth Promoting Aspergillus niger (CSR3) Provides Insight into the Synthesis of Secondary Metabolites and Its Comparative Genomics. J. Fungi 2022, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.F.M.; Esteves, A.C.; Alves, A. Marine Fungi: Opportunities and Challenges. Encyclopedia 2022, 2, 559–577. [Google Scholar] [CrossRef]
- Mohana, N.C.; Rao, H.Y.; Rakshith, D.; Mithun, P.R.; Nuthan, B.R.; Satish, S. Omics based approach for biodiscovery of microbial natural products in antibiotic resistance era. J. Genet. Eng. Biotechnol. 2018, 16, 1–8. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. AntiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Robey, M.T.; Caesar, L.K.; Drott, M.T.; Keller, N.P.; Kelleher, N.L. An interpreted atlas of biosynthetic gene clusters from 1,000 fungal genomes. Proc. Natl. Acad. Sci. USA 2021, 118, e2020230118. [Google Scholar] [CrossRef]
- Grover, S.; Lawrence, C.B. The Alternaria alternata mycotoxin alternariol suppresses lipopolysaccharide-induced inflammation. Int. J. Mol. Sci. 2017, 18, 1577. [Google Scholar] [CrossRef] [Green Version]
- Reveglia, P.; Masi, M.; Evidente, A. Melleins—Intriguing natural compounds. Biomolecules 2020, 10, 772. [Google Scholar] [CrossRef]
- Ballio, A.; Carilli, A.; Santurbano, B.; Tuttobello, L. Produzione di fusicoccina in scala pilota. Ann. Ist. Sup. Sanità 1968, 4, 317–332. [Google Scholar]
- Marrè, E. Fusicoccin: A tool in plant physiology. Annu. Rev. Plant Physiol. 1979, 30, 273–288. [Google Scholar] [CrossRef]
- Bacha, N.; Dao, H.P.; Atoui, A.; Mathieu, F.; O’Callaghan, J.; Puel, O.; Liboz, T.; Dobson, A.D.W.; Lebrihi, A. Cloning and characterization of novel methylsalicylic acid synthase gene involved in the biosynthesis of isoasperlactone and asperlactone in Aspergillus westerdijkiae. Fungal Genet. Biol. 2009, 46, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.; Balcells, M.; Sala, N.; Sanchis, V.; Canela, R. Bactericidal and fungicidal activity of Aspergillus ochraceus metabolites and some derivatives. Pestic. Sci. 1998, 53, 9–14. [Google Scholar] [CrossRef]
- Balcells, M.; Canela, R.; Coll, J.; Sanchís, V.; Torres, M. Effect of fungal metabolites and some derivatives against Tribolium castaneum (Herbst) and Nezara viridula (L.). Pest. Sci. 1995, 45, 319–323. [Google Scholar] [CrossRef]
- Giacometti, R.; Jacobi, V.; Kronberg, F.; Panagos, C.; Edison, A.S.; Zavala, J.A. Digestive activity and organic compounds of Nezara viridula watery saliva induce defensive soybean seed responses. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pointer, M.D.; Gage, M.J.; Spurgin, L.G. Tribolium beetles as a model system in evolution and ecology. Heredity 2021, 126, 869–883. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, S.Q.; Tang, H.Y.; Li, X.J.; Zhang, L.; Xiao, J.; Gao, Y.Q.; Zhang, A.L.; Gao, J.M. Potential allelopathic indole diketopiperazines produced by the plant endophytic Aspergillus fumigatus using the one strain–many compounds method. J. Agric. Food Chem. 2013, 61, 11447–11452. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xiao, J.; Sun, Q.Q.; Qin, J.C.; Pescitelli, G.; Gao, J.M. Characterization of cytochalasins from the endophytic Xylaria sp. and their biological functions. J. Agric. Food Chem. 2014, 62, 10962–10969. [Google Scholar] [CrossRef]
- Izumi, Y.; Ohtani, K.; Miyamoto, Y.; Masunaka, A.; Fukumoto, T.; Gomi, K.; Tada, Y.; Ichimura, K.; Peever, T.L.; Akimitsu, K. A polyketide synthase gene, ACRTS2, is responsible for biosynthesis of host-selective ACR-toxin in the rough lemon pathotype of Alternaria alternata. Mol. Plant Microbe Interact. 2012, 25, 1419–1429. [Google Scholar] [CrossRef] [Green Version]
- Masunaka, A.; Ohtani, K.; Peever, T.L.; Timmer, L.W.; Tsuge, T.; Yamamoto, M.; Yamamoto, H.; Akimitsu, K. An isolate of Alternaria alternata that is pathogenic to both tangerines and rough lemon and produces two host-selective toxins, ACT-and ACR-toxins. Phytopathology 2005, 95, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuge, T.; Harimoto, Y.; Akimitsu, K.; Ohtani, K.; Kodama, M.; Akagi, Y.; Egusa, M.; Yamamoto, M.; Otani, H. Hostselective toxins produced by the plant pathogenic fungus Alternaria alternata. FEMS Microbiol. Rev. 2013, 37, 44–66. [Google Scholar] [CrossRef] [PubMed]
- Gautier, C.; Pinson-Gadais, L.; Richard-Forget, F. Fusarium mycotoxins enniatins: An updated review of their occurrence, the producing Fusarium species, and the abiotic determinants of their accumulation in crop harvests. J. Agric. Food Chem. 2020, 68, 4788–4798. [Google Scholar] [CrossRef] [PubMed]
- Prosperini, A.; Berrada, H.; Ruiz, M.J.; Caloni, F.; Coccini, T.; Spicer, L.J.; Perego, M.C.; Lafranconi, A. A review of the mycotoxin enniatin B. Front. Public Health 2017, 5, 304. [Google Scholar] [CrossRef] [PubMed]
- Kleigrewe, K.; Söhnel, A.C.; Humpf, H.U. A new high-performance liquid chromatography–tandem mass spectrometry method based on dispersive solid phase extraction for the determination of the mycotoxin fusarin C in corn ears and processed corn samples. J. Agric. Food Chem 2011, 59, 10470–10476. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, W.; Lin, Y.; Lin, Y.; Tang, Z.; Wang, J.; Tao, J.; Tang, W.; Liu, W. Characterization of a carboxyl methyltransferase in Fusarium graminearum provides insights into the biosynthesis of fusarin A. Org. Biomol. Chem. 2021, 19, 6638–6643. [Google Scholar] [CrossRef]
- Ugai, T.; Minami, A.; Fujii, R.; Tanaka, M.; Oguri, H.; Gomi, K.; Oikawa, H. Heterologous expression of highly reducing polyketide synthase involved in betaenone biosynthesis. Chem. Commun. 2015, 51, 1878–1881. [Google Scholar] [CrossRef]
- Qi, J.; Jiang, L.; Zhao, P.; Chen, H.; Jia, X.; Zhao, L.; Dai, H.; Hu, J.; Liu, C.; Shim, S.H.; et al. Chaetoglobosins and azaphilones from Chaetomium globosum associated with Apostichopus japonicus. Appl. Microbiol. Biotechnol. 2020, 104, 545–1553. [Google Scholar] [CrossRef]
- Moolhuijzen, P.M.; Muria-Gonzalez, M.J.; Syme, R.; Rawlinson, C.; See, P.T.; Moffat, C.S.; Ellwood, S.R. Expansion and conservation of biosynthetic gene clusters in pathogenic Pyrenophora spp. Toxins 2020, 12, 242. [Google Scholar] [CrossRef] [Green Version]
- Kaul, S.; Sharma, T.K.; Dhar, M. “Omics” tools for better understanding the plant–endophyte interactions. Front. Plant Sci. 2016, 7, 955. [Google Scholar] [CrossRef] [Green Version]
- Balotf, S.; Wilson, R.; Tegg, R.S.; Nichols, D.S.; Wilson, C.R. Shotgun proteomics as a powerful tool for the study of the proteomes of plants, their pathogens, and plant–pathogen interactions. Proteomes 2022, 10, 5. [Google Scholar] [CrossRef]
- Yuan, J.; Zhang, W.; Sun, K.; Tang, M.-J.; Chen, P.-X.; Li, X.; Dai, C.-C. Comparative Transcriptomics and Proteomics of Atractylodes lancea in Response to Endophytic Fungus Gilmaniella sp. AL12 Reveals Regulation in Plant Metabolism. Front. Microbiol. 2019, 10, 1208. [Google Scholar] [CrossRef] [PubMed]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, D.P.; Prabha, R.; Sahu, P.K.; Gupta, V.K.; et al. Abiotic Stress Responses and Microbe-Mediated Mitigation in Plants: The Omics Strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, A.; Kataoka, R. Metabolite profiling reveals a complex response of plants to application of plant growth-promoting endophytic bacteria. Microbiol. Res. 2020, 234, 126421. [Google Scholar] [CrossRef] [PubMed]
- Spina, R.; Saliba, S.; Dupire, F.; Ptak, A.; Hehn, A.; Piutti, S.; Poinsignon, S.; Leclerc, S.; Bouguet-Bonnet, S.; Laurain-Mattar, D. Molecular identification of endophytic bacteria in Leucojum aestivum in vitro culture, NMR-based metabolomics study and LC-MS analysis leading to potential Amaryllidaceae alkaloid production. Int. J. Mol. Sci. 2021, 22, 1773. [Google Scholar] [CrossRef] [PubMed]
- Kemkuignou, B.M.; Schweizer, L.; Lambert, C.; Anoumedem, E.G.M.; Kouam, S.F.; Stadler, M.; Marin-Felix, Y. New polyketides from the liquid culture of Diaporthe breyniae sp. nov.(Diaporthales, Diaporthaceae). MycoKeys 2022, 90, 85–118. [Google Scholar] [CrossRef]
- Yedukondalu, N.; Arora, P.; Wadhwa, B.; Malik, F.A.; Vishwakarma, R.A.; Gupta, V.K.; Riyaz-Ul-Hassan, S.; Ali, A. Diapolic acid A–B from an endophytic fungus, Diaporthe terebinthifolii depicting antimicrobial and cytotoxic activity. J. Antibiot. 2017, 70, 212–215. [Google Scholar] [CrossRef]
- Mandavid, H.; Rodrigues, A.M.; Espindola, L.S.; Eparvier, V.; Stien, D. Secondary metabolites isolated from the amazonian endophytic fungus Diaporthe sp. SNB-GSS10. J. Nat. Prod. 2015, 78, 1735–1739. [Google Scholar] [CrossRef]
- Gupta, S.; Schillaci, M.; Roessner, U. Metabolomics as an emerging tool to study plant–microbe interactions. Emerg. Top. Life Sci. 2022, 6, 175–183. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Babalola, O.O. Meta-omics of endophytic microbes in agricultural biotechnology. Biocatal. Agric. Biotechnol. 2022, 42, 102332. [Google Scholar] [CrossRef]
- Cao, M.; Koulman, A.; Johnson, L.J.; Lane, G.A.; Rasmussen, S. Advanced data-mining strategies for the analysis of direct-infusion ion trap mass spectrometry data from the association of perennial ryegrass with its endophytic fungus, Neotyphodium lolii. Plant Physiol. 2008, 146, 1501–1514. [Google Scholar] [CrossRef] [PubMed]


| Compound | Strain | Host | References |
|---|---|---|---|
| 3,4-Dihydro-8-hydroxy-3,5-dimethylisocoumarin | D. eres | Hedera helix | [103] |
| (6S,7R,9R)6,7-Dihydroxy-9-propylnon-4-eno-9-lactone | Phomopsis sp. (syn. Diaporthe sp.) | Achyranthes bidentata | [100,102] |
| 4,6-dihydroxymellein | P. helianthi (syn. D. helianthi) | Helianthus annus | [104] |
| 4-Hydroxybenzaldehyde | D. eres | Vitis vinifera | [105] |
| 4-Hydroxybenzoic acid | D. eres | Vitis vinifera | [105] |
| 8-hydroxy-3,7-dimethylisochroman-1-one | D. eres | Hedera helix | [99] |
| 5-(hydroxymethyl) mellein | Phomopsis sp. (syn. Diaporthe sp.) | Musa acuminata Cistus monspeliensis | [106,107] |
| 2-(4-hydroxyphenyl)-ethanol | D. eres | Hedera helix | [99,103] |
| 5-methylmellein | Phomopsis sp. (syn. Diaporthe sp.) | Musa acuminata Cistus monspeliensis | [106,107] |
| 3-Nitropropionic acid | D. gulyae | Carthamus lanatus | [95] |
| alternariol 4,10-dimethyl | P. phragmitis (syn. D. eres) | Actinidia chinensis | [73] |
| Altersolanol A | P. foeniculi (syn. D. angelicae) | Foeniculum vulgare | [108] |
| Altersolanol J | P. foeniculi (syn. D. angelicae) | Foeniculum vulgare | [108] |
| Convolvulanic acid A | P. convolvulus (syn. D. convolvuli) | Convolvulus arvensis | [109] |
| Convolvulanic acid B | P. convolvulus (syn. D. convolvuli) | Convolvulus arvensis | [109] |
| Convolvulol | P. convolvulus (syn. D. convolvuli) | Convolvulus arvensis | [109] |
| p-Cresol | D. eres | Vitis vinifera | [105] |
| Cytochalasin H, N | Phomopsis sp. (syn. Diaporthe sp.) D. miriciae | Achyranthes bidentata, Copaifera pubiflora and Melocactus ernestii | [74,102] |
| dideacetylfusicoccin | P. amygdali (syn. D. amygdali) | - | [110] |
| Epoxycytochalasin H | Phomopsis sp. (syn. Diaporthe sp.) | Achyranthes bidentata | [102] |
| etheralternariol methyl ether | P. phragmitis (syn. D. eres) | Actinidia chinensis | [73] |
| Foeniculoxin | P. foeniculi (syn. D. angelicae) | Foeniculum vulgare | [111] |
| Fusicoccin | P. amygdali (syn. D. amygdali) | Prunus dulcis Prunus persica | [112] |
| Gulypyrone A | D. gulyae | Carthamus lanatus | [95] |
| Gulypyrone B | D. gulyae | Carthamus lanatus | [95] |
| isofusicoccin | P. amygdali (syn. D. amygdali) | - | [110] |
| Kongiidiazadione | D. kongii | Carthamus lanatus | [101] |
| monodeacetylfusicoccin | P. amygdali (syn. D. amygdali) | - | [110] |
| Nectriapyrone | D. kongii D. eres P. foeniculi (syn. D. angelicae) | Carthamus lanatus Vitis vinifera Foeniculum vulgare | [101,105] |
| Phomentrioloxin B | D. gulyae | Carthamus lanatus | [95,100] |
| Phomopsolide B | Phomopsis sp. (syn. Diaporthe sp.) | Vitis vinifera | [66] |
| Phomopsolidone A | Phomopsis sp. (syn. Diaporthe sp.) | Vitis vinifera | [66] |
| Phomopsolidone B | Phomopsis sp. (syn. Diaporthe sp.) | Vitis vinifera | [66] |
| Phomopsin A | P. leptostromiformis (syn. D. toxica) | Lupinus sp. | [113] |
| Phomozin | P. helianthi (syn. D. helianthi) | Helianthus annus | [114] |
| α-pyrone convolvulopyrone | P. convolvulus (syn. D. convolvuli) | Convolvulus arvensis | [109] |
| Sydowinin A | Phomopsis sp. (syn. Diaporthe sp.) | Vitis vinifera | [66] |
| Sydowinol | Phomopsis sp. (syn. Diaporthe sp.) | Vitis vinifera | [66] |
| Tyrosol | D. eres | Vitis vinifera Hedera helix | [103,105] |
| Host Plant | Endophyte | Benefits | References |
|---|---|---|---|
| Acampe praemorsa | D. eucalyptorum | Increase the fresh-weight and enhance the growth of ornamental orchids (Dendrobium sp.) | [126] |
| Combretum lanceolatum | D. phaseolorum | Improve the biosynthesis of primary metabolites involved in Combretum lanceolatum self-defence | [127,128] |
| Festuca rubra | Diaporthe sp. | Improve nutrient uptake, growth, and salinity tolerance of Lolium perenne and the hybrid from Triticum durum × Hordeum | [124] |
| Helianthus tuberosus | D. phaseolorum | Enhance the growth and increases chlorophyll content of sunchoke plants (Helianthus tuberosus) | [11] |
| Justicia brandegeana | D. masirevicii | Growth-promoting effects on tomato plants and suppression of F. oxysporum symptoms in this host | [129] |
| Lactuca sativa | P. amygdali (syn. D. amygdali) | Cell enlargement, cotyledon growth and seed germination | [130,131] |
| Noccaea goesingensis | D. eres | Improve the biomass Noccaea goesingensis and increase nickel accumulation. Potentially used as biofertilizer. | [124] |
| Oryza sativa | P. liquidambaris (syn. D. liquidambaris) | Enhance growth of rice plants under nitrogen-deficient conditions, and induces rice resistance to rice spikelet rot disease caused by Fusarium graminearum | [121] |
| Piper nigrum | Phomopsis sp. (syn. Diaporthe sp.) | Enhance growth of rice plants due to the production of gibberellins and indole acetic acid | [132] |
| Terminalia arjuna Phlogacanthus thyrsiflorus | D. phaseolorum | Increase root length and enhance plant growth of Cicer arietinum | [133] |
| Triticum durum × Hordeum chilense | Diaporthe sp. | Increase concentrations of calcium, magnesium, sulphur, iron, and boron, thus increasing root and shoot biomass of T. durum × H. chilense | [122] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hilário, S.; Gonçalves, M.F.M. Endophytic Diaporthe as Promising Leads for the Development of Biopesticides and Biofertilizers for a Sustainable Agriculture. Microorganisms 2022, 10, 2453. https://doi.org/10.3390/microorganisms10122453
Hilário S, Gonçalves MFM. Endophytic Diaporthe as Promising Leads for the Development of Biopesticides and Biofertilizers for a Sustainable Agriculture. Microorganisms. 2022; 10(12):2453. https://doi.org/10.3390/microorganisms10122453
Chicago/Turabian StyleHilário, Sandra, and Micael F. M. Gonçalves. 2022. "Endophytic Diaporthe as Promising Leads for the Development of Biopesticides and Biofertilizers for a Sustainable Agriculture" Microorganisms 10, no. 12: 2453. https://doi.org/10.3390/microorganisms10122453
APA StyleHilário, S., & Gonçalves, M. F. M. (2022). Endophytic Diaporthe as Promising Leads for the Development of Biopesticides and Biofertilizers for a Sustainable Agriculture. Microorganisms, 10(12), 2453. https://doi.org/10.3390/microorganisms10122453