

TLR4/CD14/MD2 Revealed as the Limited Toll-like Receptor Complex for Chlamydia trachomatis-Induced NF-κB Signaling
 , ,
, ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Strains and Ct Culture
2.2. Cell Lines and Culture Conditions
2.3. Reagents and Antibodies
2.4. Stimulation of Reporter Cells
2.5. Analysis by Flow Cytometry
2.6. Infectivity Assay of Reporter Cells using Imaging Flow Cytometry
3. Results
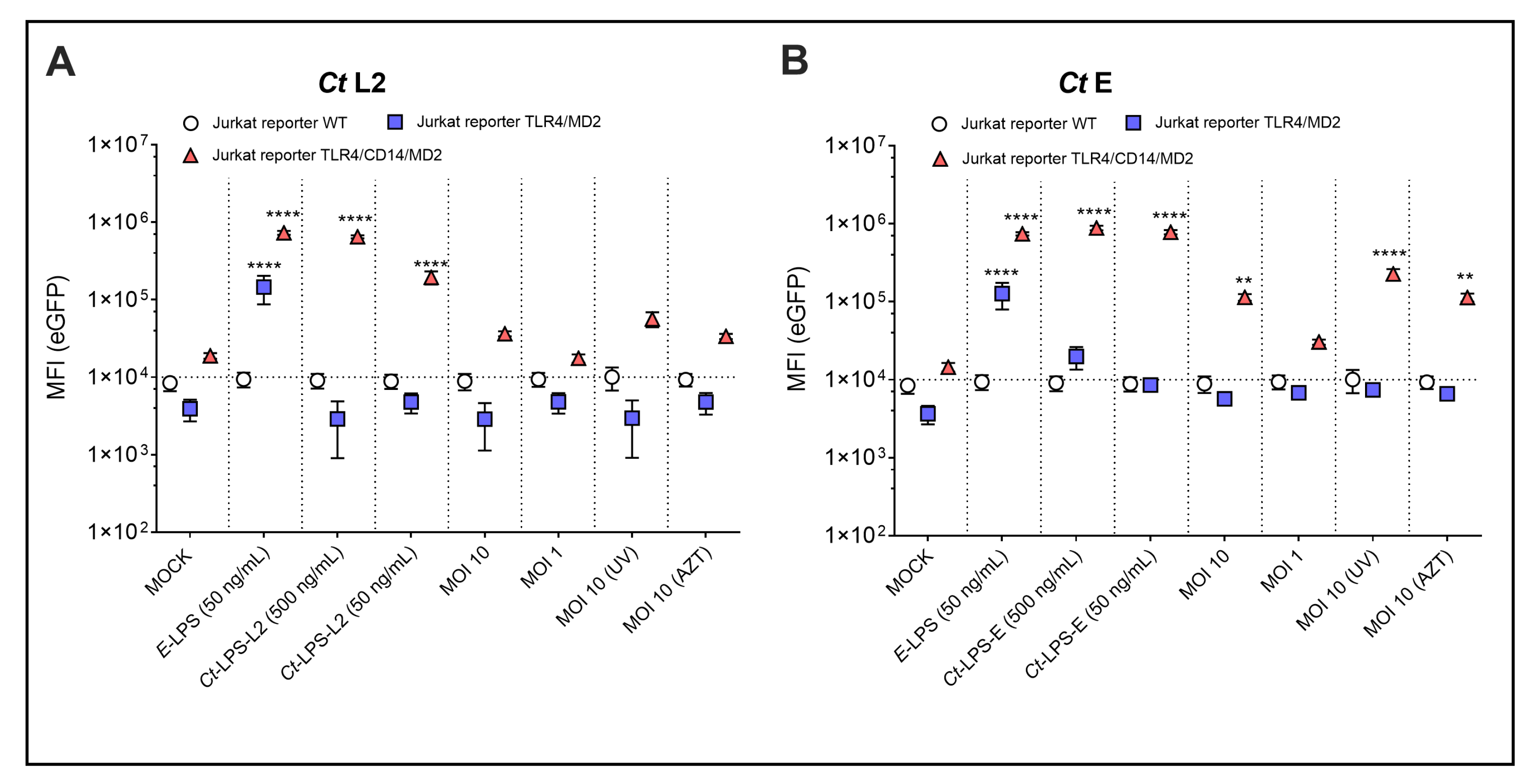
3.1. Identification and Analysis of TLRs for Ct Recognition
3.2. Identification and Analysis of TLRs for Ct Recognition
3.3. Simultaneous Analysis of Infectivity and Signal Induction by Ct
3.4. Identification and Enumeration of Ct-Infected Cells by Imaging Flow Cytometry
3.5. The Role of the TLR4/CD14/MD2 Complex in Ct Uptake
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Unemo, M.; Bradshaw, C.S.; Hocking, J.S.; de Vries, H.J.C.; Francis, S.C.; Mabey, D.; Marrazzo, J.M.; Sonder, G.J.B.; Schwebke, J.R.; Hoornenborg, E.; et al. Sexually transmitted infections: Challenges ahead. Lancet. Infect. Dis. 2017, 17, e235–e279. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Guidelines for the Treatment of Chlamydia trachomatis; World Health Organization: Geneva, Switzerland, 2016; p. 47. [Google Scholar]
- Wang, S.P.; Grayston, J.T. Three new serovars of Chlamydia trachomatis: Da, Ia, and L2a. J. Infect. Dis. 1991, 163, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Jordan, S.; Nelson, D.; Geisler, W. Chlamydia trachomatis Infections. In Chlamydia Biology: From Genome to Disease; Caister Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Mabey, D.; Peeling, R.W. Lymphogranuloma venereum. Sex. Transm. Infect. 2002, 78, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Gervassi, A.; Alderson, M.R.; Suchland, R.; Maisonneuve, J.F.; Grabstein, K.H.; Probst, P. Differential regulation of inflammatory cytokine secretion by human dendritic cells upon Chlamydia trachomatis infection. Infect. Immun. 2004, 72, 7231–7239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, B.; Njau, F.; Thalmann, J.; Haller, H.; Wagner, A.D. Differential infection outcome of Chlamydia trachomatis in human blood monocytes and monocyte-derived dendritic cells. BMC Microbiol. 2014, 14, 209. [Google Scholar] [CrossRef]
- Ward, H.; Alexander, S.; Carder, C.; Dean, G.; French, P.; Ivens, D.; Ling, C.; Paul, J.; Tong, W.; White, J.; et al. The prevalence of lymphogranuloma venereum infection in men who have sex with men: Results of a multicentre case finding study. Sex. Transm. Infect. 2009, 85, 173–175. [Google Scholar] [CrossRef] [Green Version]
- Geisler, W.M.; Chow, J.M.; Schachter, J.; McCormack, W.M. Pelvic examination findings and Chlamydia trachomatis infection in asymptomatic young women screened with a nucleic acid amplification test. Sex. Transm. Dis. 2007, 34, 335–338. [Google Scholar] [CrossRef]
- Darville, T.; Hiltke, T.J. Pathogenesis of Genital Tract Disease Due to Chlamydia trachomatis. J. Infect. Dis. 2010, 201, 114–125. [Google Scholar] [CrossRef] [Green Version]
- Hafner, L.M. Pathogenesis of fallopian tube damage caused by Chlamydia trachomatis infections. Contraception 2015, 92, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Darville, T.; O’Neill, J.M.; Andrews, C.W.J.; Nagarajan, U.M.; Stahl, L.; Ojcius, D.M. Toll-like receptor-2, but not Toll-like receptor-4, is essential for development of oviduct pathology in chlamydial genital tract infection. J. Immunol. 2003, 171, 6187–6197. [Google Scholar] [CrossRef]
- Erridge, C.; Pridmore, A.; Eley, A.; Stewart, J.; Poxton, I.R. Lipopolysaccharides of Bacteroides fragilis, Chlamydia trachomatis and Pseudomonas aeruginosa signal via Toll-like receptor 2. J. Med. Microbiol. 2004, 53, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Prebeck, S.; Kirschning, C.; Dürr, S.; da Costa, C.; Donath, B.; Brand, K.; Redecke, V.; Wagner, H.; Miethke, T. Predominant role of toll-like receptor 2 versus 4 in Chlamydia pneumoniae-induced activation of dendritic cells. J. Immunol. 2001, 167, 3316–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, J.K.; Kim, S.J.; Rah, S.H.; Kang, J.I.; Jung, H.E.; Lee, D.; Lee, H.K.; Lee, J.O.; Park, B.S.; Yoon, T.Y.; et al. Reconstruction of LPS Transfer Cascade Reveals Structural Determinants within LBP, CD14, and TLR4-MD2 for Efficient LPS Recognition and Transfer. Immunity 2017, 46, 38–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heine, H.; Müller-Loennies, S.; Brade, L.; Lindner, B.; Brade, H. Endotoxic activity and chemical structure of lipopolysaccharides from Chlamydia trachomatis serotypes E and L2 and Chlamydophila psittaci 6BC. Eur. J. Biochem. 2003, 270, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Heine, H.; Gronow, S.; Zamyatina, A.; Kosma, P.; Brade, H. Investigation on the agonistic and antagonistic biological activities of synthetic Chlamydia lipid A and its use in in vitro enzymatic assays. J. Endotoxin Res. 2007, 13, 126–132. [Google Scholar] [CrossRef]
- Scior, T.; Alexander, C.; Zaehringer, U. Reviewing and identifying amino acids of human, murine, canine and equine TLR4/MD-2 receptor complexes conferring endotoxic innate immunity activation by LPS/lipid A, or antagonistic effects by Eritoran, in contrast to species-dependent modulation by l. Comput. Struct. Biotechnol. J. 2013, 5, e201302012. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Wang, F.; Lu, Z.; Ying, H.; Zhang, H.; Ding, W.; Wang, C.; Shi, L. MAPK kinase 3 potentiates Chlamydia HSP60-induced inflammatory response through distinct activation of NF-κB. J. Immunol. 2013, 191, 386–394. [Google Scholar] [CrossRef] [Green Version]
- Massari, P.; Toussi, D.N.; Tifrea, D.F.; de la Maza, L.M. Toll-like receptor 2-dependent activity of native major outer membrane protein proteosomes of Chlamydia trachomatis. Infect. Immun. 2013, 81, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Wantia, N.; Rodriguez, N.; Cirl, C.; Ertl, T.; Dürr, S.; Layland, L.E.; Wagner, H.; Miethke, T. Toll-like receptors 2 and 4 regulate the frequency of IFNγ-producing CD4+ T-cells during pulmonary infection with Chlamydia pneumoniae. PLoS ONE 2011, 6, e26101. [Google Scholar] [CrossRef]
- Nosratababadi, R.; Bagheri, V.; Zare-Bidaki, M.; Hakimi, H.; Zainodini, N.; Kazemi Arababadi, M. Toll like receptor 4: An important molecule in recognition and induction of appropriate immune responses against Chlamydia infection. Comp. Immunol. Microbiol. Infect. Dis. 2017, 51, 27–33. [Google Scholar] [CrossRef]
- Prebeck, S.; Brade, H.; Kirschning, C.J.; Prazeres Da Costa, C.; Dürr, S.; Wagner, H.; Miethke, T. The Gram-negative bacterium Chlamydia trachomatis L2stimulates tumor necrosis factor secretion by innate immune cells independently of its endotoxin. Microbes Infect. 2003, 5, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Scidmore, M.A. Cultivation and Laboratory Maintenance of Chlamydia trachomatis. In Current Protocols in Microbiology; Wiley Online Library: Hoboken, NJ, USA, 2005; pp. 11A.1.1–11A.1.25, Chapter 1A; ISBN 9780471729259. [Google Scholar]
- Klasinc, R.; Reiter, M.; Digruber, A.; Tschulenk, W.; Walter, I.; Kirschner, A.; Spittler, A.; Stockinger, H. A Novel Flow Cytometric Approach for the Quantification and Quality Control of Chlamydia trachomatis Preparations. Pathogens 2021, 10, 1617. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int. J. Cancer 1980, 26, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Jutz, S.; Leitner, J.; Schmetterer, K.; Doel-Perez, I.; Majdic, O.; Grabmeier-Pfistershammer, K.; Paster, W.; Huppa, J.B.; Steinberger, P. Assessment of costimulation and coinhibition in a triple parameter T cell reporter line: Simultaneous measurement of NF-κB, NFAT and AP-1. J. Immunol. Methods 2016, 430, 10–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battin, C.; Hennig, A.; Mayrhofer, P.; Kunert, R.; Zlabinger, G.J.; Steinberger, P.; Paster, W. A human monocytic NF-κB fluorescent reporter cell line for detection of microbial contaminants in biological samples. PLoS ONE 2017, 12, e0178220. [Google Scholar] [CrossRef] [Green Version]
- Radakovics, K.; Battin, C.; Leitner, J.; Geiselhart, S.; Paster, W.; Stöckl, J.; Hoffmann-Sommergruber, K.; Steinberger, P. A Highly Sensitive Cell-Based TLR Reporter Platform for the Specific Detection of Bacterial TLR Ligands. Front. Immunol. 2022, 12, 817604. [Google Scholar] [CrossRef]
- Cossarizza, A.; Chang, H.-D.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef] [Green Version]
- Haridas, V.; Ranjbar, S.; Vorobjev, I.A.; Goldfeld, A.E.; Barteneva, N.S. Imaging flow cytometry analysis of intracellular pathogens. Methods 2017, 112, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Autenrieth, S.E.; Warnke, P.; Wabnitz, G.H.; Lucero Estrada, C.; Pasquevich, K.A.; Drechsler, D.; Günter, M.; Hochweller, K.; Novakovic, A.; Beer-Hammer, S.; et al. Depletion of Dendritic Cells Enhances Innate Anti-Bacterial Host Defense through Modulation of Phagocyte Homeostasis. PLoS Pathog. 2012, 8, e1002552. [Google Scholar] [CrossRef]
- Kubo, T.; Ishida, K.; Matsuo, J.; Nakamura, S.; Hayashi, Y.; Sakai, H.; Yoshida, M.; Takahashi, K.; Hirai, I.; Yamamoto, Y.; et al. Chlamydia trachomatis serovar L2 infection model using human lymphoid Jurkat cells. Microb. Pathog. 2012, 53, 1–11. [Google Scholar] [CrossRef]
- Faris, R.; Andersen, S.E.; McCullough, A.; Gourronc, F.; Klingelhutz, A.J.; Weber, M.M. Chlamydia trachomatis Serovars Drive Differential Production of Proinflammatory Cytokines and Chemokines Depending on the Type of Cell Infected. Front. Cell. Infect. Microbiol. 2019, 9, 399. [Google Scholar] [CrossRef] [PubMed]
- Finethy, R.; Coers, J. Sensing the enemy, containing the threat: Cell-autonomous immunity to Chlamydia trachomatis. FEMS Microbiol. Rev. 2016, 40, 875–893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.; Jain, P.; Bettahi, I.; Pal, S.; Tifrea, D.; de la Maza, L.M. A TLR2 agonist is a more effective adjuvant for a Chlamydia major outer membrane protein vaccine than ligands to other TLR and NOD receptors. Vaccine 2011, 29, 6641–6649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, C.M.; AbdelRahman, Y.M.; Green, E.; Darville, H.K.; Saira, K.; Smith, B.; Darville, T.; Scurlock, A.M.; Meyer, C.R.; Belland, R.J. Toll-Like Receptor 2 Activation by Chlamydia trachomatis Is Plasmid Dependent, and Plasmid-Responsive Chromosomal Loci Are Coordinately Regulated in Response to Glucose Limitation by C. trachomatis but Not by C. muridarum. Infect. Immun. 2011, 79, 1044–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagarajan, U.M.; Ojcius, D.M.; Stahl, L.; Rank, R.G.; Darville, T. Chlamydia trachomatis induces expression of IFN-gamma-inducible protein 10 and IFN-beta independent of TLR2 and TLR4, but largely dependent on MyD88. J. Immunol. 2005, 175, 450–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gracey, E.; Lin, A.; Akram, A.; Chiu, B.; Inman, R.D. Intracellular Survival and Persistence of Chlamydia muridarum Is Determined by Macrophage Polarization. PLoS ONE 2013, 8, e69421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchacher, T.; Wiesinger-Mayr, H.; Vierlinger, K.; Rüger, B.M.; Stanek, G.; Fischer, M.B.; Weber, V. Human blood monocytes support persistence, but not replication of the intracellular pathogen C. pneumoniae. BMC Immunol. 2014, 15, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buchacher, T.; Ohradanova-Repic, A.; Stockinger, H.; Fischer, M.B.; Weber, V. M2 Polarization of Human Macrophages Favors Survival of the Intracellular Pathogen Chlamydia pneumoniae. PLoS ONE 2015, 10, e0143593. [Google Scholar] [CrossRef] [Green Version]
- Yeung, A.T.Y.; Hale, C.; Lee, A.H.; Gill, E.E.; Bushell, W.; Parry-Smith, D.; Goulding, D.; Pickard, D.; Roumeliotis, T.; Choudhary, J.; et al. Exploiting induced pluripotent stem cell-derived macrophages to unravel host factors influencing Chlamydia trachomatis pathogenesis. Nat. Commun. 2017, 8, 15013. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Briones, M.; Janice, C.; Lei, L.; Patton, J.M.; Ma, L.; McClarty, G.; Caldwell, H.D. Chlamydia trachomatis Lipopolysaccharide Evades the Canonical and Noncanonical Inflammatory Pathways To Subvert Innate Immunity. MBio 2019, 10, e00595-19. [Google Scholar] [CrossRef]
- Rosadini, C.V.; Kagan, J.C. Early innate immune responses to bacterial LPS. Curr. Opin. Immunol. 2017, 44, 14–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rund, S.; Lindner, B.; Brade, H.; Holst, O. Structural Analysis of the Lipopolysaccharide from Chlamydia trachomatis Serotype L2. J. Biol. Chem. 1999, 274, 16819–16824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingalls, R.R.; Rice, P.A.; Qureshi, N.; Takayama, K.; Lin, J.S.; Golenbock, D.T. The inflammatory cytokine response to Chlamydia trachomatis infection is endotoxin mediated. Infect. Immun. 1995, 63, 3125–3130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morré, S.A.; Murillo, L.S.; Bruggeman, C.A.; Peña, A.S. The role that the functional Asp299Gly polymorphism in the toll-like receptor-4 gene plays in susceptibility to Chlamydia trachomatis-associated tubal infertility. J. Infect. Dis. 2003, 187, 341–343. [Google Scholar] [CrossRef] [Green Version]
- Zanoni, I.; Ostuni, R.; Marek, L.R.; Barresi, S.; Barbalat, R.; Barton, G.M.; Granucci, F.; Kagan, J.C. CD14 controls the LPS-induced endocytosis of Toll-like receptor 4. Cell 2011, 147, 868–880. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klasinc, R.; Battin, C.; Paster, W.; Reiter, M.; Schatzlmaier, P.; Rhein, P.; Spittler, A.; Steinberger, P.; Stockinger, H. TLR4/CD14/MD2 Revealed as the Limited Toll-like Receptor Complex for Chlamydia trachomatis-Induced NF-κB Signaling. Microorganisms 2022, 10, 2489. https://doi.org/10.3390/microorganisms10122489
Klasinc R, Battin C, Paster W, Reiter M, Schatzlmaier P, Rhein P, Spittler A, Steinberger P, Stockinger H. TLR4/CD14/MD2 Revealed as the Limited Toll-like Receptor Complex for Chlamydia trachomatis-Induced NF-κB Signaling. Microorganisms. 2022; 10(12):2489. https://doi.org/10.3390/microorganisms10122489
Chicago/Turabian StyleKlasinc, Romana, Claire Battin, Wolfgang Paster, Michael Reiter, Philipp Schatzlmaier, Peter Rhein, Andreas Spittler, Peter Steinberger, and Hannes Stockinger. 2022. "TLR4/CD14/MD2 Revealed as the Limited Toll-like Receptor Complex for Chlamydia trachomatis-Induced NF-κB Signaling" Microorganisms 10, no. 12: 2489. https://doi.org/10.3390/microorganisms10122489
APA StyleKlasinc, R., Battin, C., Paster, W., Reiter, M., Schatzlmaier, P., Rhein, P., Spittler, A., Steinberger, P., & Stockinger, H. (2022). TLR4/CD14/MD2 Revealed as the Limited Toll-like Receptor Complex for Chlamydia trachomatis-Induced NF-κB Signaling. Microorganisms, 10(12), 2489. https://doi.org/10.3390/microorganisms10122489