
Integron Functionality and Genome Innovation: An Update on the Subtle and Smart Strategy of Integrase and Gene Cassette Expression Regulation


Abstract
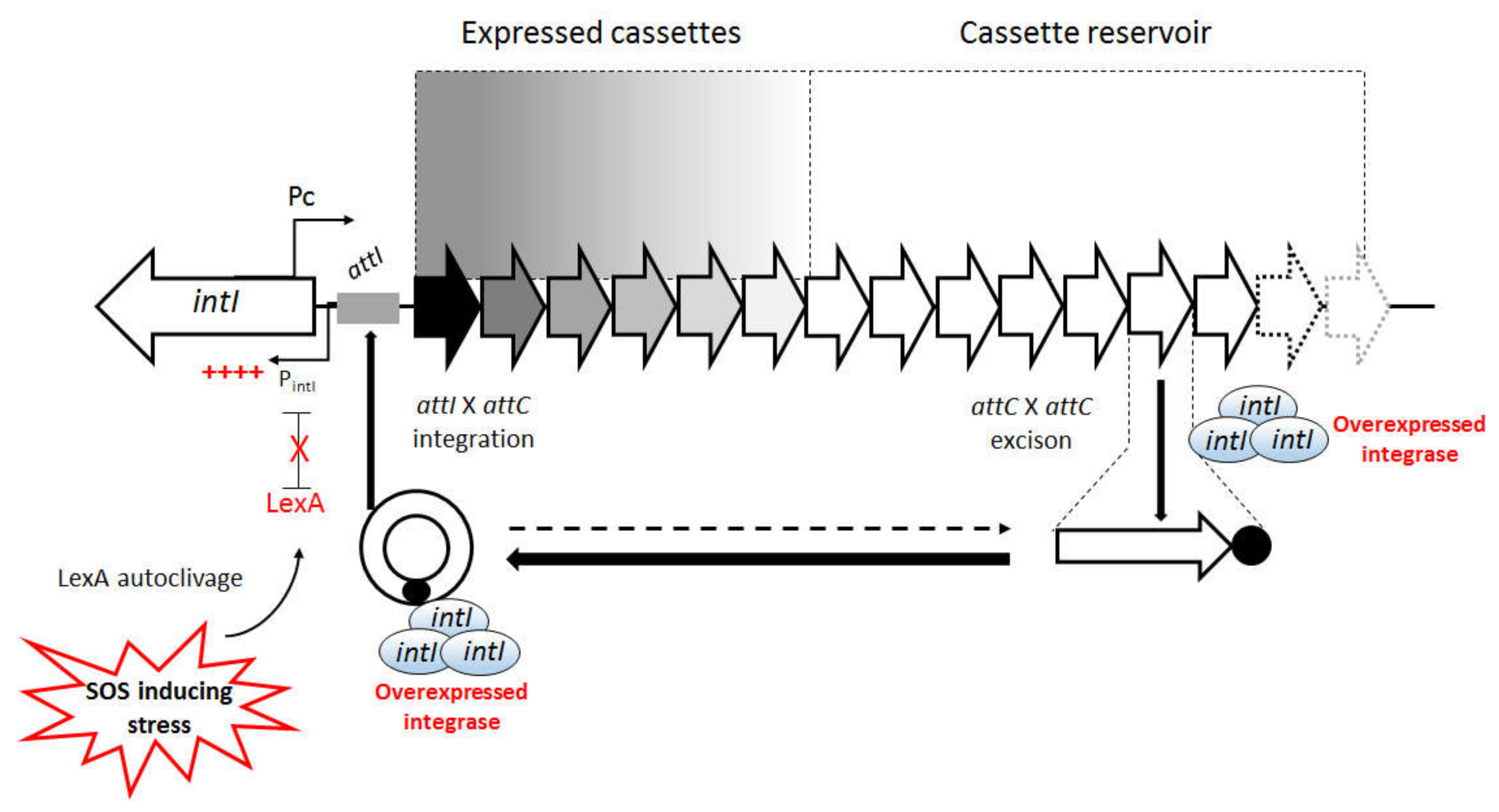
:1. Introduction
Integron Functionality Is Directly Linked with Adaptation on Demand
2. Transcriptional Regulation in Integrons
2.1. The Role of Pc Promoters on Gene Cassette Transcription
2.1.1. Pc Promoters in Class 1 Integrons

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pc Variant 1 | –35 Sequence | Spacer Region (bp) | –10 Sequence | Transcription Strength 2 | Reference |
|---|---|---|---|---|---|
| Pc variants with polymorphisms inside –35 and –10 regions | |||||
| PcS | TTGACA | 17 | TAAACT | 1240 | [2,3,26,28] |
| PcH2 | TTGACA | 17 | TAAGCT | 840 | [26,27,28] |
| PcH1 | TGGACA | 17 | TAAACT | 220 | [2,3,26,28] |
| PcSS | TTGATA | 17 | TAAACT | 100 | [26,38] |
| PcPUO | TCGACA | 17 | TAAACT | 60 | [26,35] |
| PcW | TGGACA | 17 | TAAGCT | 40 | [2,3,26,28] |
| PcIn42 | TTGGCA | 17 | TAAACT | 40 | [26,36] |
| PcIn116 | TTGACA | 17 | TGAACT | 10 | [26,37] |
| Pc variants with polymorphisms upstream the –10 region | |||||
| PcSTGN–10 | TTGACA | 17 | (TGN)TAAACT | 1160 | [26] |
| PcH2TGN–10 | TTGACA | 17 | (TGN)TAAGCT | 860 | [26] |
| PcWTGN–10 | TGGACA | 17 | (TGN)TAAGCT | 720 | [26,39] |
| PcH1TTN–10 | TGGACA | 17 | (TTN)TAAACT | 375 | [26] |
| PcWTAN–10 | TGGACA | 17 | (TAN)TAAGCT | undetectable | [26] |
| Pc variants combined with P2 | |||||
| PcS + P2 | TTGACA TTGTTA | 17 17 | TAAACT TACAGT | 1000 | [26,28] |
| PcWTGN–10 + P2 | TGGACA TTGTTA | 17 17 | (TGN)TAAGCT TACAGT | 620 | [26] |
| PcH1 + P2 | TGGACA TTGTTA | 17 17 | TAAACT TACAGT | 380 | [26,28] |
| PcW + P2 | TGGACA TTGTTA | 17 17 | TAAGCT TACAGT | 340 | [2,3,26,28] |
| Pc variants combined with P2 variants | |||||
| PcWTGN–10 + P2m1 | TGGACA TTGTTA | 17 17 | (TGN)TAAGCT GACAGT | NP | [26] |
| PcH1 + P2m1 | TGGACA TTGTTA | 17 17 | TAAACT GACAGT | NP | [26] |
| PcW + P2m2 | TGGACA TTGTTA | 17 17 | TAAGCT TACACA | NP | [26] |
| PcW + P2m3 | TGGACA TTGTTA | 17 17 | TAAGCT TACAAT | 720 | [29] |
2.1.2. Pc Promoter in Class 2 Integrons
2.1.3. Pc Promoter in Class 3 Integrons
2.1.4. Pc Promoter in Chromosomal Integrons
2.2. The Role of Gene Cassette Position Effect on Transcription
2.3. The Role of Cassette-Borne Promoter on Gene Cassette Transcription
2.4. Transcription of Gene Cassettes Independent of Pc and Cassette-Borne Promoters
2.4.1. Insertion Sequences Elements
2.4.2. Group II Intron Elements
2.4.3. ORF-Less Cassettes
2.4.4. Fused Gene Cassettes
2.5. Regulation of Integrase Transcription in MI and CI by Stress Response Mechanisms
2.5.1. The Role of SOS Response
2.5.2. The Role CRP-cAMP Catabolite Repression Pathway and Environmental Physicochemical Conditions
2.5.3. The Role of the Stringent Response
2.5.4. The Role of Other Stress Response Regulators
2.6. Interplay between Integrase and Gene Cassette Expression in Class 1 Integrons
2.6.1. Integrase Activity Is Linked and Inversely Correlated to Gene Cassette Expression
2.6.2. Transcription Interference between PintI1 and Cassette Promoters in Class 1 Integrons
2.6.3. Global Regulators Concomitantly Control the Activity of PintI1 and Cassette Promoters in Class 1 Integrons
2.7. Integrase Expression in Class 2 Integrons
2.8. Integrase Expression in Class 3 Integrons
3. Post-Transcriptional and Translational Regulation in Integrons
3.1. The Role of attI1 Site on Integron-Associated Gene Cassette Translation
3.2. The Role of attC Site on Integron-Associated Gene Cassette Translation
3.3. Other Aspects Influencing Translation of Downstream Integron-Associated Gene Cassettes
3.4. Control of Gene Cassette Expression by Translation Attenuation
3.5. Integron-Encoded Regulatory RNA
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hall, R.M.; Collis, C.M. Mobile gene cassettes and integrons: Capture and spread of genes by site-specific recombination. Mol. Microbiol. 1995, 15, 593–600. [Google Scholar] [CrossRef]
- Collis, C.M.; Hall, R.M. Expression of antibiotic resistance genes in the integrated cassettes of integrons. Antimicrob. Agents Chemother. 1995, 39, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévesque, C.; Brassard, S.; Lapointe, J.; Roy, P.H. Diversity and relative strength of tandem promoters for the antibiotic-resistance genes of several integrons. Gene 1994, 142, 49–54. [Google Scholar] [CrossRef]
- Cambray, G.; Guerout, A.M.; Mazel, D. Integrons. Annu. Rev. Genet. 2010, 44, 141–166. [Google Scholar] [CrossRef] [PubMed]
- Recchia, G.D.; Hall, R.M. Gene cassettes: A new class of mobile element. Microbiology 1995, 141, 3015–3027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stokes, H.W.; O’Gorman, D.B.; Recchia, G.D.; Parsekhian, M.; Hall, R.M. Structure and function of 59-base element recombination sites associated with mobile gene cassettes. Mol. Microbiol. 1997, 26, 731–745. [Google Scholar] [CrossRef] [PubMed]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Mazel, D.; Dychinco, B.; Webb, V.A.; Davies, J. A distinctive class of integron in the Vibrio cholerae genome. Science 1998, 280, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Rowe-Magnus, D.A.; Guerout, A.M.; Mazel, D. Bacterial resistance evolution by recruitment of super-integron gene cassettes. Mol. Microbiol. 2002, 43, 1657–1669. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, E.L.; Dos Santos Freitas, F.; Vieira, V.V.; Vicente, A.C. New qnr gene cassettes associated with superintegron repeats in Vibrio cholerae O1. Emerg. Infect. Dis. 2008, 14, 1129–1131. [Google Scholar] [CrossRef]
- Fonseca, E.L.; Vicente, A.C.P. Epidemiology of qnrVC alleles and emergence out of the Vibrionaceae family. J. Med. Microbiol. 2013, 62, 1628–1630. [Google Scholar] [CrossRef] [Green Version]
- Labbate, M.; Case, R.J.; Stokes, H.W. The integron/gene cassette system: An active player in bacterial adaptation. Methods Mol. Biol. 2009, 532, 103–125. [Google Scholar] [CrossRef]
- Escudero, J.A.; Loot, C.; Nivina, A.; Mazel, D. The Integron: Adaptation on Demand. Microbiol. Spectr. 2015, 3, MDNA3-0019-2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaly, T.M.; Geoghegan, J.L.; Tetu, S.G.; Gillings, M.R. The Peril and Promise of Integrons: Beyond Antibiotic Resistance. Trends Microbiol. 2020, 28, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Depardieu, F.; Podglajen, I.; Leclercq, R.; Collatz, E.; Courvalin, P. Modes and modulations of antibiotic resistance gene expression. Clin. Microbiol. Rev. 2007, 20, 79–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacotte, Y.; Ploy, M.C.; Raherison, S. Class 1 integrons are low-cost structures in Escherichia coli. ISME J. 2017, 11, 1535–1544. [Google Scholar] [CrossRef] [Green Version]
- Guerin, E.; Cambray, G.; Sanchez-Alberola, N.; Campoy, S.; Erill, I.; Da Re, S.; Gonzalez-Zorn, B.; Barbé, J.; Ploy, M.C.; Mazel, D. The SOS response controls integron recombination. Science 2009, 324, 1034. [Google Scholar] [CrossRef] [Green Version]
- Baharoglu, Z.; Bikard, D.; Mazel, D. Conjugative DNA transfer induces the bacterial SOS response and promotes antibiotic resistance development through integron activation. PLoS Genet. 2010, 6, e1001165. [Google Scholar] [CrossRef] [Green Version]
- Cambray, G.; Sanchez-Alberola, N.; Campoy, S.; Guerin, É.; Da Re, S.; González-Zorn, B.; Ploy, M.C.; Barbé, J.; Mazel, D.; Erill, I. Prevalence of SOS-mediated control of integron integrase expression as an adaptive trait of chromosomal and mobile integrons. Mob. DNA 2011, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Cagle, C.A.; Shearer, J.E.S.; Summers, A.O. Regulation of the integrase and cassette promoters of the class 1 integron by nucleoid-associated proteins. Microbiology 2011, 157, 2841–2853. [Google Scholar] [CrossRef]
- Baharoglu, Z.; Krin, E.; Mazel, D. Connecting environment and genome plasticity in the characterization of transformation-induced SOS regulation and carbon catabolite control of the Vibrio cholerae integron integrase. J. Bacteriol. 2012, 194, 1659–1667. [Google Scholar] [CrossRef] [Green Version]
- Krin, E.; Cambray, G.; Mazel, D. The superintegron integrase and the cassette promoters are co-regulated in Vibrio cholerae. PLoS ONE 2014, 9, e91194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collis, C.M.; Hall, R.M. Gene cassettes from the insert region of integrons are excised as covalently closed circles. Mol. Microbiol. 1992, 6, 2875–2885. [Google Scholar] [CrossRef] [PubMed]
- Gillings, M.R. Integrons: Past, present, and future. Microbiol. Mol. Biol. Rev. 2014, 78, 257–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nivina, A.; Escudero, J.A.; Vit, C.; Mazel, D.; Loot, C. Efficiency of integron cassette insertion in correct orientation is ensured by the interplay of the three unpaired features of attC recombination sites. Nucleic Acids Res. 2016, 44, 7792–7803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jové, T.; Da Re, S.; Denis, F.; Mazel, D.; Ploy, M.C. Inverse correlation between promoter strength and excision activity in class 1 integrons. PLoS Genet. 2010, 6, e1000793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunny, K.L.; Hall, R.M.; Stokes, H.W. New mobile gene cassettes containing an aminoglycoside resistance gene, aacA7, and a chloramphenicol resistance gene, catB3, in an integron in pBWH301. Antimicrob. Agents Chemother. 1995, 39, 686–693. [Google Scholar] [CrossRef] [Green Version]
- Papagiannitsis, C.C.; Tzouvelekis, L.S.; Miriagou, V. Relative strengths of the class 1 integron promoter hybrid 2 and the combinations of strong and hybrid 1 with an active p2 promoter. Antimicrob. Agents Chemother. 2009, 53, 277–280. [Google Scholar] [CrossRef] [Green Version]
- Vinué, L.; Jové, T.; Torres, C.; Ploy, M.C. Diversity of class 1 integron gene cassette Pc promoter variants in clinical Escherichia coli strains and description of a new P2 promoter variant. Int. J. Antimicrob. Agents. 2011, 38, 526–529. [Google Scholar] [CrossRef] [Green Version]
- Schimitz, F.J.; Hafner, D.; Geisel, R.; Follmann, P.; Kirschke, C.; Verhoef, J.; Kohrer, K.; Fluit, A.C. Increased prevalence of class I integrons in Escherichia coli, Klebsiella species, and Enterobacter species isolates over a 7-year period in a German university hospital. J. Clin. Microbiol. 2001, 39, 3724–3726. [Google Scholar] [CrossRef] [Green Version]
- Lindstedt, B.A.; Heir, E.; Nygård, I.; Kapperud, G. Characterization of class I integrons in clinical strains of Salmonella enterica subsp. enterica serovars Typhimurium and Enteritidis from Norwegian hospitals. J. Med. Microbiol. 2003, 52, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Cocchi, S.; Grasselli, E.; Gutacker, M.; Benagli, C.; Convert, M.; Piffaretti, J.-C. Distribution and characterization of integrons in Escherichia coli strains of animal and human origin. FEMS Immunol. Med. Microbiol. 2007, 50, 126–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura, A.; Jové, T.; Ploy, M.C.; Henriques, I.; Correia, A. Diversity of gene cassette promoters in class 1 integrons from wastewater environments. Appl. Environ. Microbiol. 2012, 78, 5413–5416. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Jiang, X.; Li, M.; Li, G.; Hu, Q.; Lu, H.; Chen, G.; Zhou, Y.; Lu, Y. Diversity of gene cassette promoter variants of class 1 integrons in uropathogenic Escherichia coli. Curr. Microbiol. 2013, 67, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Riccio, M.L.; Franceschini, N.; Boschi, L.; Caravelli, B.; Cornaglia, G.; Fontana, R.; Amicosante, G.; Rossolini, G.M. Characterization of the metallo-beta-lactamase determinant of Acinetobacter baumannii AC-54/97 reveals the existence of bla(IMP) allelic variants carried by gene cassettes of different phylogeny. Antimicrob. Agents Chemother. 2000, 44, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Houang, E.T.; Chu, Y.W.; Lo, W.S.; Chu, K.Y.; Cheng, A.F. Epidemiology of rifampin ADP-ribosyltransferase (arr-2) and metallo-beta-lactamase (blaIMP-4) gene cassettes in class 1 integrons in Acinetobacter strains isolated from blood cultures in 1997 to 2000. Antimicrob. Agents Chemother. 2003, 47, 1382–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Power, P.; Galleni, M.; Di Conza, J.; Ayala, J.A.; Gutkind, G. Description of In116, the first blaCTX-M-2-containing complex class 1 integron found in Morganella morganii isolates from Buenos Aires, Argentina. J. Antimicrob. Chemother. 2005, 55, 461–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brízio, A.; Conceição, T.; Pimentel, M.; da Silva, G.; Duarte, A. High-level expression of IMP-5 carbapenemase owing to point mutation in the -35 promoter region of class 1 integron among Pseudomonas aeruginosa clinical isolates. Int. J. Antimicrob. Agents 2006, 27, 27–31. [Google Scholar] [CrossRef]
- Nesvera, J.; Hochmannová, J.; Pátek, M. An integron of class 1 is present on the plasmid pCG4 from gram-positive bacterium Corynebacterium glutamicum. FEMS Microbiol Lett. 1998, 169, 391–395. [Google Scholar] [CrossRef] [Green Version]
- Soufi, L.; Sáenz, Y.; Vinué, L.; Abbassi, M.S.; Hammami, S.; Torres, C. Characterization of Pc promoter variants of class 1 integrons in Escherichia coli isolates from poultry meat. Foodborne Pathog. Dis. 2013, 10, 1075–1077. [Google Scholar] [CrossRef]
- Naas, T.; Mikami, Y.; Imai, T.; Poirel, L.; Nordmann, P. Characterization of In53, a class 1 plasmid- and composite transposon-located integron of Escherichia coli which carries an unusual array of gene cassettes. J. Bacteriol. 2001, 183, 235–249. [Google Scholar] [CrossRef] [Green Version]
- Tseng, C.S.; Yen, Y.C.; Chang, C.C.; Hsu, Y.M. Polymorphism of gene cassette promoter variants of class 1 integron harbored in S. Choleraesuis and Typhimurium isolated from Taiwan. Biomedicine 2014, 4, 20. [Google Scholar] [CrossRef] [PubMed]
- Couvé-Deacon, E.; Jové, T.; Afouda, P.; Barraud, O.; Tilloy, V.; Scaon, E.; Hervé, B.; Burucoa, C.; Kempf, M.; Marcos, J.Y.; et al. Class 1 integrons in Acinetobacter baumannii: A weak expression of gene cassettes to counterbalance the lack of LexA-driven integrase repression. Int. J. Antimicrob. Agents 2019, 53, 491–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Wang, X.; Kong, N.; Cao, M.; Zhang, L.; Wei, Q.; Liu, W. Polymorphisms of Gene Cassette Promoters of the Class 1 Integron in Clinical Proteus Isolates. Front. Microbiol. 2019, 10, 790. [Google Scholar] [CrossRef]
- Hall, R.M.; Stokes, H.W. Integrons: Novel DNA elements which capture genes by site-specific recombination. Genetica 1993, 90, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Hansson, K.; Sundström, L.; Pelletier, A.; Roy, P.H. IntI2 integron integrase in Tn7. J. Bacteriol. 2002, 184, 1712–1721. [Google Scholar] [CrossRef] [Green Version]
- Biskri, L.; Mazel, D. Erythromycin esterase gene ere(A) is located in a functional gene cassette in an unusual class 2 integron. Antimicrob. Agents Chemother. 2003, 47, 3326–3331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jové, T.; Da Re, S.; Tabesse, A.; Gassama-Sow, A.; Ploy, M.C. Gene Expression in Class 2 Integrons Is SOS-Independent and Involves Two Pc Promoters. Front. Microbiol. 2017, 8, 1499. [Google Scholar] [CrossRef] [Green Version]
- da Fonseca, É.L.; dos Santos Freitas, F.; Vicente, A.C. Pc promoter from class 2 integrons and the cassette transcription pattern it evokes. J. Antimicrob. Chemother. 2011, 66, 797–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barlow, R.S.; Gobius, K.S. Diverse class 2 integrons in bacteria from beef cattle sources. J. Antimicrob. Chemother. 2006, 58, 1133–1138. [Google Scholar] [CrossRef] [Green Version]
- Márquez, C.; Labbate, M.; Ingold, A.J.; Roy Chowdhury, P.; Ramírez, M.S.; Centrón, D.; Borthagaray, G.; Stokes, H.W. Recovery of a functional class 2 integron from an Escherichia coli strain mediating a urinary tract infection. Antimicrob. Agents Chemother. 2008, 52, 4153–4154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Minguela, C.M.; Apajalahti, J.H.; Chai, B.; Cole, J.R.; Tiedje, J.M. Worldwide prevalence of class 2 integrases outside the clinical setting is associated with human impact. Appl. Environ. Microbiol. 2009, 75, 5100–5110. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Hu, Q.; Li, S.; Lu, H.; Chen, G.; Shen, B.; Zhang, P.; Zhou, Y. A novel functional class 2 integron in clinical Proteus mirabilis isolates. J. Antimicrob. Chemother. 2014, 69, 973–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakawa, Y.; Murakami, M.; Suzuki, K.; Ito, H.; Wacharotayankun, R.; Ohsuka, S.; Kato, N.; Ohta, M. A novel integron-like element carrying the metallo-beta-lactamase gene blaIMP. Antimicrob. Agents Chemother. 1995, 39, 1612–1615. [Google Scholar] [CrossRef] [Green Version]
- Collis, C.M.; Kim, M.J.; Partridge, S.R.; Stokes, H.W.; Hall, R.M. Characterization of the class 3 integron and the site-specific recombination system it determines. J. Bacteriol. 2002, 184, 3017–3026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correia, M.; Boavida, F.; Grosso, F.; Salgado, M.J.; Lito, L.M.; Cristino, J.M.; Mendo, S.; Duarte, A. Molecular characterization of a new class 3 integron in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2003, 47, 2838–2843. [Google Scholar] [CrossRef] [Green Version]
- Coleman, N.V.; Holmes, A.J. The native Pseudomonas stutzeri strain Q chromosomal integron can capture and express cassette-associated genes. Microbiology 2005, 151, 1853–1864. [Google Scholar] [CrossRef] [Green Version]
- Yachie, N.; Arakawa, K.; Tomita, M. On the interplay of gene positioning and the role of rho-independent terminators in Escherichia coli. FEBS Lett. 2006, 580, 6909–6914. [Google Scholar] [CrossRef] [Green Version]
- Jacquier, H.; Zaoui, C.; Sanson-le Pors, M.J.; Mazel, D.; Berçot, B. Translation regulation of integrons gene cassette expression by the attC sites. Mol. Microbiol. 2009, 72, 1475–1486. [Google Scholar] [CrossRef]
- Lovett, P.S. Translational attenuation as the regulator of inducible cat genes. J. Bacteriol. 1990, 172, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Coyne, S.; Guigon, G.; Courvalin, P.; Périchon, B. Screening and Quantification of the Expression of Antibiotic Resistance Genes in Acinetobacter baumannii with a Microarray. Antimicrob. Agents Chemother. 2010, 54, 333–340. [Google Scholar] [CrossRef] [Green Version]
- Bissonnette, L.; Champetier, S.; Buisson, J.P.; Roy, P.H. Characterization of the nonenzymatic chloramphenicol resistance (cmlA) gene of the In4 integron of Tn1696: Similarity of the product to transmembrane transport proteins. J. Bacteriol. 1991, 173, 4493–4502. [Google Scholar] [CrossRef] [Green Version]
- Ploy, M.C.; Courvalin, P.; Lambert, T. Characterization of In40 of Enterobacter aerogenes BM2688, a class 1 integron with two new gene cassettes, cmlA2 and qacF. Antimicrob. Agents Chemother. 1998, 42, 2557–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Lambert, T.; Türkoglü, S.; Ronco, E.; Gaillard, J.; Nordmann, P. Characterization of Class 1 integrons from Pseudomonas aeruginosa that contain the bla(VIM-2) carbapenem-hydrolyzing beta-lactamase gene and of two novel aminoglycoside resistance gene cassettes. Antimicrob. Agents Chemother. 2001, 45, 546–552. [Google Scholar] [CrossRef] [Green Version]
- Quiroga, M.P.; Andres, P.; Petroni, A.; Soler Bistué, A.J.; Guerriero, L.; Vargas, L.J.; Zorreguieta, A.; Tokumoto, M.; Quiroga, C.; Tolmasky, M.E.; et al. Complex class 1 integrons with diverse variable regions, including aac(6′)-Ib-cr, and a novel allele, qnrB10, associated with ISCR1 in clinical enterobacterial isolates from Argentina. Antimicrob. Agents Chemother. 2007, 51, 4466–4470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partridge, S.R.; Tsafnat, G.; Coiera, E.; Iredell, J.R. Gene cassettes and cassette arrays in mobile resistance integrons. FEMS Microbiol. Rev. 2009, 33, 757–784. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Moreno, M.O.; Estepa, V.; Sáenz, Y.; Cortell-Ortolá, M.; Fort-Gallifa, I.; Ruiz, J.; Torres, C. Intrahospitalary dissemination of Klebsiella pneumoniae carrying bla(DHA-1) and qnrB4 genes within a novel complex class 1 integron. Diagn. Microbiol. Infect. Dis. 2012, 73, 210–211. [Google Scholar] [CrossRef]
- Raherison, S.; Jove, T.; Gaschet, M.; Pinault, E.; Tabesse, A.; Torres, C.; Ploy, M.C. Expression of the aac(6′)-Ib-cr Gene in Class 1 Integrons. Antimicrob. Agents Chemother. 2017, 61, e02704-16. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, E.L.; Vicente, A.C. Functional characterization of a Cassette-specific promoter in the class 1 integron-associated qnrVC1 gene. Antimicrob. Agents Chemother. 2012, 56, 3392–3394. [Google Scholar] [CrossRef] [Green Version]
- Xia, R.; Guo, X.; Zhang, Y.; Xu, H. qnrVC-like gene located in a novel complex class 1 integron harboring the ISCR1 element in an Aeromonas punctata strain from an aquatic environment in Shandong Province, China. Antimicrob. Agents Chemother. 2010, 54, 3471–3474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bado, I.; Papa-Ezdra, R.; Cordeiro, N.; Outeda, M.; Caiata, L.; García-Fulgueiras, V.; Seija, V.; Vignoli, R. Detection of qnrVC6, within a new genetic context, in an NDM-1-producing Citrobacter freundii clinical isolate from Uruguay. J. Glob. Antimicrob. Resist. 2018, 14, 95–98. [Google Scholar] [CrossRef]
- Rowe-Magnus, D.A.; Guerout, A.M.; Biskri, L.; Bouige, P.; Mazel, D. Comparative analysis of superintegrons: Engineering extensive genetic diversity in the Vibrionaceae. Genome Res. 2003, 13, 428–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szekeres, S.; Dauti, M.; Wilde, C.; Mazel, D.; Rowe-Magnus, D.A. Chromosomal toxin-antitoxin loci can diminish large-scale genome reductions in the absence of selection. Mol. Microbiol. 2007, 63, 1588–1605. [Google Scholar] [CrossRef] [PubMed]
- Guérout, A.M.; Iqbal, N.; Mine, N.; Ducos-Galand, M.; Van Melderen, L.; Mazel, D. Characterization of the phd-doc and ccd toxin-antitoxin cassettes from Vibrio superintegrons. J. Bacteriol. 2013, 195, 2270–2283. [Google Scholar] [CrossRef] [Green Version]
- Tansirichaiya, S.; Mullany, P.; Roberts, A.P. Promoter activity of ORF-less gene cassettes isolated from the oral metagenome. Sci. Rep. 2019, 9, 8388. [Google Scholar] [CrossRef] [Green Version]
- Michael, C.A.; Labbate, M. Gene cassette transcription in a large integron-associated array. BMC Genet. 2010, 11, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirel, L.; Decousser, J.W.; Nordmann, P. Insertion sequence ISEcp1B is involved in expression and mobilization of a bla(CTX-M) beta-lactamase gene. Antimicrob. Agents Chemother. 2003, 47, 2938–2945. [Google Scholar] [CrossRef] [Green Version]
- Vandecraen, J.; Chandler, M.; Aertsen, A.; Houdt, R.V. The impact of insertion sequences on bacterial genome plasticity and adaptability. Crit. Rev. Microbiol. 2017, 43, 709–730. [Google Scholar] [CrossRef]
- Goussard, S.; Sougakoff, W.; Mabilat, C.; Bauernfeind, A.; Courvalin, P. An IS1-like element is responsible for high-level synthesis of extended-spectrum beta-lactamase TEM-6 in Enterobacteriaceae. J. Gen. Microbiol. 1991, 137, 2681–2687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, A.E.; Skurray, R.A.; Firth, N. An IS257-derived hybrid promoter directs transcription of a tetA(K) tetracycline resistance gene in the Staphylococcus aureus chromosomal mec region. J. Bacteriol. 2000, 182, 3345–3352. [Google Scholar] [CrossRef] [Green Version]
- Mussi, M.A.; Limansky, A.S.; Viale, A.M. Acquisition of resistance to carbapenems in multidrug-resistant clinical strains of Acinetobacter baumannii: Natural insertional inactivation of a gene encoding a member of a novel family of beta-barrel outer membrane proteins. Antimicrob. Agents Chemother. 2005, 49, 1432–1440. [Google Scholar] [CrossRef] [Green Version]
- Toleman, M.A.; Bennett, P.M.; Walsh, T.R. ISCR elements: Novel gene-capturing systems of the 21st century? Microbiol. Mol. Biol. Rev. 2006, 70, 296–316. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Martínez, J.M.; Velasco, C.; García, I.; Cano, M.E.; Martínez-Martínez, L.; Pascual, A. Characterisation of integrons containing the plasmid-mediated quinolone resistance gene qnrA1 in Klebsiella pneumoniae. Int. J. Antimicrob. Agents. 2007, 29, 705–709. [Google Scholar] [CrossRef]
- Wachino, J.; Yoshida, H.; Yamane, K.; Suzuki, S.; Matsui, M.; Yamagishi, T.; Tsutsui, A.; Konda, T.; Shibayama, K.; Arakawa, Y. SMB-1, a novel subclass B3 metallo-beta-lactamase, associated with ISCR1 and a class 1 integron, from a carbapenem-resistant Serratia marcescens clinical isolate. Antimicrob. Agents Chemother. 2011, 55, 5143–5149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Conza, J.A.; Gutkind, G.O.; Mollerach, M.E.; Ayala, J.A. Transcriptional analysis of the bla(CTX-M-2) gene in Salmonella enterica serovar Infantis. Antimicrob. Agents Chemother. 2005, 49, 3014–3017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mammeri, H.; Poirel, L.; Nordmann, P. Bactericidal activity of fluoroquinolones against plasmid-mediated QnrA-producing Escherichia coli. Clin. Microbiol. Infect. 2005, 11, 1048–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lallement, C.; Pasternak, C.; Ploy, M.C.; Jové, T. The Role of ISCR1-Borne POUT Promoters in the Expression of Antibiotic Resistance Genes. Front. Microbiol. 2018, 9, 2579. [Google Scholar] [CrossRef]
- Arduino, S.M.; Roy, P.H.; Jacoby, G.A.; Orman, B.E.; Pineiro, S.A.; Centron, D. blaCTX-M-2 is located in an unusual class 1 integron (In35) which includes Orf513. Antimicrob. Agents Chemother. 2002, 46, 2303–2306. [Google Scholar] [CrossRef] [Green Version]
- Kamruzzaman, M.; Patterson, J.D.; Shoma, S.; Ginn, A.N.; Partridge, S.R.; Iredell, J.R. Relative Strengths of Promoters Provided by Common Mobile Genetic Elements Associated with Resistance Gene Expression in Gram-Negative Bacteria. Antimicrob. Agents Chemother. 2015, 59, 5088–5091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centrón, D.; Roy, P.H. Presence of a group II intron in a multiresistant Serratia marcescens strain that harbors three integrons and a novel gene fusion. Antimicrob. Agents Chemother. 2002, 46, 1402–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambowitz, A.M.; Zimmerly, S. Group II introns: Mobile ribozymes that invade DNA. Cold Spring Harb. Perspect. Biol. 2011, 3, a003616. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, C.; Centrón, D. Using genomic data to determine the diversity and distribution of target site motifs recognized by class C-attC group II introns. J. Mol. Evol. 2009, 68, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Léon, G.; Quiroga, C.; Centrón, D.; Roy, P.H. Diversity and strength of internal outward-oriented promoters in group IIC-attC introns. Nucleic Acids Res. 2010, 38, 8196–8207. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Shi, L.; Yang, W.; Li, L.; Yamasaki, S. New array of aacA4-catB3-dfrA1 gene cassettes and a noncoding cassette from a class-1-integron-positive clinical strain of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2006, 50, 2278–2279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tansirichaiya, S.; Rahman, M.A.; Antepowicz, A.; Mullany, P.; Roberts, A.P. Detection of novel integrons in the metagenome of human saliva. PLoS ONE 2016, 11, e0157605. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-L.; Tang, D.-J.; Jiang, R.-P.; He, Y.-Q.; Jiang, B.-L.; Lu, G.-T.; Tang, J.-L. sRNA-Xcc1, an integron-encoded transposon- and plasmid-transferred trans-acting sRNA, is under the positive control of the key virulence regulators HrpG and HrpX of Xanthomonas campestris pathovar campestris. RNA Biol. 2011, 8, 947–953. [Google Scholar] [CrossRef] [Green Version]
- Tolmasky, M.E.; Crosa, J.H. Genetic organization of antibiotic resistance genes (aac(6′)-Ib, aadA, and oxa9) in the multiresistance transposon Tn1331. Plasmid 1993, 29, 31–40. [Google Scholar] [CrossRef]
- Fonseca, E.L.; Vicente, A.C. Polycistronic transcription of fused cassettes and identification of translation initiation signals in an unusual gene cassette array from Pseudomonas aeruginosa. F1000Research 2015, 2, 99. [Google Scholar] [CrossRef]
- Baharoglu, Z.; Mazel, D. SOS, the formidable strategy of bacteria against aggressions. FEMS Microbiol. Rev. 2014, 38, 1126–1145. [Google Scholar] [CrossRef] [Green Version]
- Erill, I.; Campoy, S.; Barbé, J. Aeons of distress: An evolutionary perspective on the bacterial SOS response. FEMS Microbiol. Rev. 2007, 31, 637–656. [Google Scholar] [CrossRef] [Green Version]
- Guérin, E.; Jové, T.; Tabesse, A.; Mazel, D.; Ploy, M.C. High-level gene cassette transcription prevents integrase expression in class 1 integrons. J. Bacteriol. 2011, 193, 5675–5682. [Google Scholar] [CrossRef]
- Beaber, J.W.; Hochhut, B.; Waldor, M.K. SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature 2004, 427, 72–74. [Google Scholar] [CrossRef]
- Miller, C.; Thomsen, L.E.; Gaggero, C.; Mosseri, R.; Ingmer, H.; Cohen, S.N. SOS response induction by beta-lactams and bacterial defense against antibiotic lethality. Science 2004, 305, 1629–1631. [Google Scholar] [CrossRef] [PubMed]
- Hocquet, D.; Llanes, C.; Thouverez, M.; Kulasekara, H.D.; Bertrand, X.; Plésiat, P.; Mazel, D.; Miller, S.I. Evidence for induction of integron-based antibiotic resistance by the SOS response in a clinical setting. PLoS Pathog. 2012, 8, e1002778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Re, S.; Garnier, F.; Guérin, E.; Campoy, S.; Denis, F.; Ploy, M.C. The SOS response promotes qnrB quinolone-resistance determinant expression. EMBO Rep. 2009, 10, 929–933. [Google Scholar] [CrossRef] [Green Version]
- Cameron, A.D.; Redfield, R.J. Non-canonical CRP sites control competence regulons in Escherichia coli and many other gamma-proteobacteria. Nucleic Acids Res. 2006, 34, 6001–6014. [Google Scholar] [CrossRef] [Green Version]
- Strugeon, E.; Tilloy, V.; Ploy, M.C.; Da Re, S. The Stringent Response Promotes Antibiotic Resistance Dissemination by Regulating Integron Integrase Expression in Biofilms. mBio 2016, 7, e00868-16. [Google Scholar] [CrossRef] [Green Version]
- Madsen, J.S.; Burmølle, M.; Hansen, L.H.; Sørensen, S.J. The interconnection between biofilm formation and horizontal gene transfer. FEMS Immunol. Med. Microbiol. 2012, 65, 183–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernier, S.P.; Lebeaux, D.; De Francesco, A.S.; Valomon, A.; Soubigou, G.; Coppée, J.Y.; Ghigo, J.M.; Beloin, C. Starvation, together with the SOS response, mediates high biofilm-specific tolerance to the fluoroquinolone ofloxacin. PLoS Genet. 2013, 9, e1003144. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, A. A polyphosphate-lon protease complex in the adaptation of Escherichia coli to amino acid starvation. Biosci. Biotechnol. Biochem. 2006, 70, 325–331. [Google Scholar] [CrossRef]
- Srivatsan, A.; Wang, J.D. Control of bacterial transcription, translation and replication by (p)ppGpp. Curr. Opin. Microbiol. 2008, 11, 100–105. [Google Scholar] [CrossRef]
- Kojic, M.; Venturi, V. Regulation of rpoS gene expression in Pseudomonas: Involvement of a TetR family regulator. J. Bacteriol. 2001, 183, 3712–3720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novovic, K.D.; Malesevic, M.J.; Filipic, B.V.; Mirkovic, N.L.; Miljkovic, M.S.; Kojic, M.O.; Jovčić, B.U. PsrA Regulator Connects Cell Physiology and Class 1 Integron Integrase Gene Expression through the Regulation of lexA Gene Expression in Pseudomonas spp. Curr. Microbiol. 2019, 76, 320–328. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, D.; Demarre, G.; Bouvier, M.; Mazel, D.; Gopaul, D.N. Structural basis for broad DNA-specificity in integron recombination. Nature 2006, 440, 1157–1162. [Google Scholar] [CrossRef]
- Demarre, G.; Frumerie, C.; Gopaul, D.N.; Mazel, D. Identification of key structural determinants of the IntI1 integron integrase that influence attC X attI1 recombination efficiency. Nucleic Acids Res. 2007, 35, 6475–6489. [Google Scholar] [CrossRef]
- Barraud, O.; Ploy, M.C. Diversity of Class 1 Integron Gene Cassette Rearrangements Selected under Antibiotic Pressure. J. Bacteriol. 2015, 197, 2171–2178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larouche, A.; Roy, P.H. Effect of attC structure on cassette excision by integron integrases. Mob. DNA 2011, 2, 3. [Google Scholar] [CrossRef] [Green Version]
- Robinson, A.; Brzoska, A.J.; Turner, K.M.; Withers, R.; Harry, E.J.; Lewis, P.J.; Dixon, N.E. Essential biological processes of an emerging pathogen: DNA replication, transcription, and cell division in Acinetobacter spp. Microbiol. Mol. Biol. Rev. 2010, 74, 273–297. [Google Scholar] [CrossRef] [Green Version]
- Shearwin, K.E.; Callen, B.P.; Egan, J.B. Transcriptional interference—A crash course. Trends Genet. 2005, 21, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Dorman, C.J. Nucleoid-associated proteins and bacterial physiology. Adv. Appl. Microbiol. 2009, 67, 47–64. [Google Scholar] [CrossRef]
- McCarthy, J.E.; Gualerzi, C. Translational control of prokaryotic gene expression. Trends Genet. 1990, 6, 78–85. [Google Scholar] [CrossRef]
- Hanau-Berçot, B.; Podglajen, I.; Casin, I.; Collatz, E. An intrinsic control element for translational initiation in class 1 integrons. Mol. Microbiol. 2002, 44, 119–130. [Google Scholar] [CrossRef]
- Papagiannitsis, C.C.; Tzouvelekis, L.S.; Tzelepi, E.; Miriagou, V. attI1-Located Small Open Reading Frames ORF-17 and ORF-11 in a Class 1 Integron Affect Expression of a Gene Cassette Possessing a Canonical Shine-Dalgarno Sequence. Antimicrob. Agents Chemother. 2017, 61, e02070-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souque, C.; Escudero, J.A.; MacLean, R.C. Integron activity accelerates the evolution of antibiotic resistance. eLife 2021, 10, e62474. [Google Scholar] [CrossRef]
- Casin, I.; Hanau-Berçot, B.; Podglajen, I.; Vahaboglu, H.; Collatz, E. Salmonella enterica serovar Typhimurium bla(PER-1)-carrying plasmid pSTI1 encodes an extended-spectrum aminoglycoside 6′-N-acetyltransferase of type Ib. Antimicrob. Agents Chemother. 2003, 47, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Harrod, R.; Rogers, E.J.; Lovett, P.S. Anti-peptidyl transferase leader peptides of attenuation-regulated chloramphenicol-resistance genes. Proc. Natl. Acad. Sci. USA 1994, 91, 5612–5616. [Google Scholar] [CrossRef] [Green Version]
- Escudero, J.A.; Loot, C.; Mazel, D. Integrons as adaptive devices. In Molecular Mechanisms of Microbial Evolution, 1st ed.; Rampelotto, P.H., Ed.; Springer: Cham, Switzerland, 2018; pp. 199–239. [Google Scholar] [CrossRef]
- Brandis, G.; Gockel, J.; Garoff, L.; Guy, L.; Hughes, D. Expression of the qepA1 gene is induced under antibiotic exposure. J. Antimicrob. Chemother. 2021, 76, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- Stokes, H.W.; Hall, R.M. Sequence analysis of the inducible chloramphenicol resistance determinant in the Tn1696 integron suggests regulation by translational attenuation. Plasmid 1991, 26, 10–19. [Google Scholar] [CrossRef]
- Jia, X.; Zhang, J.; Sun, W.; He, W.; Jiang, H.; Chen, D.; Murchie, A.I. Riboswitch regulation of aminoglycoside resistance acetyl and adenyl transferases. Cell 2013, 153, 1419–1420. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Zhang, X.; Zhang, J.; Jia, X.; Zhang, J.; Sun, W.; Jiang, H.; Chen, D.; Murchie, A.I. Riboswitch control of induction of aminoglycoside resistance acetyl and adenyl-transferases. RNA Biol. 2013, 10, 1266–1273. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Murchie, A.I. An aminoglycoside sensing riboswitch controls the expression of aminoglycoside resistance acetyltransferase and adenyltransferases. Biochim. Biophys. Acta 2014, 1839, 951–958. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, G.; Zhang, X.; Chang, Y.; Wang, S.; He, W.; Sun, W.; Chen, D.; Murchie, A.I.H. Aminoglycoside riboswitch control of the expression of integron associated aminoglycoside resistance adenyltransferases. Virulence 2020, 11, 1432–1442. [Google Scholar] [CrossRef] [PubMed]
- Anderson, S.E.; Chin, C.Y.; Weiss, D.S.; Rather, P.N. Copy Number of an Integron-Encoded Antibiotic Resistance Locus Regulates a Virulence and Opacity Switch in Acinetobacter baumannii AB5075. mBio 2020, 11, e02338-20. [Google Scholar] [CrossRef] [PubMed]
- Roth, A.; Breaker, R.R. Integron attI1 sites, not riboswitches, associate with antibiotic resistance genes. Cell 2013, 153, 1417–1418. [Google Scholar] [CrossRef] [Green Version]
- Carrier, M.C.; Lalaouna, D.; Massé, E. Broadening the Definition of Bacterial Small RNAs: Characteristics and Mechanisms of Action. Annu. Rev. Microbiol. 2018, 72, 141–161. [Google Scholar] [CrossRef] [PubMed]
- Chin, C.Y.; Tipton, K.A.; Farokhyfar, M.; Burd, E.M.; Weiss, D.S.; Rather, P.N. A high-frequency phenotypic switch links bacterial virulence and environmental survival in Acinetobacter baumannii. Nat. Microbiol. 2018, 3, 563–569. [Google Scholar] [CrossRef]












| Integron Features | Resistance/Mobile Integrons (Example of Class 1 Integron) | Chromosomal Integrons (Example of IntVchA) |
|---|---|---|
| Genetic context | Mobile genetic elements (transposons, plasmids, ICEs) | Chromosome |
| Integrase gene | intI1 | intIA (intI4) |
| Number of gene cassettes | Dozens | Hundreds |
| Nature of gene cassettes | Antibiotic resistance | Unknown functions, metabolic functions, antibiotic resistance (less frequent) |
| Cassette recombination sites | attC (variable in sequence and size) | VCR (~120-bp repeat) |
| Pc2 Promoter Variants | –35 Sequence | Spacer Region (bp) | –10 Sequence | Position | Transcriptional Activity | Reference |
|---|---|---|---|---|---|---|
| Pc2A Pc2A-V2 | TTTTAA TTTTAA | 17 17 | TAAAAT TAAAGT | attI2 | YES | [48,49] |
| Pc2B PC2B-V2 | TTGTAT TTATAT | 16 16 | TTTAAT TTTAAT | attI2 | YES | [48] |
| Pc2C | GTGACA | 19 | TAAAAT | attI2 | YES | [48] |
| Pc2D | TTGCAA | 18 | TATTCT | attI2 | NO | [48] |
| Pc2E | TGGCTA | 17 | (TGN)TAAGCT | intI2 | NO | [48] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fonseca, É.L.; Vicente, A.C. Integron Functionality and Genome Innovation: An Update on the Subtle and Smart Strategy of Integrase and Gene Cassette Expression Regulation. Microorganisms 2022, 10, 224. https://doi.org/10.3390/microorganisms10020224
Fonseca ÉL, Vicente AC. Integron Functionality and Genome Innovation: An Update on the Subtle and Smart Strategy of Integrase and Gene Cassette Expression Regulation. Microorganisms. 2022; 10(2):224. https://doi.org/10.3390/microorganisms10020224
Chicago/Turabian StyleFonseca, Érica L., and Ana Carolina Vicente. 2022. "Integron Functionality and Genome Innovation: An Update on the Subtle and Smart Strategy of Integrase and Gene Cassette Expression Regulation" Microorganisms 10, no. 2: 224. https://doi.org/10.3390/microorganisms10020224
APA StyleFonseca, É. L., & Vicente, A. C. (2022). Integron Functionality and Genome Innovation: An Update on the Subtle and Smart Strategy of Integrase and Gene Cassette Expression Regulation. Microorganisms, 10(2), 224. https://doi.org/10.3390/microorganisms10020224