
Trypanosomes of the Trypanosoma theileri Group: Phylogeny and New Potential Vectors

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Trapping and Processing, and Sampling of Deer Blood
2.2. Dissection and Cultivation
2.3. PCR Detection of Kinetoplastids, Sequencing
2.4. Prevalence of T. Theileri
2.5. Experimental Infections and Prediuresis Experiments
2.6. Light and Scanning Electron Microscopy
2.7. Phylogenetic Analysis
3. Results
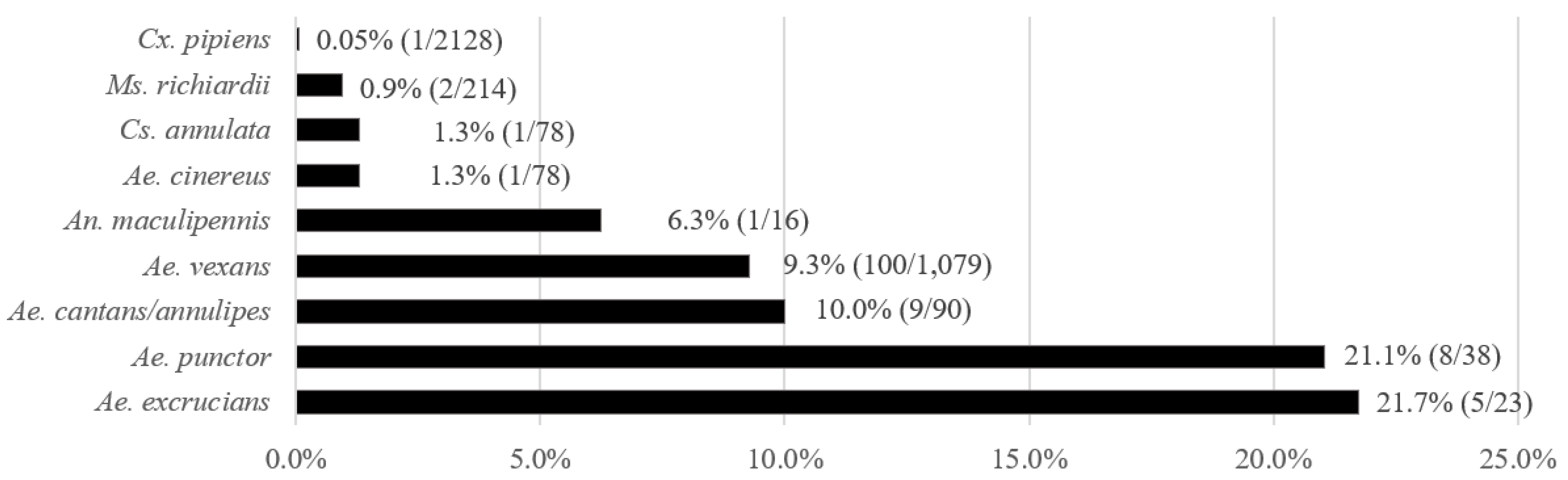
3.1. Prevalence of T. theileri in Mosquitoes
3.2. Prevalence of T. theileri in Deer Keds
3.3. Prevalence of T. melophagium in Sheep Keds
3.4. Prevalence of T. theileri in Tabanids
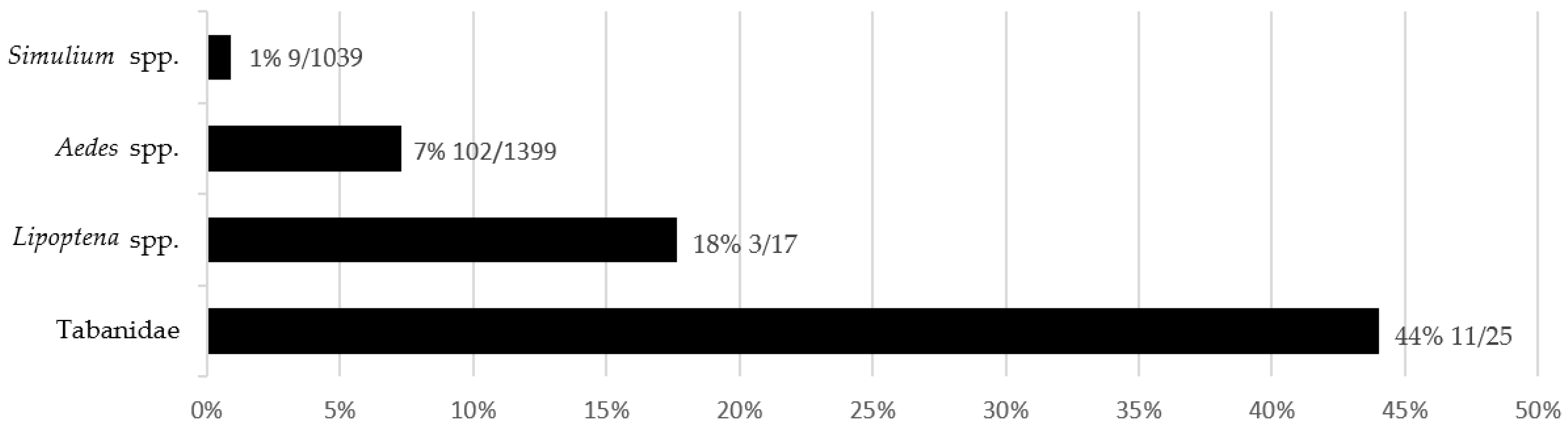
3.5. Comparison of T. theileri Prevalence in Insects
3.6. Detection of T. theileri in Deer Blood
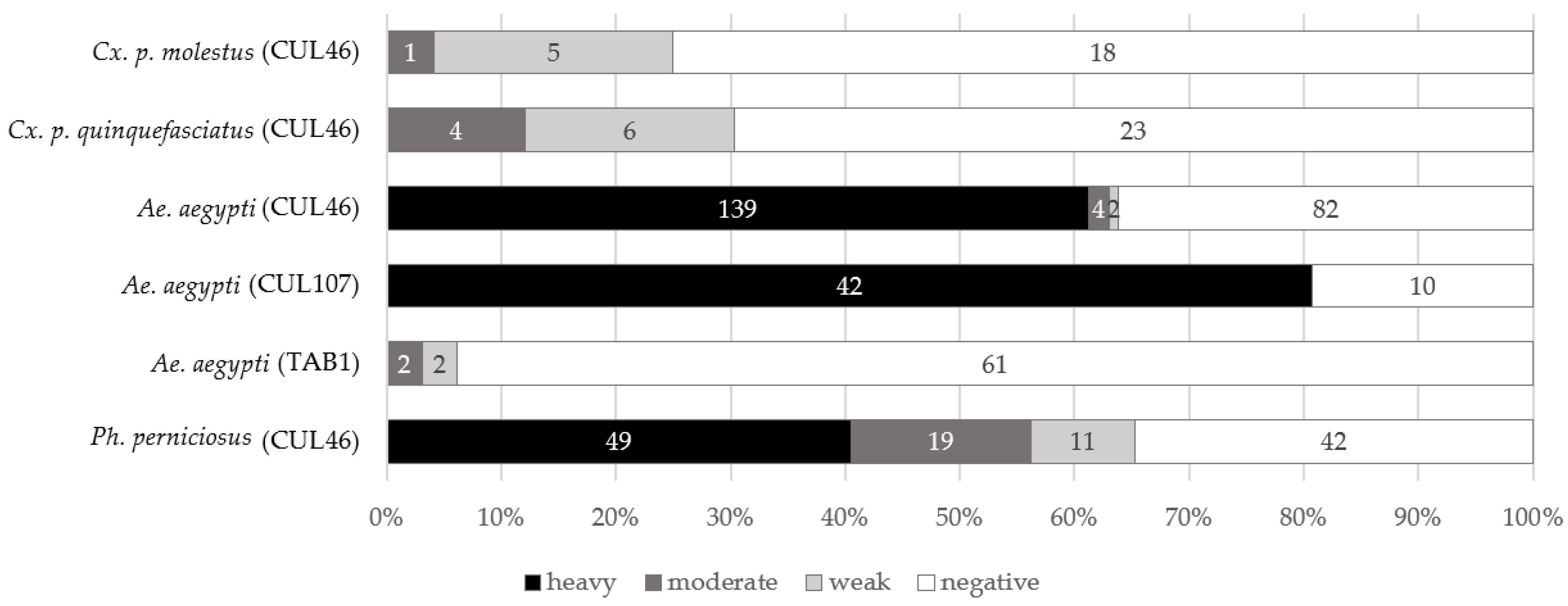
3.7. Experimental Infections of Mosquitoes
3.7.1. Experiments with Cx. p. quinquefasciatus and Cx. p. molestus
3.7.2. Infectious Experiments with Aedes aegypti
3.7.3. Experimental Infection of the Sand Fly Phlebotomus perniciosus
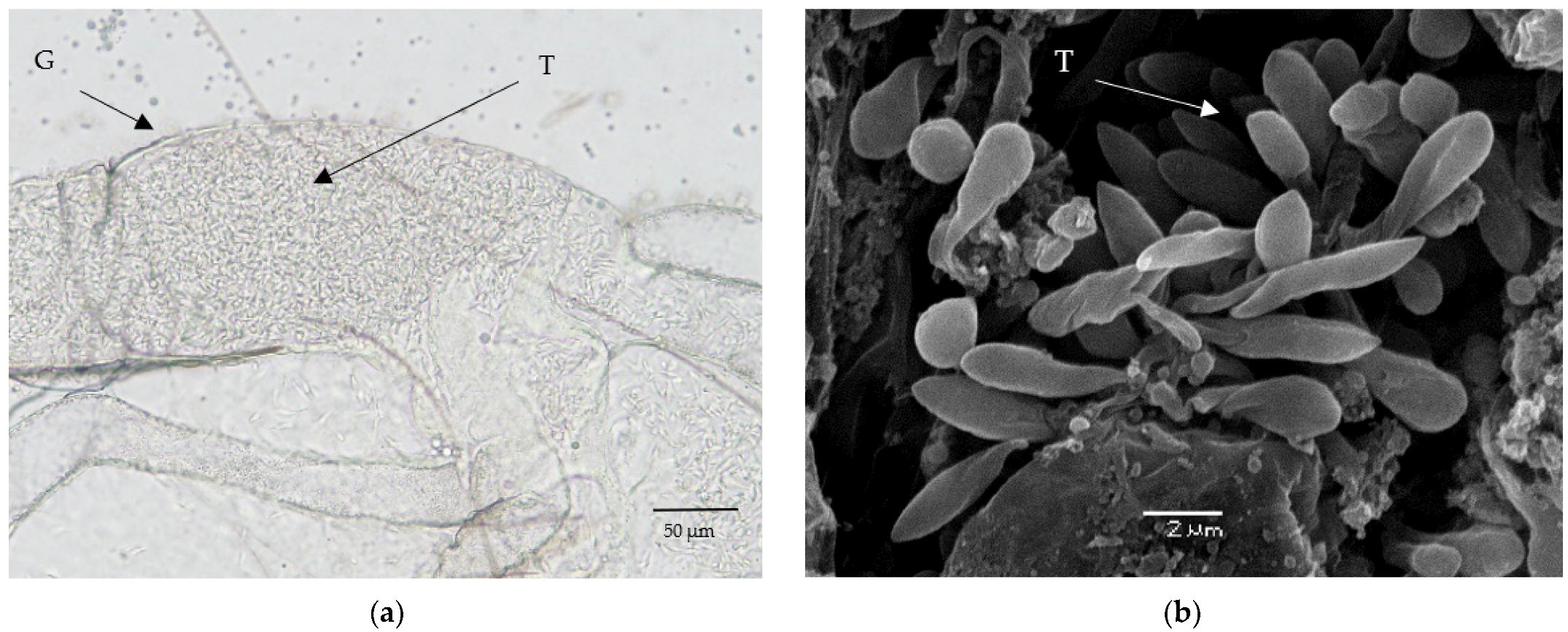
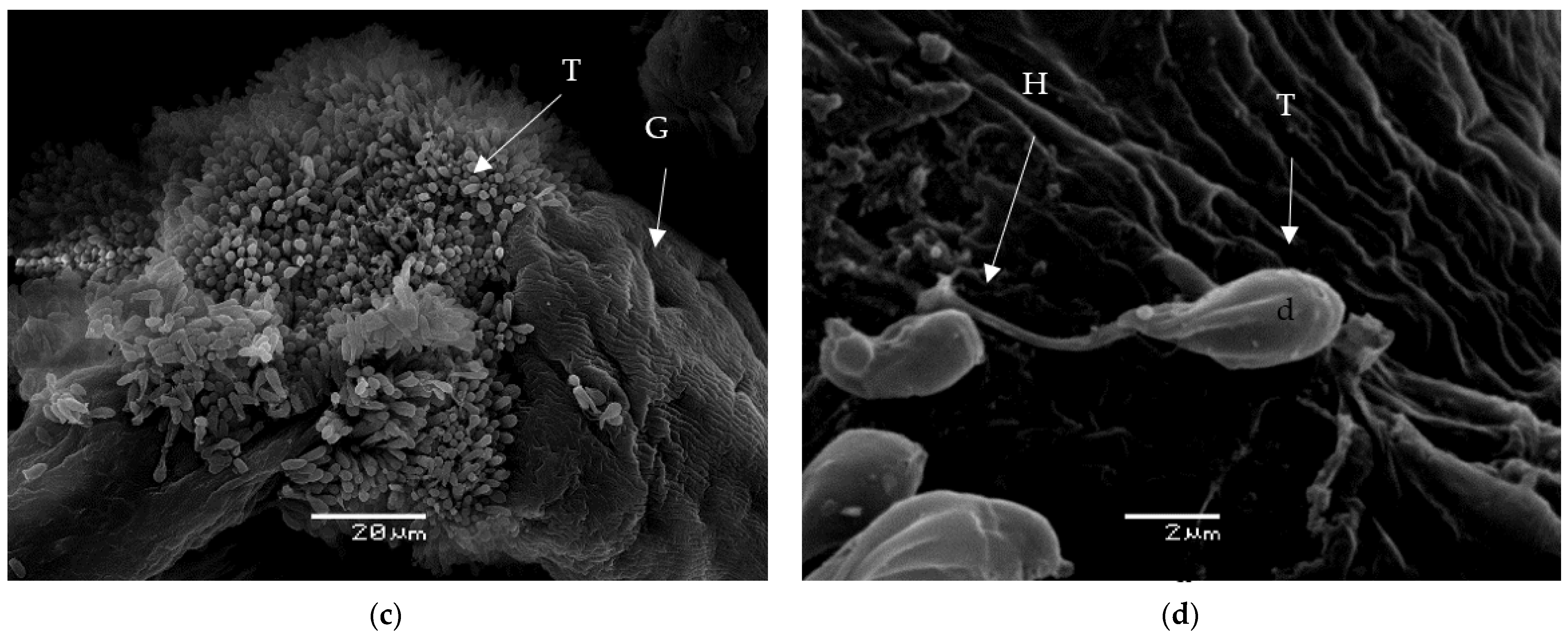
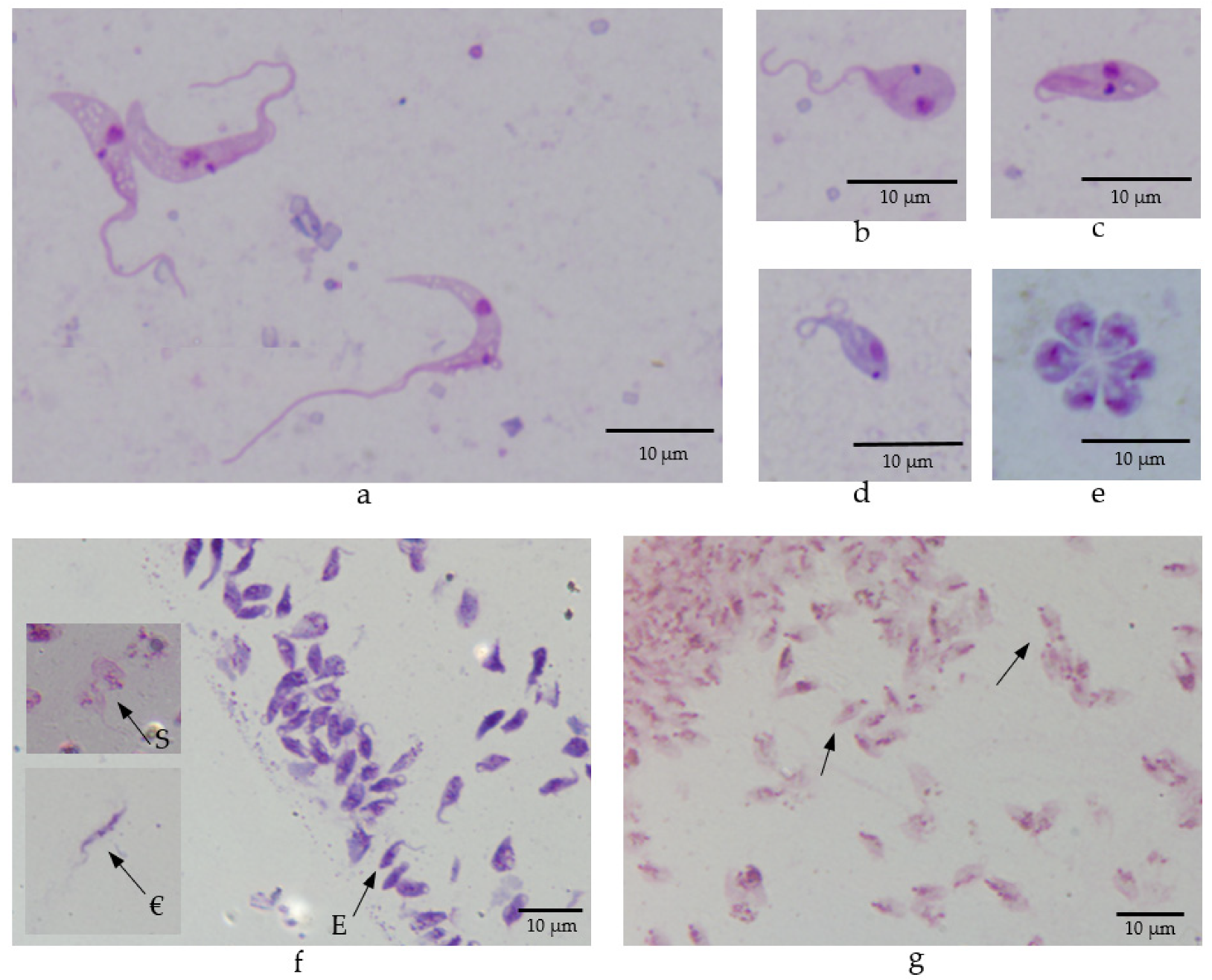
3.8. Morphology of Trypanosomes in Vectors
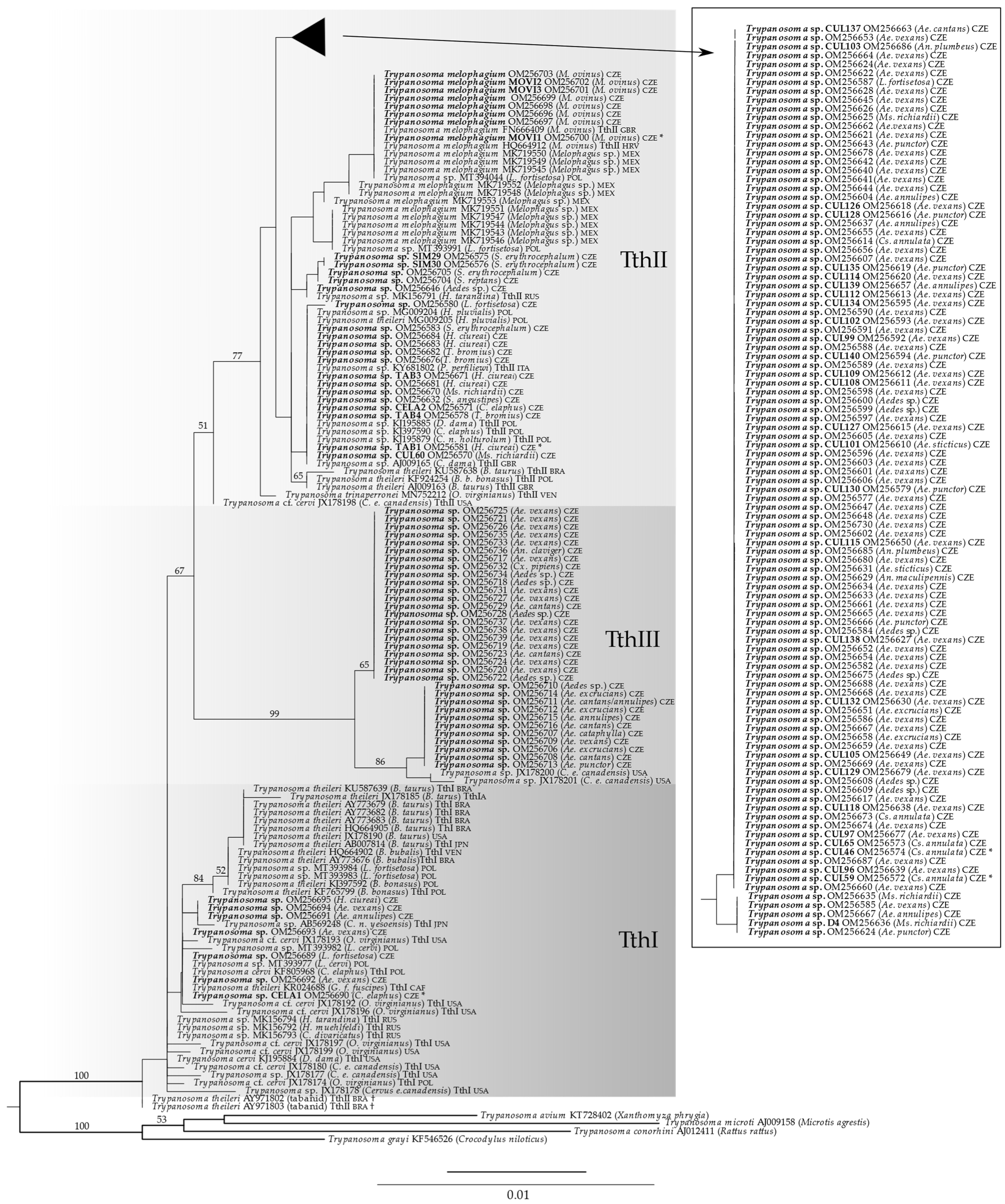
3.9. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adl, S.M.; Bass, D.; Lane, C.E.; Lukeš, J.; Schoch, C.L.; Smirnov, A.; Agatha, S.; Berney, C.; Brown, M.W.; Burki, F.; et al. Revisions to the Classification, Nomenclature, and Diversity of Eukaryotes. J. Eukaryot. Microbiol. 2019, 66, 4–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoare, C.A. The Trypanosomes of Mammals: Zoological Monograph; Blackwell Scientific Publications: Oxford, UK, 1972. [Google Scholar]
- Rodrigues, A.C.; Campaner, M.; Takata, C.S.A.; Dell’ Porto, A.; Milder, R.V.; Takeda, G.F.; Teixeira, M.M.G. Brazilian isolates of Trypanosoma (Megatrypanum) theileri: Diagnosis and differentiation of isolates from cattle and water buffalo based on biological characteristics and randomly amplified DNA sequences. Vet. Parasitol. 2003, 116, 185–207. [Google Scholar] [CrossRef]
- Kingston, N.; Morton, J.K. Trypanosoma cervi sp. n. from elk (Cervus canadensis) in Wyoming. J. Parasitol. 1975, 61, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Kingston, N.; Bobek, B.; Perzanowski, K.; Wita, I.; Maki, L. Description of Trypanosoma (Megatrypanum) stefanskii sp. n. from roe deer (Capreolus capreolus) in Poland. J. Helminthol. Soc. 1992, 59, 89–95. [Google Scholar]
- Garcia, H.A.; Blanco, P.A.; Rodrigues, A.C.; Rodrigues, C.M.F.; Takata, C.S.A.; Campaner, M.; Camargo, E.P.; Teixeira, M.M.G. Pan-american Trypanosoma (Megatrypanum) trinaperronei n. sp. in the white-tailed deer Odocoileus virginianus Zimmermann and its deer ked Lipoptena mazamae Rondani, 1878: Morphological, developmental and phylogeographical characterisation. Parasit. Vectors 2020, 13, 308. [Google Scholar] [CrossRef]
- Suganuma, K.; Kondoh, D.; Sivakumar, T.; Mizushima, D.; Elata, A.T.M.; Thekisoe, O.M.M.; Yokoyama, N.; Inoue, N. Molecular characterization of a new Trypanosoma (Megatrypanum) theileri isolate supports the two main phylogenetic lineages of this species in Japanese cattle. Parasitol. Res. 2019, 118, 1927–1935. [Google Scholar] [CrossRef]
- Wells, E.A. Subgenus Megatrypanum. In Biology of the Kinetoplastida; Lumsden, W.H.R., Evans, D.A., Eds.; Academic Press: Cambridge, MA, USA, 1976; Volume 1, pp. 257–284. [Google Scholar]
- Doherty, M.L.; Windle, H.; Voorheis, H.P.; Larkin, H.; Casey, M.; Clery, D.; Murray, M. Clinical disease associated with Trypanosoma theileri infection in a calf in Ireland. Vet. Rec. 1993, 132, 653–656. [Google Scholar] [CrossRef]
- Seifi, H.A. Clinical trypanosomosis due to Trypanosoma theileri in a cow in Iran. Trop. Anim. Health Prod. 1995, 27, 93–94. [Google Scholar] [CrossRef]
- Greco, A.; Loria, G.R.; Dara, S.; Luckins, T.; Sparagano, O. First isolation of Trypanosoma theileri in sicilian cattle. Vet. Res. Commun. 2000, 24, 471–475. [Google Scholar] [CrossRef]
- Braun, U.; Rogg, E.; Walser, M.; Nehrbass, D.; Guscetti, F.; Mathis, A.; Deplazes, P. Trypanosoma theileri in the cerebrospinal fluid and brain of a heifer with suppurative meningoencephalitis. Vet. Rec. 2002, 150, 18–19. [Google Scholar] [CrossRef]
- Sood, N.K.; Singla, L.D.; Singh, R.S.; Uppal, S.K. Association of Trypanosoma theileri with peritonitis in a pregnant cross-bred cow: A case report. Vet. Med. 2011, 56, 82–84. [Google Scholar] [CrossRef] [Green Version]
- Hajihassani, A.; Maroufi, S.; Esmaeilnejad, B.; Khorram, H.; Tavassoli, M.; Dalir-Naghadeh, B.; Samiei, A. Hemolytic anemia associated with Trypanosoma theileri in a cow from Kurdistan province, West of Iran. Vet. Res. Forum 2020, 11, 191–193. [Google Scholar] [CrossRef] [PubMed]
- Bittner, L.; Krämer, K.; Wöckel, A.; Snedec, T.; Delling, C.; Böttcher, D.; Köller, G.; Baumgartner, W.; Richardt, W.; Starke, A. Malnutrition as the cause of recumbency in suckler cows associated with Trypanosoma theileri infection. Acta Vet. Scand. 2021, 63, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.C.; Paiva, F.; Campaner, M.; Stevens, J.R.; Noyes, H.A.; Teixeira, M.M.G. Phylogeny of Trypanosoma (Megatrypanum) theileri and related trypanosomes reveals lineages of isolates associated with artiodactyl hosts diverging on SSU and ITS ribosomal sequences. Parasitology 2006, 132, 215–224. [Google Scholar] [CrossRef]
- Garcia, H.A.; Rodrigues, A.C.; Martinkovic, F.; Minervino, A.H.H.; Campaner, M.; Nunes, V.L.B.; Paiva, F.; Hamilton, P.B.; Teixeira, M.M.G. Multilocus phylogeographical analysis of Trypanosoma (Megatrypanum) genotypes from sympatric cattle and water buffalo populations supports evolutionary host constraint and close phylogenetic relationships with genotypes found in other ruminants. Int. J. Parasitol. 2011, 41, 1385–1396. [Google Scholar] [CrossRef]
- Yokoyama, N.; Sivakumar, T.; Fukushi, S.; Tattiyapong, M.; Tuvshintulga, B.; Kothalawala, H.; Silva, S.S.P.; Igarashi, I.; Inoue, N. Genetic diversity in Trypanosoma theileri from Sri Lankan cattle and water buffaloes. Vet. Parasitol. 2015, 207, 335–341. [Google Scholar] [CrossRef]
- Pacheco, M.A.; Cepeda, A.S.; Bernotienė, R.; Lotta, I.A.; Matta, N.E.; Valkiūnas, G.; Escalante, A.A. Primers targeting mitochondrial genes of avian haemosporidians: PCR detection and differential DNA amplification of parasites belonging to different genera. Int. J. Parasitol. 2018, 48, 657–670. [Google Scholar] [CrossRef]
- Gibson, W.; Pilkington, J.G.; Pemberton, J.M. Trypanosoma melophagium from the sheep ked Melophagus ovinus on the island of St Kilda. Parasitology 2010, 137, 1799–1804. [Google Scholar] [CrossRef] [Green Version]
- Martinković, F.; Matanović, K.; Rodrigues, A.C.; Garcia, H.A.; Teixeira, M.M.G. Trypanosoma (Megatrypanum) melophagium in the sheep ked Melophagus ovinus from organic farms in Croatia: Phylogenetic inferences support restriction to sheep and sheep keds and close relationship with trypanosomes from other ruminant species. J. Eukaryot. Microbiol. 2012, 59, 134–144. [Google Scholar] [CrossRef]
- Fisher, A.C.; Schuster, G.; Cobb, W.J.; James, A.M.; Cooper, S.M.; Peréz de León, A.A.; Holman, P.J. Molecular characterization of Trypanosoma (Megatrypanum) spp. infecting cattle (Bos taurus), white-tailed deer (Odocoileus virginianus), and elk (Cervus elaphus canadensis) in the United States. Vet. Parasitol. 2013, 197, 29–42. [Google Scholar] [CrossRef]
- Böse, R.; Friedhoff, K.T.; Olbrich, S. Transmission of Megatrypanum Trypanosomes to Cervus dama by Tabanidae. J. Protozool. 1987, 34, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Böhm, M.; White, P.C.L.; Chambers, J.; Smith, L.; Hutchings, M.R. Wild deer as a source of infection for livestock and humans in the UK. Vet. J. 2007, 174, 260–276. [Google Scholar] [CrossRef] [PubMed]
- Kingston, N.; Thorne, E.T.; Thomas, G.M.; McHolland, L.; Trueblood, M.S. Further studies on trypanosomes in game animals in Wyoming II. J. Wildl. Dis. 1981, 17, 539–546. [Google Scholar] [CrossRef] [Green Version]
- Lanevschi-Pietersma, A.; Ogunremi, O.; Desrochers, A. Parasitemia in a neonatal bison calf. Vet. Clin. Pathol. 2004, 33, 173–176. [Google Scholar] [CrossRef]
- Nijhout, H.F.; Grant, M.C. Diuresis after a bloodmeal in female Anopheles freeborni. J. Insect Physiol. 1978, 24, 293–298. [Google Scholar] [CrossRef]
- Briegel, H.; Rezzonico, L. Concentration of host blood protein during feeding by anopheline mosquitoes (Diptera: Culicidae). J. Med. Entomol. 1985, 22, 612–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddrell, S.H. Excretion in the blood-sucking bug, Rhodnius prolixus Stal. I. The control of diuresis. J. Exp. Biol. 1964, 41, 163–176. [Google Scholar] [CrossRef]
- Gee, J.D. The control of diuresis in the tsetse fly Glossina austeni: A preliminary investigation of the diuretic hormone. J. Exp. Biol. 1975, 63, 391–401. [Google Scholar] [CrossRef]
- Jones, J.C.; Brandt, E. Fluid excretion by adult Aedes aegypti mosquitoes. J. Insect Physiol. 1981, 27, 545–549. [Google Scholar] [CrossRef]
- Sádlová, J.; Reishig, J.; Volf, P. Prediuresis in female Phlebotomus sandflies (Diptera: Psychodidae). Eur. J. Entomol. 1998, 95, 643–647. [Google Scholar]
- Fialová, M.; Santolíková, A.; Brotánková, A.; Brzoňová, J.; Svobodová, M. Complete Life Cycle of Trypanosoma thomasbancrofti, an Avian Trypanosome Transmitted by Culicine Mosquitoes. Microorganisms 2021, 9, 2101. [Google Scholar] [CrossRef] [PubMed]
- Mutero, C.M.; Mutinga, M.J. Defecation by Anopheles arabiensis mosquitoes of host blood infected with live Trypanosoma congolense. Trop. Med. Parasitol. 1993, 44, 23–26. [Google Scholar] [PubMed]
- Sádlová, J.; Volf, P. Occurrence of Leishmania major in sandfly urine. Parasitology 1999, 118, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Galen, S.C.; Borner, J.; Perkins, S.L.; Weckstein, J.D. Phylogenomics from transcriptomic “bycatch” clarify the origins and diversity of avian trypanosomes in North America. PLoS ONE 2020, 15. [Google Scholar] [CrossRef]
- Böse, R.; Heister, N.C. Development of Trypanosoma (M.) theileri in Tabanids. J. Eukaryot. Microbiol. 1993, 40, 788–792. [Google Scholar] [CrossRef]
- Ganyukova, A.I.; Zolotarev, A.V.; Malysheva, M.N.; Frolov, A.O. First record of Trypanosoma theileri-like flagellates in horseflies from Northwest Russia. Protistology 2018, 12, 223–230. [Google Scholar] [CrossRef]
- Böse, R.; Petersen, K. Lipoptena cervi (Diptera), a potencial vector of Megatrypanum trypanosomes of deer (Cervidae). Parasitol. Res. 1991, 77, 723–725. [Google Scholar] [CrossRef]
- Werszko, J.; Steiner-Bogdaszewska, Z.; Jeżewski, W.; Szewczyk, T.; Kuryło, G.; Wołkowycki, M.; Wróblewski, P.; Karbowiak, G. Molecular detection of Trypanosoma spp. in Lipoptena cervi and Lipoptena fortisetosa (Diptera: Hippoboscidae) and their potential role in the transmission of pathogens. Parasitology 2020, 147, 1629–1635. [Google Scholar] [CrossRef]
- Schoener, E.; Uebleis, S.S.; Cuk, C.; Nawratil, M.; Obwaller, A.G.; Zechmeister, T.; Lebl, K.; Rádrová, J.; Zittra, C.; Votýpka, J.; et al. Trypanosomatid parasites in Austrian mosquitoes. PLoS ONE 2018, 13, e0141332. [Google Scholar] [CrossRef] [Green Version]
- Calzolari, M.; Rugna, G.; Clementi, E.; Carra, E.; Pinna, M.; Bergamini, F.; Fabbi, M.; Dottori, M.; Sacchi, L.; Votýpka, J. Isolation of a trypanosome related to Trypanosoma theileri (Kinetoplastea: Trypanosomatidae) from Phlebotomus perfiliewi (Diptera: Psychodidae). Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Votýpka, J.; Rádrová, J.; Skalický, T.; Jirků, M.; Jirsová, D.; Mihalca, A.D.; D’Amico, G.; Petrželková, K.J.; Modrý, D.; Lukeš, J. A tsetse and tabanid fly survey of African great apes habitats reveals the presence of a novel trypanosome lineage but the absence of Trypanosoma brucei. Int. J. Parasitol. 2015, 45, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Ngomtcho, S.C.H.; Weber, J.S.; Ngo Bum, E.; Gbem, T.T.; Kelm, S.; Achukwi, M.D. Molecular screening of tsetse flies and cattle reveal different Trypanosoma species including T. grayi and T. theileri in northern Cameroon. Parasites Vectors 2017, 10, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krinsky, A.W.L.; Burgdorfer, W. Trypanosomes in Amblyomma americanum from Oklahoma. J. Parasitol. 1976, 62, 824–825. [Google Scholar] [CrossRef] [PubMed]
- Shastri, U.V.; Deshpande, P.D. Hyalomma anatolicum anatolicum (Koch, 1844) as a possible vector for transmission of Trypanosoma theileri, Laveran, 1902 in cattle. Vet. Parasitol. 1981, 9, 151–155. [Google Scholar] [CrossRef]
- Latif, A.A.; Bakheit, M.A.; Mohamed, A.E.; Zweygarth, E. High infection rates of the tick Hyalomma anatolicum anatolicum with Trypanosoma theileri. Onderstepoort J. Vet. Res. 2004, 71, 251–256. [Google Scholar] [CrossRef] [Green Version]
- Martins, J.R.; Leite, R.C.; Doyle, R.L. Tripanosomatides like Trypanosoma theileri in the cattle tick Boophilus microplus. Rev. Bras. Parasitol. Vet. 2008, 17, 113–114. [Google Scholar] [CrossRef] [Green Version]
- Molyneux, D.H. Trypanosoma (Megatrypanum) melophagium: Modes of attachment of parasites to mid-gut, hindgut and rectum of the sheep ked, Melophagus ovinus. Acta Trop. 1975, 32, 65–74. [Google Scholar]
- Molyneux, D.H.; Selkirk, M.; Lavin, D. Trypanosoma (Megatrypanum) melophagium in the sheep ked, Melophagus ovinus: A scanning electron microscope (SEM) study of the parasites and the insect gut wall surfaces. Acta Trop. 1978, 35, 319–328. [Google Scholar]
- Votýpka, J.; Szabová, J.; Rádrová, J.; Zídková, L.; Svobodová, M. Trypanosoma culicavium sp. nov., an avian trypanosome transmitted by Culex mosquitoes. Int. J. Syst. Evol. Microbiol. 2012, 62, 745–754. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.C.; Garcia, H.A.; Batista, J.S.; Minervino, A.H.H.; Góes-Cavalcante, G.; Maia da Silva, F.; Ferreira, R.C.; Campaner, M.; Paiva, F.; Teixeira, M.M.G. Characterization of spliced leader genes of Trypanosoma (Megatrypanum) theileri: Phylogeographical analysis of brazilian isolates from cattle supports spatial clustering of genotypes and parity with ribosomal markers. Parasitology 2010, 137, 111–122. [Google Scholar] [CrossRef]
- Pacheco, T.D.A.; Marcili, A.; da Costa, A.P.; Witter, R.; Melo, A.L.T.; Boas, R.V.; Chitarra, C.S.; Dutra, V.; Nakazato, L.; de Pacheco, R.C. Genetic diversity and molecular survey of Trypanosoma (Megatrypanum) theileri in cattle in Brazil’s western Amazon region. Rev. Bras. Parasitol. Vet. 2018, 27, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Jaimes-Dueñez, J.; Triana-Chávez, O.; Mejía-Jaramillo, A.M. Spatial-temporal and phylogeographic characterization of Trypanosoma spp. in cattle (Bos taurus) and buffaloes (Bubalus bubalis) reveals transmission dynamics of these parasites in Colombia. Vet. Parasitol. 2018, 249, 30–42. [Google Scholar] [CrossRef] [PubMed]
- Kramář, J. Fauna ČSR, Svazek 13, Komáři Bodaví—Culicinae; Czechoslovak Academy of Sciences: Prague, Czech Republic, 1958. [Google Scholar]
- Chvála, M.; Hůrka, K.; Chalupský, J.; Knoz, J.; Minář, J.; Országh, I. Hippoboscidae—Klošovití. In Fauna ČSR, Svazek 22, Krevsající Mouchy a Střečci; Nakladatelství Československé Akademie Věd: Prague, Czech Republic, 1980; pp. 475–478. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Medlin, L.K.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Maslov, D.A.; Lukeš, J.; Jirků, M.; Simpson, L. Phylogeny of trypanosomes as inferred from the small and large subunit rRNAs: Implications for the evolution of parasitism in the trypanosomatid protozoa. Mol. Biochem. Parasitol. 1996, 75, 197–205. [Google Scholar] [CrossRef]
- Santolíková, A. Role Klošů v Přenosu Ptačích Trypanosom (The Role of Hippoboscids in Avian Trypanosomes Transmission). Master’s Thesis, Charles University, Prague, Czech Republic, 2019. [Google Scholar]
- Zídková, L.; Cepicka, I.; Szabová, J.; Svobodová, M. Biodiversity of avian trypanosomes. Infect. Genet. Evol. 2012, 12, 102–112. [Google Scholar] [CrossRef]
- Hlaváčová, J.; Votýpka, J.; Volf, P. The effect of temperature on Leishmania (Kinetoplastida: Trypanosomatidae) development in sand flies. J. Med. Entomol. 2013, 50, 955–958. [Google Scholar] [CrossRef] [Green Version]
- Rasband, W.S. ImageJ; National Institute of Health: Bethesda, MD, USA, 1997. [Google Scholar]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucl. Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Hall, T. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Böse, R.; Petersen, K.; Pospichal, H.; Buchanan, N.; Tait, A. Characterization of Megatrypanum trypanosomes from European Cervidae. Parasitology 1993, 107, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Börstler, J.; Jöst, H.; Garms, R.; Krüger, A.; Tannich, E.; Becker, N.; Schmidt-Chanasit, J.; Lühken, R. Host-feeding patterns of mosquito species in Germany. Paras. Vectors 2016, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schönenberger, A.C.; Wagner, S.; Tuten, H.C.; Schaffner, F.; Torgerson, P.; Furrer, S.; Mathis, A.; Silaghi, C. Host preferences in host-seeking and blood-fed mosquitoes in Switzerland. Med. Vet. Entomol. 2016, 30, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Rádrová, J.; Šeblová, V.; Votýpka, J. Feeding behavior and spatial distribution of Culex mosquitoes (Diptera: Culicidae) in wetland areas of the Czech Republic. J. Med. Entomol. 2013, 50, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Small, R.W. A review of Melophagus ovinus (L.), the sheep ked. Vet. Parasitol. 2005, 130, 141–155. [Google Scholar] [CrossRef]
- Seblova, V.; Sadlova, J.; Carpenter, S.; Volf, P. Development of Leishmania parasites in Culicoides nubeculosus (Diptera: Ceratopogonidae) and implications for screening vector competence. J. Med. Entomol. 2012, 49, 967–970. [Google Scholar] [CrossRef] [Green Version]
- Werszko, J.; Szewczyk, T.; Steiner-Bogdaszewska, Z.; Wróblewski, P.; Karbowiak, G.; Laskowski, Z. Molecular detection of Megatrypanum trypanosomes in tabanid flies. Med. Vet. Entomol. 2019, 1–5. [Google Scholar] [CrossRef]
- Hoffmann, M.; Buscher, G.; Friedhoff, K.T. Stercorarian trypanosomes from deer (Cervidae) in Germany. J. Protozool. 1984, 31, 581–584. [Google Scholar] [CrossRef]
- Neumuller, M.; Nilsson, K.; Pahlson, C. Trypanosoma spp. in Swedish game animals. Parasitol. Res. 2012, 110, 135–139. [Google Scholar] [CrossRef]
- Githeko, A.K.; Lindsay, S.W.; Confalonieri, U.E.; Patz, J.A. Climate change and vector-borne diseases: A regional analysis. Bull. World Health Organ. 2000, 78, 1136–1147. [Google Scholar] [CrossRef]
- Gage, K.L.; Burkot, T.R.; Eisen, R.J.; Hayes, E.B. Climate and vectorborne diseases. Am. J. Prev. Med. 2008, 35, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, J.M.; Lazzari, C.R.; Lahondère, C. Effects of the environmental temperature on Aedes aegypti and Aedes albopictus mosquitoes: A review. Insects 2018, 9, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galková, Z. Tiplíci Jako Přenašeči Infekčních Onemocnění a Jejich Výskyt na Území ČR. Master’s Thesis, Charles University, Prague, Czech Republic, 2010. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vector Species | Trypanosome Isolate | Environmental Temperature (°C) |
|---|---|---|
| Culex p. quinquefasciatus | CUL59 (CUL/CZ/2015/CUL59) ex Culiseta annulata * | 15 |
| 21 | ||
| Cx. p. quinquefasciatus Cx. p. molestus | CUL46 (CUL/CZ/2014/CUL46) ex Cs. annulata * | 8–11 |
| 15 | ||
| 8–11→15 | ||
| 21 | ||
| Aedes aegypti | CUL46 (CUL/CZ/2014/CUL46) ex Cs. annulata | 21 |
| CUL107 (AED/CZ/2018/CUL107) ex Aedes vexans | ||
| CELA1 (CER/CZ/2017/CELA1) ex Cervus elaphus | ||
| TAB1 (HYB/CZ/2019/TAB1) ex Hybomitra ciureai | ||
| MOVI1 (MEL/CZ/2017/MOVI1) ex Melophagus ovinus | ||
| Phlebotomus pernicious | CUL46 (CUL/CZ/2014/CUL46) ex Cs. annulata * | 21 |
| Species | n Specimens | n Kinetoplastid + (Prevalence) | n T. Theileri + (Prevalence) |
|---|---|---|---|
| Hybomitra ciureai | 16 | 11 (69%) | 8 (50%) |
| Tabanus bromius | 6 | 4 | 3 |
| Haematopota pluvialis | 2 | 0 | 0 |
| Atylotus leowianus | 1 | 0 | 0 |
| Total | 25 | 15 (60%) | 11 (44%) |
| Morphotype | Tabanid Mean (Range) (µm) | Mosquito Mean (Range) (µm) | Sandfly Mean (Range) (µm) | Prediuresis Mean (Range) (µm) |
|---|---|---|---|---|
| Elongated epimastigote | 16.0 (8.9–22.6) * | 15.3 (12.6–23.5) * | 16.8 (11.4–25.0) | - |
| Sphaeromastigote | - | 8.4 (7.0–10.0) * | 7.4 (3.6–13.9) | - |
| Droplet-shaped epimastigote | 7.5 (5.1–11.0) | 7.2 (3.3–23.0) | 8.8 (5.0–16.5) | 5.3 (4.3–7.0) |
| Metacyclic stages | 4.5 (3.9–7.8) | 5.0 (3.3–8.0) | 5.5 (3.5–7.4) | 5.3 (3.8–6.8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brotánková, A.; Fialová, M.; Čepička, I.; Brzoňová, J.; Svobodová, M. Trypanosomes of the Trypanosoma theileri Group: Phylogeny and New Potential Vectors. Microorganisms 2022, 10, 294. https://doi.org/10.3390/microorganisms10020294
Brotánková A, Fialová M, Čepička I, Brzoňová J, Svobodová M. Trypanosomes of the Trypanosoma theileri Group: Phylogeny and New Potential Vectors. Microorganisms. 2022; 10(2):294. https://doi.org/10.3390/microorganisms10020294
Chicago/Turabian StyleBrotánková, Anna, Magdaléna Fialová, Ivan Čepička, Jana Brzoňová, and Milena Svobodová. 2022. "Trypanosomes of the Trypanosoma theileri Group: Phylogeny and New Potential Vectors" Microorganisms 10, no. 2: 294. https://doi.org/10.3390/microorganisms10020294
APA StyleBrotánková, A., Fialová, M., Čepička, I., Brzoňová, J., & Svobodová, M. (2022). Trypanosomes of the Trypanosoma theileri Group: Phylogeny and New Potential Vectors. Microorganisms, 10(2), 294. https://doi.org/10.3390/microorganisms10020294