
Lactobacilli Strain Mixture Alleviates Bacterial Vaginosis through Antibacterial and Antagonistic Activity in Gardnerella vaginalis-Infected C57BL/6 Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Lactobacilli Strains and GV
2.2. Detection of Hydrogen Peroxide (H2O2) Production
2.3. Analysis of Lactic Acid Production
2.4. Resistance to Spermicide
2.5. Antibacterial Effects of Lactobacilli Strains on GV Growth
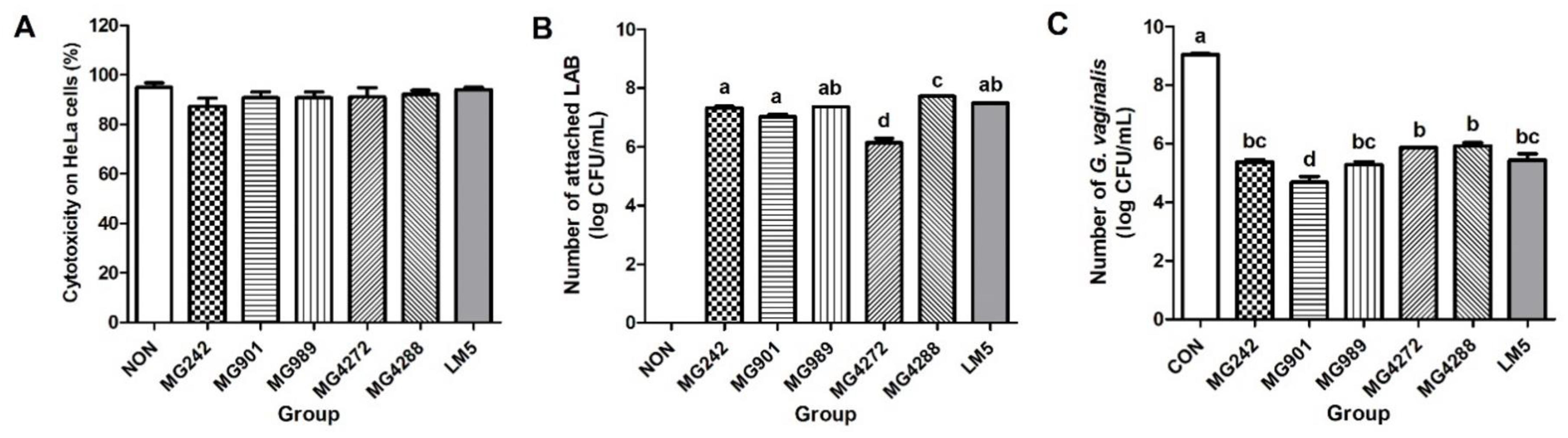
2.6. The Cytotoxic Effect of Lactobacilli Strains on HeLa Cells
2.7. Adhesion Ability of Lactobacilli Strains to HeLa Cells
2.8. Antagonistic Activity of Lactobacilli Strains against GV Adhesion to HeLa Cells
2.9. GV-Infected BV Mice Model and LM5 Administration
2.10. GV Proliferation in Vaginal Tissues
2.11. Myeloperoxidase (MPO) Activity in Vaginal Lysates
2.12. Pro-Inflammatory Cytokines Levels in Vaginal Tissue Lysates
2.13. Histopathological Examination
2.14. Statistical Analysis
3. Results
3.1. Lactic Acid and H2O2 Production
3.2. Nonoxynol-9 Susceptibility
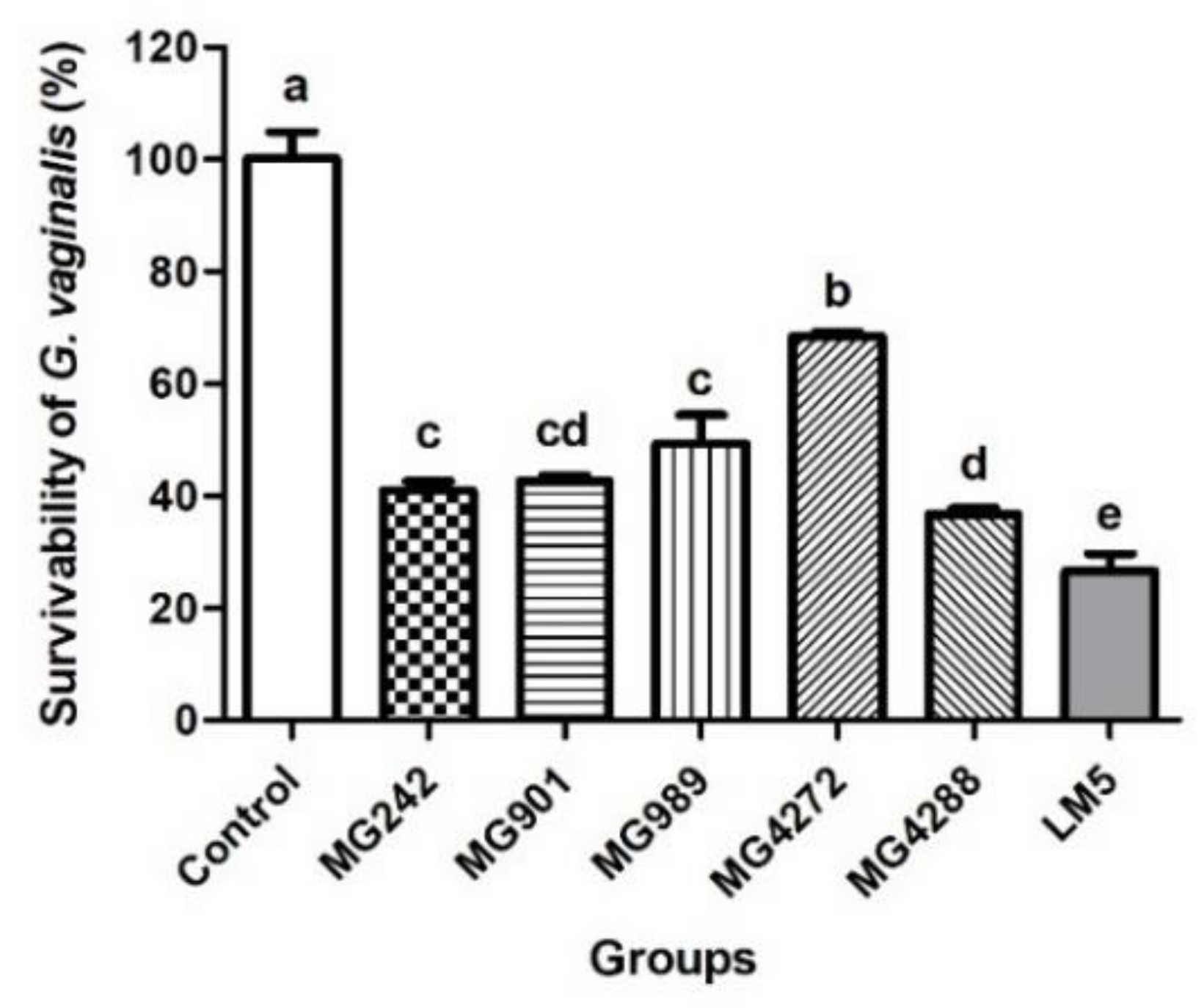
3.3. Antibacterial Effect of Lactobacilli Strains against GV
3.4. Antagonistic Effect of Lactobacilli Strains on GV Adhesion to HeLa Cells
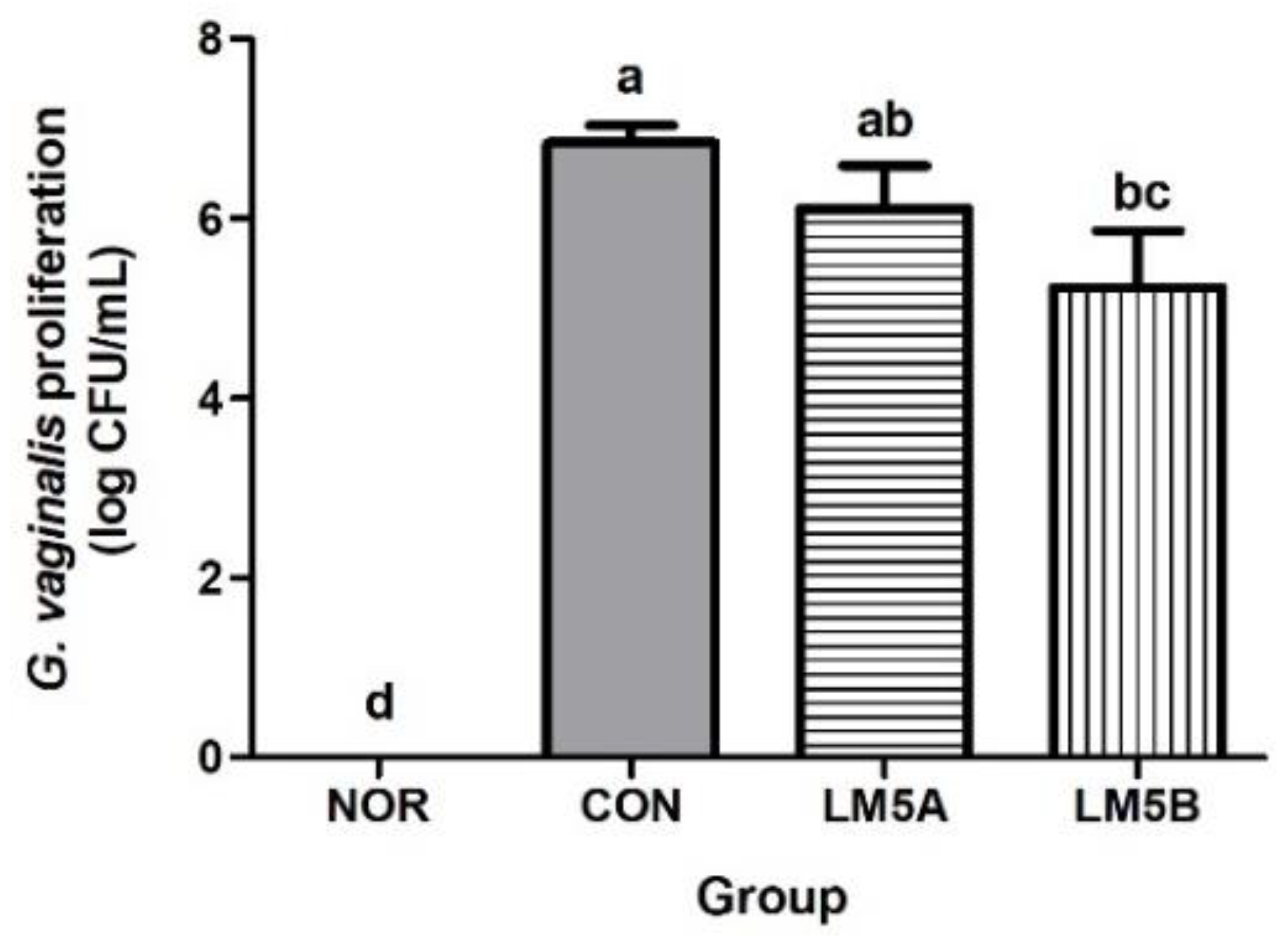
3.5. Inhibitory Effect of LM5 on Vaginal GV Proliferation in GV-Infected BV Mice
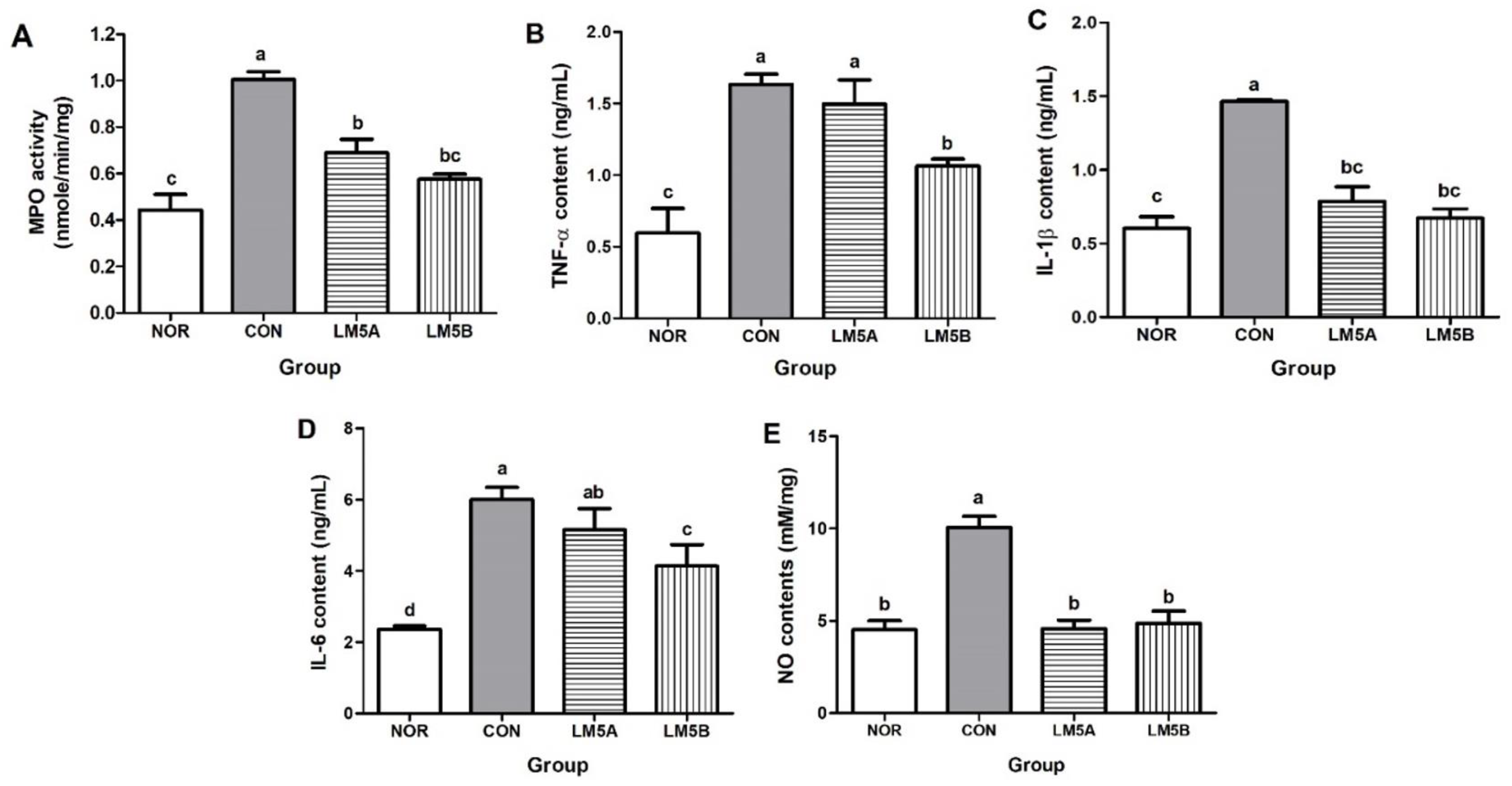
3.6. Inhibitory Effect of LM5 on MPO Activity in Vaginal Tissue
3.7. Inhibitory Effect of LM5 on Proinflammatory Cytokines Production in Vaginal Tissue
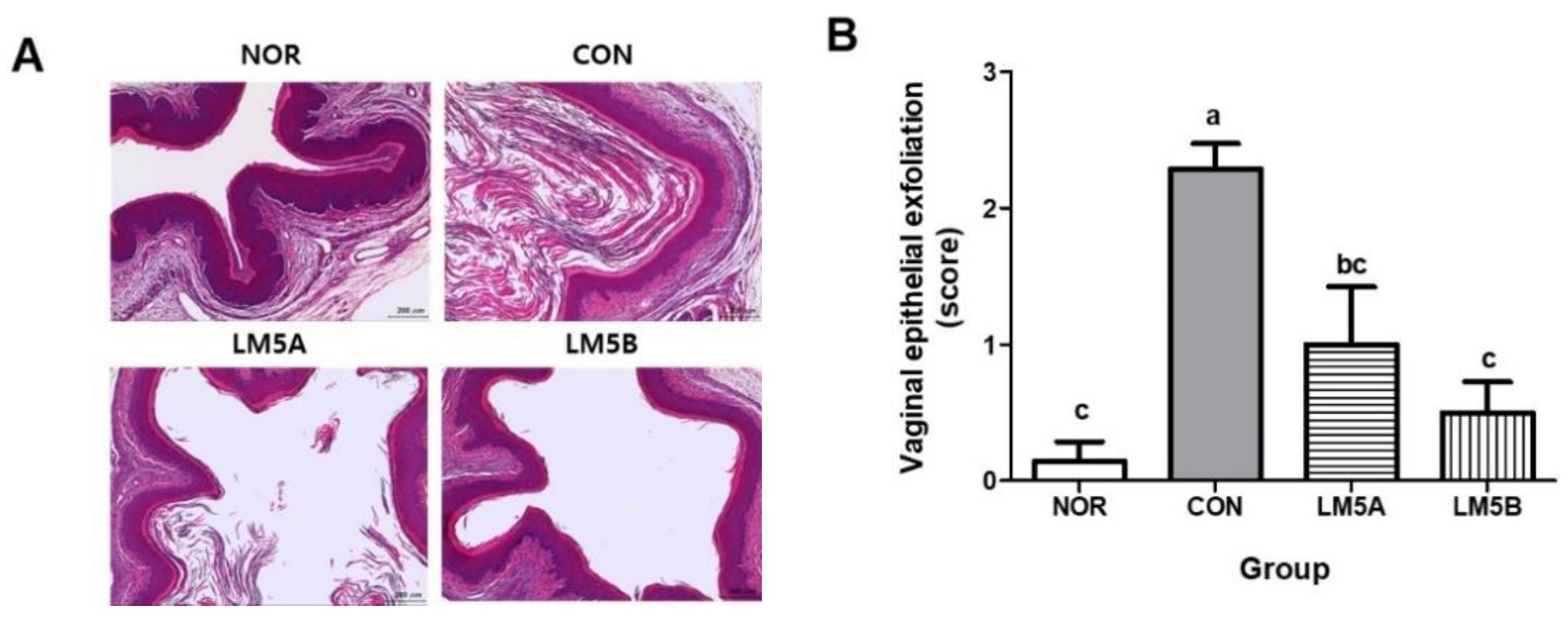
3.8. Histopathological Analysis on Vaginal Tissues of GV-Infected Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mendling, W. Vaginal microbiota. Adv. Exp. Med. Biol. 2016, 902, 83–93. [Google Scholar] [PubMed]
- Moosa, Y.; Kwon, D.; de Oliveira, T.; Wong, E.B. Determinants of vaginal microbiota composition. Front. Cell. Infect. Microbiol. 2020, 10, 467. [Google Scholar] [CrossRef] [PubMed]
- Paladine, H.L.; Desai, U.A. Vaginitis: Diagnosis and treatment. Am. Fam. Physician 2018, 97, 321–329. [Google Scholar]
- Redelinghuys, M.J.; Geldenhuys, J.; Jung, H.; Kock, M.M. Bacterial vaginosis: Current diagnostic avenues and future opportunities. Front. Cell. Infect. Microbiol. 2020, 10, 354. [Google Scholar] [CrossRef] [PubMed]
- Muzny, C.A.; Blanchard, E.; Taylor, C.M.; Aaron, K.J.; Talluri, R.; Griswold, M.E.; Redden, D.T.; Luo, M.; Welsh, D.A.; Van Der Pol, W.J. Identification of key bacteria involved in the induction of incident bacterial vaginosis: A prospective study. J. Infect. Dis. 2018, 218, 966–978. [Google Scholar] [PubMed]
- Jones, A. Bacterial Vaginosis: A review of treatment, recurrence, and disparities. J. Nurse Pract. 2019, 15, 420–423. [Google Scholar] [CrossRef] [Green Version]
- Kaambo, E.; Africa, C.; Chambuso, R.; Passomore, J.S. Vaginal microbiomes associated with aerobic vaginitis and bacterial vaginosis. Front. Public Health 2018, 6, 78. [Google Scholar] [CrossRef]
- Kovachev, S. Defense factors of vaginal lactobacilli. Crit. Rev. Microbiol. 2018, 44, 31–39. [Google Scholar] [CrossRef]
- Chen, X.; Lu, Y.; Chen, T.; Li, R. The female vaginal microbiome in health and bacterial vaginosis. Front. Cell. Infect. Microbiol. 2021, 11, 631972. [Google Scholar] [CrossRef] [PubMed]
- Tachedjian, G.; Aldunate, M.; Bradshaw, C.S.; Cone, R.A. The role of lactic acid production by probiotic Lactobacillus species in vginal health. Res. Microbiol. 2017, 168, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Bastani, P.; Homayouni, A.; Ziyadi, S.; Tabrizian, V.G. Dairy probiotic foods and bacterial vaginosis: A review on mechanism of action. In Probiotics, ed EC Rigobelo; Intech Publishing: Rijeka, Croatia, 2012; pp. 445–456. [Google Scholar]
- Reid, G.; Charbonneau, D.; Erb, J.; Kochanowski, B.; Beuerman, D.; Poehner, R.; Bruce, A.W. Oral use of Lactobacillus rhamnosus GR-1 and L. fermentum RC-14 significantly alters vaginal flora: Randomized, placebo-controlled trial in 64 healthy women. FEMS Microbiol. Immunol. 2003, 35, 131–134. [Google Scholar] [CrossRef] [Green Version]
- Alberti, D.D.; Russo, R.; Terruzzi, F.; Nobile, V.; Ouwehand, A.C. Lactobacilli vaginal colonization after oral consumption of Respecta® complex: A randomized controlled pilot study. Arch. Gynecol. Obstet. 2015, 292, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Paek, N.S.; Lee, Y.Y.; Han, S.H.; Kang, C.H.; So, J.S. Characterization and inhibitory activity of Lactobacillus plantarum MG 989 and Lactobacillus fermentum MG901 isolated from vaginal microbiota of Korean women against Gardnerella vaginalis and Candida albicans. Korean Soc. Biotechnol. Bioeng. J. 2016, 31, 40–45. [Google Scholar]
- Jung, Y.; Kang, C.H.; Shin, Y.J.; So, J.S. Characterization and antifungal activity against Candida albicans of vaginal Lactobacillus spp. isolated from Korean women. Korean Soc. Biotechnol. Bioeng. J. 2017, 32, 146–152. [Google Scholar]
- Kang, C.H.; Kim, Y.G.; Han, S.H.; Kim, J.S.; Paek, N.S.; So, J.S. In vitro probiotic properties of vaginal Lactobacillus fermentum MG901 and Lactobacillus plantarum MG989 against Candia albicans. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 228, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.H.; Han, S.H.; Kim, Y.G.; Paek, N.S.; So, J.S. In vitro probiotic properties of Lactobacillus salivarius MG242 isolated from human vagianl. Probiotics Antimicrob. Proteins 2018, 10, 343–349. [Google Scholar] [CrossRef]
- Kim, Y.G.; Han, S.H.; Kim, J.S.; Jeong, Y.; Paek, N.S.; Kang, C.H. Probiotic properties and inhibitory activity of lactic acid bacteria isolated from vaginal microbiota of Korean women against Gardnerella vaginalis and Candia albicans. Korean Soc. Biotechnol. Bioeng. J. 2019, 34, 91–98. [Google Scholar]
- Kim, H.; Kim, Y.G.; Kang, C.H. In vivo confirmation of the antimicrobial effect of probiotic candidates against Gardnerella vaginalis. Microorganisms 2021, 9, 1690. [Google Scholar] [CrossRef] [PubMed]
- Won, G.; Choi, S.I.; Kang, C.H.; Kim, G.H. Lactiplantibacillus plantarum MG4296 and Lacticaseibacillus paracasei MG5012 ameliorates insulin resistance in palmitic acid-induced HepG2 cells and high fat diet-induced mice. Microorganisms 2021, 9, 1139. [Google Scholar] [CrossRef] [PubMed]
- McGroarty, J.A.; Tomeczek, L.; Pond, D.G.; Reid, G.; Bruce, A.W. Hydrogen peroxide production by Lactobacillus species: Correlation with susceptibility to the spermicidal compound nonoxynol-9. J. Infect. Dis. 1992, 165, 1142–1144. [Google Scholar] [CrossRef]
- Joo, H.M.; Hyun, Y.J.; Myoung, K.S.; Ahn, Y.T.; Lee, J.H.; Huh, C.S.; Han, M.J.; Kim, D.H. Lactobacillus johnsonii HY7042 ameliorates Gardnerella vaginalis-induced vaginosis by killing Gardnerella vaginalis and inhibiting NF-κB activation. Int. Immunopharmacol. 2011, 11, 1758–1765. [Google Scholar] [CrossRef]
- Gilbert, N.M.; Lewis, W.G.; Lewis, A.L. Clinical features of bacterial vaginosis in a murine model of vaginal infection with Gardnerella vaginalis. PLoS One 2013, 8, e5959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.E.; Jeong, J.J.; Choi, S.Y.; Kim, H.; Han, M.J.; Kim, D.H. Lactobacillus rhamnosus HN001 and Lactobacillus acidophilus GLa-14 attenuate Gardnerella vaginalis-infected bacterial vaginosis in mice. Nutrients 2017, 9, 531. [Google Scholar] [CrossRef] [Green Version]
- Daniel, M.; Pascual, L.; Barberis, L. Curative effect of the probiotic strain Lactobacillus fermentum L23 in a murine model of vaginal infection by Gardnerella vaginalis. Lett. Appl. Microbiol. 2014, 59, 93–98. [Google Scholar] [CrossRef]
- Sulin, C.; How, C.B.; Jamil, A.A.M.; Yih, C.S.; Meleh, H.U.; Lung, L.T.T. Characterisation of the probiotic qualities exhibited by Lactobacilli strains isolated from the anogenital tract. Malaysian J. Med. Health Sci. 2019, 15, 37–45. [Google Scholar]
- Damke, E.; Tsuzuki, J.K.; Chassot, F.; Cortez, D.A.; Ferreira, I.C.; Mesquita, C.S.; da-Silva, V.R.; Svidzinski, T.I.; Consolaro, M.E. Spermicidal and anti-Trichomonas vaginalis activity of Brazilian Sapindus saponaria. BMC complement. med. ther. 2013, 13, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, A.K.; Pottinger, P.S. Management of urinary tract infections in the era of increasing antimicrobial resistance. Med. Clin. North Am. 2013, 97, 737–757. [Google Scholar] [CrossRef] [PubMed]
- Schwebke, J.R.; Muzny, C.A.; Josey, W.E. Role of Gardnerella vaginalis in the pathogenesis of bacterial vaginosis: A conceptual model. J. Infect. Dis. 2014, 210, 338–343. [Google Scholar] [CrossRef]
- Morrill, S.; Gilbert, N.M.; Lewis, A.L. Gardnerella vaginalis as a cause of bacterial vaginosis: Appraisal of the evidence from in vivo models. Front. Cell. Infect. Microbiol. 2020, 10, 168. [Google Scholar] [CrossRef]
- Gillor, O.; Nigro, L.M.; Riley, M.A. Genetically engineered bacteriocins and their potential as the next generation of antimicrobials. Curr. Pharm. Des. 2005, 11, 1067–1075. [Google Scholar] [CrossRef] [Green Version]
- Macklaim, J.M.; Clemente, J.C.; Knight, R.; Gloor, G.B.; Reid, G. Changes in vaginal microbiota following antimicrobial and probiotic therapy. Microb. ecol. health dis. 2015, 26, 27799. [Google Scholar] [CrossRef] [PubMed]
- Soubhye, J.; Aldib, I.; Delporte, C.; Prévost, M.; Dufrasne, F.; Antwerpen, P.V. Myeloperoxidase as a target for the treatment of inflammatory syndromes: Mechanisms and structure activity relationships of inhibition. Curr. Med. Chem. 2016, 23, 3975–4008. [Google Scholar] [CrossRef]
- Trifonova, R.T.; Bajpai, M.; Pasicznyk, J.; Chandra, N.; Doncel, G.F.; Fichorova, R.N. Biomarkers of leukocyte traffic and activation in the vagianl mucosa. Biomarkers 2007, 12, 608–622. [Google Scholar] [CrossRef]
- Turovskiy, Y.; Noll, K.S.; Chikindas, M.L. The etiology of bacterial vaginosis. J. Appl. Microbiol. 2011, 110, 1105–1128. [Google Scholar] [CrossRef] [PubMed]
- Chee, W.J.Y.; Chew, S.Y.; Than, L.T.L. Vaginal microbiota and the potential of Lactobacillus derivatives in maintaining vaginal health. Microb. Cell Fact. 2020, 19, 203. [Google Scholar] [CrossRef]
- Homayouni, A.; Bastani, P.; Ziyadi, S.; Mohammad-Alizadeh-Charandabi, S.; Ghalibaf, M.; Mortazavian, A.M.; Mehrabany, E.V. Effects of Probiotics on the recurrence of bacterial vaginosis: A review. J. Low. Genit. Tract Dis. 2013, 18, 79–86. [Google Scholar] [CrossRef]
- Bradshaw, C.S.; Morton, A.N.; Hocking, J.; Garland, S.M.; Morris, M.B.; Moss, L.M.; Horvath, L.B.; Kuzevska, I.; Fairley, C.K. High recurrence rates of bacterial vaginosis over the course of 12 months after oral metronidazole therapy and factors associated with recurrence. J. Infect. Dis. 2006, 193, 1478–1486. [Google Scholar] [CrossRef] [Green Version]
- Machado, A.; Cerca, N. Influence of biofilm formation by Gardnerella vaginalis and other anaerobes on bacterial vaginosis. J. Infect. Dis. 2015, 212, 1856–1861. [Google Scholar] [CrossRef] [Green Version]
- Kumherová, M.; Veselá, K.; Kosová, M.; Mašata, J.; Horáčková, Š.; & Šmidrkal, J. Novel potential probiotic Lactobacilli for prevention and treatment of vulvovaginal infections. Probiotics Antimicrob. Proteins 2021, 13, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Atassi, F.; Brassart, D.; Grob, P.; Graf, F.; Servin, A.L. Lactobacillus strains isolated from the vaginal microbiota of healthy women inhibit Prevotella bivia and Gardnerella vaginalis in coculture and cell culture. FEMS Immunol. Med. Microbiol. 2006, 48, 424–432. [Google Scholar] [CrossRef] [Green Version]
- Sethi, S.; Das, A.; Sharma, M. Inhibition of Gardnerella vaginalis by lactobacilli. Int. J. Gynecol. Obstet. 2006, 93, 158–159. [Google Scholar] [CrossRef] [PubMed]
- Atassi, F.; Servin, A.L. Individual and co-operative roles of lactic acid and hydrogen peroxide in the killing activity of enteric strain Lactobacillus johnsonii NCC933 and vaginal strain Lactobacillus gasseri KS120.1 against enteric, uropathogenic and vaginosis-associated pathogens. FEMS Microbiol. Lett. 2010, 304, 29–38. [Google Scholar] [PubMed]
- Dover, S.E.; Aroutcheva, A.A.; Faro, S.; Chikindas, M.L. Natural antimicrobials and their role in vaginal health: A short review. Int. J. Probiotics Prebiotics 2008, 3, 219–230. [Google Scholar]
- Bang, J.H.; Shin, H.J.; Choi, H.J.; Kim, D.W.; Ahn, C.S.; Jeong, Y.K.; Joo, W.H. Probiotic potential of Lactobacillus isolates. J. Life Sci. 2012, 22, 251–258. [Google Scholar] [CrossRef]
- Martin, R.; Martin, C.; Escobedo, S.; Suarez, J.E.; Quiros, L.M. Surface glycosaminoglycans mediate adherence between HeLa cells and Lactobacillus salivarius Lv72. BMC Microbiol. 2013, 13, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coudeyras, S.; Jugie, G.; Vermerie, M.; Forestier, C. Adhesion of human probiotic Lactobacillus rhamnosus to cervical and vaginal cells and interaction with vaginosis-associated pathogens. Infect. Dis. Obstet. Gynecol. 2008, 2008, 549640. [Google Scholar] [CrossRef] [Green Version]
- Qian, Z.; Zhu, H.; Zhao, D.; Yang, P.; Gao, F.; Lu, C.; Lu, C.; Yin, Y.; Kan, S.; Chen, D. Probiotic Lactobacillus sp. strains inhibit growth, adhesion, biofilm formation, and gene expression of bacterial vaginosis-inducing Gardnerella vaginalis. Microorganisms 2021, 9, 728. [Google Scholar] [CrossRef] [PubMed]
- Hardy, L.; Jespers, V.; Van den Bulck, M.; Buyze, J.; Mwambarangwe, L.; Musengamana, V.; Vaneechoutte, M.; Crucitti, T. The presence of the putative Gardnerella vaginalis sialidase A gene in vaginal specimens is associated with bacterial vaginosis biofilm. PLoS One 2017, 12, e0172522. [Google Scholar] [CrossRef] [Green Version]
- Giraldo, P.C.; de Carvalho, J.B.J.; do Amaral, R.L.G.; da Silveira Gonçalves, A.K.; Eleutério Jr, J.; Guimarães, F. Identification of immune cells by flow cytometry in vaginal lavages from women with vulvovaginitis and normal microbiota. Am. J. Reprod. Immunol. 2012, 67, 198–205. [Google Scholar] [CrossRef]
- Mitchell, C.; Marrazzo, J. Bacterial vaginosis and the cervicovaginal immune response. Am. J. Reprod. Immunol. 2014, 71, 555–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cauci, S.; Culhane, J.F.; di Santolo, M.; McCollum, K. Among pregnant women with bacterial vaginosis, the hydrolytic enzymes sialidase and prolidase are positively associated with interleukin-1beta. Am. J. Obstet. Gynecol. 2008, 198, 132.e1–e7. [Google Scholar] [CrossRef]
- Muzny, C.A.; Łaniewski, P.; Schwebke, J.R.; Herbst-Kralovetz, M.M. Host-vaginal microbiota interactions in the pathogenesis of bacterial vaginosis. Curr. Opin. Infect. Dis. 2020, 33, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Marrs, C.N.; Knobel, S.M.; Zhu, W.Q.; Sweet, S.D.; Chaudhry, A.R.; Alcendor, D.J. Evidence for Gardnerella vaginalis uptake and internalization by squamous vaginal epithelial cells: Implications for the pathogenesis of bacterial vaginosis. Microbes Infect. 2012, 14, 500–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemalatha, R.; Ramalaxmi, B.A.; KrishnaSwetha, G.; Kumar, P.U.; Rao, D.M.; Balakrishna, N.; Annapurna, V. Cervico-vaginal inflammatory cytokines and sphingomyelinase in women with and without bacterial vaginosis. Am. J. Med. Sci. 2012, 344, 35–39. [Google Scholar] [CrossRef]
- Wasiela, M.; Krzemiński, Z.; Kalinka, J.; Brzezińska-Błaszczyk, E. Correlation between levels of selected cytokines in cervico-vaginal fluid of women with abnormal vaginal bacterial flora. Med Dosw Mikrobiol. 2005, 57, 327–333. [Google Scholar] [PubMed]
- Santos, C.M.A.; Pires, M.C.V.; Leão, T.L.; Silva, A.K.S.; Miranda, L.S.; Martins, F.S.; Silva, A.M.; Nicoli, J.R. Anti-inflammatory effect of two Lactobacillus strains during infection with Gardnerella vaginalis and Candida albicans in a HeLa cell culture model. Microbiology 2018, 164, 349–358. [Google Scholar] [CrossRef]
- Swidsinski, A.; Mendling, W.; Loening-Baucke, V.; Ladhoff, A.; Swidsinski, S.; Hale, L.P.; Lochs, H. Adherent biofilms in bacterial vaginosis. Obstet. Gynecol. 2005, 106, 1013–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | pH | H2O2 Content (g/L) | Lactic Acid Content D (−) + L (+)(g/L) | N-9 Susceptibility (+/−) |
|---|---|---|---|---|
| Ligilactobacillus salivarius MG242 | 3.77 ± 0.02 | 1.23 ± 0.00 | 17.44 ± 1.1 | - |
| Limosilactobacillusfermentum MG901 | 4.32 ± 0.02 | 1.93 ± 0.08 | 11.83 ± 1.8 | - |
| Lactiplantibacillus plantarum MG989 | 3.76 ± 0.01 | 0.96 ± 0.09 | 11.83 ± 0.9 | - |
| Lacticaseibacillus paracasei MG4272 | 3.78 ± 0.01 | 0.80 ± 0.06 | 18.76 ± 2.4 | - |
| Lacticaseibacillus rhamnosus MG4288 | 3.83 ± 0.01 | 0.78 ± 0.05 | 15.43 ± 1.0 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, S.-I.; Won, G.; Kim, Y.; Kang, C.-H.; Kim, G.-H. Lactobacilli Strain Mixture Alleviates Bacterial Vaginosis through Antibacterial and Antagonistic Activity in Gardnerella vaginalis-Infected C57BL/6 Mice. Microorganisms 2022, 10, 471. https://doi.org/10.3390/microorganisms10020471
Choi S-I, Won G, Kim Y, Kang C-H, Kim G-H. Lactobacilli Strain Mixture Alleviates Bacterial Vaginosis through Antibacterial and Antagonistic Activity in Gardnerella vaginalis-Infected C57BL/6 Mice. Microorganisms. 2022; 10(2):471. https://doi.org/10.3390/microorganisms10020471
Chicago/Turabian StyleChoi, Soo-Im, GaYeong Won, YongGyeong Kim, Chang-Ho Kang, and Gun-Hee Kim. 2022. "Lactobacilli Strain Mixture Alleviates Bacterial Vaginosis through Antibacterial and Antagonistic Activity in Gardnerella vaginalis-Infected C57BL/6 Mice" Microorganisms 10, no. 2: 471. https://doi.org/10.3390/microorganisms10020471
APA StyleChoi, S. -I., Won, G., Kim, Y., Kang, C. -H., & Kim, G. -H. (2022). Lactobacilli Strain Mixture Alleviates Bacterial Vaginosis through Antibacterial and Antagonistic Activity in Gardnerella vaginalis-Infected C57BL/6 Mice. Microorganisms, 10(2), 471. https://doi.org/10.3390/microorganisms10020471