
Herpesvirus Infections in KIR2DL2-Positive Multiple Sclerosis Patients: Mechanisms Triggering Autoimmunity

 ,
,  , , , ,
, , , ,  , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Quantification of Peripheral Blood Antibodies
2.3. Genotyping of KIR and HLA
2.4. Quantification of Cytokine Levels
2.5. Peripheral Blood Mononuclear Cell (PBMC) Culture
2.6. Peripheral Blood Monocyte-Derived Microglia
2.7. HHV Infection
2.8. Viral Load Quantification
2.9. Cytometric Analysis and CD107a Degranulation Assay
2.10. Statistical Analyses
3. Results
3.1. KIR/HLA Typing
3.2. Microglial Cells-NK Cells Co-Culture during HHVs’ Infection
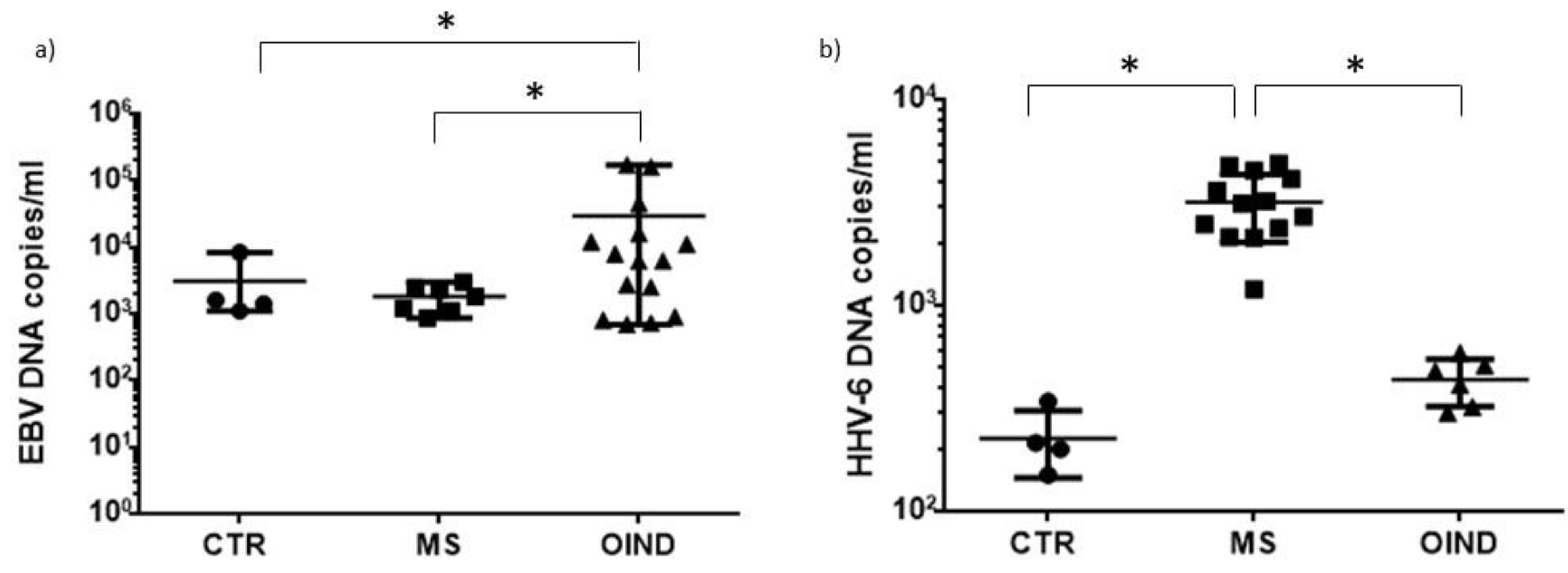
3.3. Levels of HHVs’ DNA in Peripheral Blood
3.4. Levels of Antibodies towards HHVs
3.5. Levels of Inflammatory Cytokines in Plasma Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, W.W.; Zhang, X.; Huang, W.J. Role of neuroinflammation in neurodegenerative diseases. Mol. Med. Rep. 2016, 13, 3391–3396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamza, T.H.; Zabetian, C.P.; Tenesa, A.; Laederach, A.; Montimurro, J.; Yearout, D.; Kay, D.M.; Doheny, K.F.; Paschall, J.; Pugh, E.; et al. Common genetic variation in the HLA region is associated with late-onset sporadic Parkinson’s disease. Nat. Genet 2010, 42, 781–785. [Google Scholar] [CrossRef] [PubMed]
- Bar-Or, A.; Pender, M.P.; Khanna, R.; Steinman, L.; Hartung, H.P.; Maniar, T.; Croze, E.; Aftab, B.T.; Giovannoni, G.; Joshi, M.A.; et al. Epstein–Barr Virus in Multiple Sclerosis: Theory and Emerging Immunotherapies. Trends Mol. Med. 2020, 26, 296–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Lafuente, R.; De Las Heras, V.; Bartolomé, M.; García-Montojo, M.; Arroyo, R. Human herpesvirus 6 and multiple sclerosis: A one-year follow-up study. Brain Pathol. 2006, 16, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Opsahl, M.L.; Kennedy, P.G.E. Early and late HHV-6 gene transcripts in multiple sclerosis lesions and normal appearing white matter. Brain 2005, 128, 516–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komaroff, A.L.; Pellett, P.E.; Jacobson, S. Human herpesviruses 6A and 6B in brain diseases: Association versus causation. Clin. Microbiol. Rev. 2020, 34, 34. [Google Scholar] [CrossRef]
- Caselli, E.; Boni, M.; Bracci, A.; Rotola, A.; Cermelli, C.; Castellazzi, M.; Di Luca, D.; Cassai, E. Detection of antibodies directed against human herpesvirus 6 U94/REP in sera of patients affected by multiple sclerosis. J. Clin. Microbiol. 2002, 40, 4131–4137. [Google Scholar] [CrossRef] [Green Version]
- Gross, C.C.; Schulte-Mecklenbeck, A.; Wiendl, H.; Marcenaro, E.; Kerlero de Rosbo, N.; Uccelli, A.; Laroni, A. Regulatory Functions of Natural Killer Cells in Multiple Sclerosis. Front Immunol. 2016, 7, 606. [Google Scholar] [CrossRef] [Green Version]
- Campbell, K.S.; Purdy, A.K. Structure/Function of Human Killer Cell Immunoglobulin-Like Receptors: Lessons from Polymorphisms, Evolution, Crystal Structures and Mutations. Immunology 2011, 132, 315–325. [Google Scholar] [CrossRef]
- Pende, D.; Falco, M.; Vitale, M.; Cantoni, C.; Vitale, C.; Munari, E.; Bertaina, A.; Moretta, F.; Del Zotto, G.; Pietra, G.; et al. Killer Ig-Like Receptors (Kirs): Their Role in Nk Cell Modulation and Developments Leading to Their Clinical Exploitation. Front Immunol. 2019, 10, 1179. [Google Scholar] [CrossRef] [Green Version]
- Rezaei, R.; Mostafaei, S.; Aslani, S.; Jamshidi, A.; Mahmoudi, M. Association study between killer immunoglobulin-like receptor polymorphisms and ankylosing spondylitis disease: An updated meta-analysis. Int. J. Rheum. Dis. 2018, 21, 1746–1755. [Google Scholar] [CrossRef] [PubMed]
- Fathollahi, A.; Aslani, S.; Mostafaei, S.; Rezaei, N.; Mahmoudi, M.J.I.R. The role of killer-cell immunoglobulin-like receptor (KIR) genes in susceptibility to inflammatory bowel disease: Systematic review and meta-analysis. BMC Med. Genet 2019, 20, 24. [Google Scholar] [CrossRef] [PubMed]
- Aghaei, H.; Mostafaei, S.; Aslani, S.; Jamshidi, A.; Mahmoudi, M. Association study between KIR polymorphisms and rheumatoid arthritis disease: An updated meta-analysis. BMC Med. Genet 2019, 20, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, R.; Gentili, V.; Casetta, I.; Caselli, E.; De Gennaro, R.; Granieri, E.; Cassai, E.; Di Luca, D.; Rotola, A. Altered natural killer cells’ response to herpes virus infection in multiple sclerosis involves KIR2DL2 expression. J. Neuroimmunol. 2012, 251, 55–64. [Google Scholar] [CrossRef]
- Rizzo, R.; Bortolotti, D.; Fainardi, E.; Gentili, V.; Bolzani, S.; Baldi, E.; Casetta, I.; Granieri, E.; Rotola, A.; Furlan, R.; et al. KIR2DL2 inhibitory pathway enhances Th17 cytokine secretion by NK cells in response to herpesvirus infection in multiple sclerosis patients. J. Neuroimmunol. 2016, 294, 1–5. [Google Scholar] [CrossRef]
- Ben Fredj, N.; Rizzo, R.; Bortolotti, D.; Nefzi, F.; Chebel, S.; Rotola, A.; Frih-Ayed, M.; Di Luca, D.; Aouni, M. Evaluation of the implication of KIR2DL2 receptor in multiple sclerosis and herpesvirus susceptibility. J. Neuroimmunol. 2014, 271, 30–35. [Google Scholar] [CrossRef]
- Estefanía, E.; Gómez-Lozano, N.; Portero, F.; de Pablo, R.; Solís, R.; Sepúlveda, S.; Vaquero, M.; González, M.A.; Suárez, E.; Roustán, G.; et al. Influence of KIR gene diversity on the course of HSV-1 infection: Resistance to the disease is associated with the absence of KIR2DL2 and KIR2DS2. Tissue Antigens 2007, 70, 34–41. [Google Scholar] [CrossRef]
- McDonald, W.I.; Compston, A.; Edan, G.; Goodkin, D.; Hartung, H.-P.; Lublin, F.D.; McFarland, H.F.; Paty, D.W.; Polman, C.H.; Reingold, S.C.; et al. Recommended diagnostic criteria for multiple sclerosis: Guidelines from the International Panel on the Diagnosis of Multiple Sclerosis. Ann. Neurol. 2001, 50, 121–127. [Google Scholar] [CrossRef]
- Kurtzke, J.F. Rating neurological impairment in multiple sclerosis: An expanded disability scale (EDSS). Neurology 1983, 33, 1444–1452. [Google Scholar] [CrossRef] [Green Version]
- Lublin, D.F.; Reingold, S.C. Defining the clinical course of multiple sclerosis: Results of an international survey. Neurology 1996, 46, 907–911. [Google Scholar] [CrossRef]
- Hochberg, M.C. Updating the American College of Rheumatology revised criteria for the classification of systemic lupus erythematosus. Arthritis Care Res. 1997, 40, 1725. [Google Scholar] [CrossRef] [PubMed]
- Alunno, A.; Najm, A.; Machado, P.M.; Bertheussen, H.; Burmester, G.-R.R.; Carubbi, F.; De Marco, G.; Giacomelli, R.; Hermine, O.; Isaacs, J.D.; et al. 2021 update of the EULAR points to consider on the use of immunomodulatory therapies in COVID-19. Ann. Rheum. Dis. 2021, 81, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Bortoluzzi, A.; Fanouriakis, A.; Appenzeller, S.; Costallat, L.; Scirè, C.A.; Murphy, E.; Bertsias, G.; Hanly, J.; Govoni, M. Validity of the Italian algorithm for the attribution of neuropsychiatric events in systemic lupus erythematosus: A retrospective multicentre international diagnostic cohort study. BMJ Open 2017, 7, e015546. [Google Scholar] [CrossRef] [PubMed]
- Bortolotti, D.; Gentili, V.; Caselli, E.; Sicolo, M.; Soffritti, I.; D’Accolti, M.; Barao, I.; Rotola, A.; Di Luca, D.; Rizzo, R. DNA Sensors’ Signaling in NK Cells During HHV-6A, HHV-6B and HHV-7 Infection. Front Microbiol. 2020, 11, 226. [Google Scholar] [CrossRef] [Green Version]
- Leone, C.; Le Pavec, G.; Même, W.; Porcheray, F.; Samah, B.; Dormont, D.; Gras, G. Characterization of human monocyte-derived microglia-like cells. Glia 2006, 54, 183. [Google Scholar] [CrossRef]
- Bortolotti, D.; Gentili, V.; Rotola, A.; Caselli, E.; Rizzo, R. HHV-6A infection induces amyloid-beta expression and activation of microglial cells. Alzheimers Res. Ther. 2019, 11, 104. [Google Scholar] [CrossRef] [Green Version]
- Bortolotti, D.; Gentili, V.; Rotola, A.; Cultrera, R.; Marci, R.; Di Luca, D.; Rizzo, R. HHV-6A infection of endometrial epithelial cells affects immune profile and trophoblast invasion. Am. J. Reprod. Immunol. 2019, 82, e13174. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Song, F.; Fernandez-Escobar, A.; Luo, G.; Wang, J.; Sun, Y. The Properties of Cytokines in Multiple Sclerosis: Pros and Cons. Am. J. Med. Sci. 2018, 356, 552–560. [Google Scholar] [CrossRef]
- Trysberg, E.; Carlsten, H.; Tarkowski, A. Intrathecal cytokines in systemic lupus erythematosus with central nervous system involvement. Lupus 2000, 9, 498–503. [Google Scholar] [CrossRef]
- Caselli, E.; Di Luca, D. Molecular biology and clinical associations of Roseoloviruses human herpesvirus 6 and human herpesvirus 7. New Microbiol. 2007, 30, 173–187. [Google Scholar]
- Henle, W.; Henle, G. Seroepidemiology of the virus. In The Epstein-Barr Virus; Epstein, M.A., Achong, B.G., Eds.; Springer: Berlin/Heidelberg, Germany, 1979; pp. 61–78. [Google Scholar]
- Donati, D.; Akhyani, N.; Fogdell-Hahn, A.; Cermelli, C.; Cassiani-Ingoni, R.; Vortmeyer, A.; Heiss, J.D.; Cogen, P.; Gaillard, W.D.; Sato, S.; et al. Detection of human herpesvirus-6 in mesial temporal lobe epilepsy surgical brain resections. Neurology 2003, 61, 1405–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harberts, E.; Yao, K.; Wohler, J.E.; Maric, D.; Ohayon, J.; Henkin, R.; Jacobson, S. Human herpesvirus-6 entry into the central nervous system through the olfactory pathway. Proc. Natl. Acad. Sci. USA 2011, 108, 13734–13739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynaud, J.M.; Jégou, J.; Welsch, J.C.; Horvat, B. Human Herpesvirus 6A Infection in CD46 Transgenic Mice: Viral Persistence in the Brain and Increased Production of Proinflammatory Chemokines via Toll-Like Receptor 9. J. Virol. 2014, 88, 5421–5436. [Google Scholar] [CrossRef] [Green Version]
- Leibovitch, E.; Wohler, J.E.; Macri, S.M.C.; Motanic, K.; Harberts, E.; Gaitán, M.I.; Maggi, P.; Ellis, M.; Westmoreland, S.; Silva, A.; et al. Novel marmoset (Callithrix jacchus) model of human Herpesvirus 6A and 6B infections: Immunologic, virologic and radiologic characterization. PLoS Pathog. 2013, 9, e1003138. [Google Scholar] [CrossRef] [PubMed]
- Castellazzi, M.; Delbue, S.; Elia, F.; Gastaldi, M.; Franciotta, D.; Rizzo, R.; Bellini, T.; Bergamaschi, R.; Granieri, E.; Fainardi, E. Epstein-Barr Virus Specific Antibody Response in Multiple Sclerosis Patients during 21 Months of Natalizumab Treatment. Dis. Markers 2015, 2015, 901312. [Google Scholar] [CrossRef]
- Mechelli, R.; Manzari, C.; Policano, C.; Annese, A.; Picardi, E.; Umeton, R.; Fornasiero, A.; D’Erchia, A.M.; Buscarinu, M.C.; Agliardi, C.; et al. Epstein-Barr virus genetic variants are associated with multiple sclerosis. Neurology 2015, 84, 1362–1368. [Google Scholar] [CrossRef] [Green Version]
- Serafini, B.; Rosicarelli, B.; Franciotta, D.; Magliozzi, R.; Reynolds, R.; Cinque, P.; Andreoni, L.; Trivedi, P.; Salvetti, M.; Faggioni, A.; et al. Dysregulated Epstein-Barr virus infection in the multiple sclerosis brain. J. Exp. Med. 2007, 204, 2899–2912. [Google Scholar] [CrossRef]
- Willis, S.N.; Stadelmann, C.; Rodig, S.J.; Caron, T.; Gattenloehner, S.; Mallozzi, S.S.; Roughan, J.E.; Almendinger, S.E.; Blewett, M.M.; Brück, W.; et al. Epstein–Barr virus infection is not a characteristic feature of multiple sclerosis brain. Brain 2009, 132, 3318–3328. [Google Scholar] [CrossRef]
- Lindner, J.M.; Cornacchione, V.; Sathe, A.; Be, C.; Srinivas, H.; Riquet, E.; Leber, X.-C.; Hein, A.; Wrobel, M.B.; Scharenberg, M.; et al. Human Memory B Cells Harbor Diverse Cross-Neutralizing Antibodies against BK and JC Polyomaviruses. Immunity 2019, 50, 668–676.e5. [Google Scholar] [CrossRef] [Green Version]
- Gelfand, J.M.; Cree, B.A.C.; Hauser, S.L. Ocrelizumab and Other CD20 + B-Cell-Depleting Therapies in Multiple Sclerosis. Neurotherapeutics 2017, 4, 835–841. [Google Scholar] [CrossRef] [Green Version]
- Thacker, E.L.; Mirzaei, F.; Ascherio, A. Infectious mononucleosis and risk for multiple sclerosis: A meta-analysis. Ann. Neurol. 2006, 59, 499–503. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, T.R.; Rostgaard, K.; Nielsen, N.M.; Koch-Henriksen, N.; Haahr, S.; Sørensen, P.S.; Hjalgrim, H. Multiple sclerosis after infectious mononucleosis. Arch. Neurol. 2007, 64, 72–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Challoner, P.B.; Smith, K.T.; Parker, J.D.; MacLeod, D.L.; Coulter, S.N.; Rose, T.M.; Schultz, E.R.; Bennett, J.L.; Garber, R.L.; Chang, M.; et al. Plaque-associated expression of human herpesvirus 6 in multiple sclerosis. Proc. Natl. Acad. Sci. USA 1995, 92, 7440–7444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engdahl, E.; Gustafsson, R.; Huang, J.; Biström, M.; Lima Bomfim, I.; Stridh, P.; Khademi, M.; Brenner, N.; Butt, J.; Michel, A.; et al. Increased Serological Response Against Human Herpesvirus 6A Is Associated With Risk for Multiple Sclerosis. Front Immunol. 2019, 10, 2715. [Google Scholar] [CrossRef] [Green Version]
- Epstein, D.J.; Dunn, J.; Deresinski, S. Infectious Complications of Multiple Sclerosis Therapies: Implications for Screening, Prophylaxis, and Management. Open Forum Infect. Dis. 2018, 5, 174. [Google Scholar] [CrossRef] [Green Version]
- Vey, N.; Karlin, L.; Sadot-Lebouvier, S.; Broussais, F.; Berton-Rigaud, D.; Rey, J.; Charbonnier, A.; Marie, D.; André, P.; Paturel, C.; et al. A phase 1 study of lirilumab (antibody against killer immunoglobulin-like receptor antibody KIR2D.; IPH2102) in patients with solid tumors and hematologic malignancies. Oncotarget 2018, 9, 17675–17688. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KIR and HLA | MS (n = 100) | OIND (n = 100) | CNTR (n = 100) | p | pc |
|---|---|---|---|---|---|
| Activating KIR | |||||
| KIR2DS1 | 50 | 36 | 37 | 0.09 | |
| KIR2DS2 | 62 | 34 | 37 | 7.0 × 10−4 | 4.2 × 10−3 |
| KIR2DS3 | 36 | 29 | 28 | 0.28 | |
| KIR2DS4 | 87 | 84 | 83 | 0 | |
| KIR2DS5 | 45 | 31 | 30 | 0.04 | |
| KIR3DS1 | 12 | 32 | 33 | 7.0 × 10−4 | 4.2 × 10−3 |
| Inhibitory KIR | |||||
| KIR2DL1 | 89 | 87 | 86 | 0.67 | |
| KIR2DL2 | 62 | 38 | 36 | 4.0 × 10−4 | 3.2 × 10−3 |
| KIR2DL3 | 72 | 76 | 77 | 0.52 | |
| KIR2DL4 | 98 | 99 | 100 | 0 | |
| KIR2DL5 | 60 | 44 | 46 | 0.06 | |
| KIR3DL1 | 94 | 81 | 83 | 0.025 | |
| KIR3DL2 | 98 | 99 | 100 | 0 | |
| KIR3DL3 | 98 | 99 | 100 | 0 | |
| KIR genotype | |||||
| AA | 28 | 33 | 35 | 0.36 | |
| Bx | 72 | 66 | 65 | ||
| HLA genotype | |||||
| C1/C1 | 26 | 21 | 20 | 0.40 | |
| C1/C2 | 46 | 25 | 23 | 9.9 × 10−4 | 5.0 × 10−3 |
| C2/C2 | 29 | 44 | 43 | 0.055 | |
| HLA-Bw4 | 78 | 73 | 71 | 0 | |
| HLA-Bw6 | 92 | 82 | 84 | 0.13 | |
| KIR and their ligands | |||||
| KIR2DS1/KIR2DL1 present/HLA-C2 present | 40 | 27 | 28 | 0.1 | |
| KIR2DS1/KIR2DL1 present/HLA-C2 absent | 10 | 7 | 8 | 0.81 | |
| KIR2DS1/KIR2DL1 absent/HLA-C2 present | 2 | 1 | 1 | 0 | |
| KIR2DS2/KIR2DL2 present/HLA-C1 present | 48 | 27 | 28 | 0.00561 | 0.037 |
| KIR2DS2/KIR2DL2 present/HLA-C1 absent | 14 | 14 | 14 | 0 | |
| KIR2DS2/KIR2DL2 absent/HLA-C1 present | 6 | 32 | 31 | 6.4 × 10−6 | 5.8 × 10−5 |
| KIR3DS1/KI3DL1 present/HLA-Bw4 present | 30 | 24 | 25 | 0.53 | |
| KIR3DS1/KI3DL1 present/HLA-Bw4 absent | 9 | 6 | 7 | 0.80 | |
| KIR3DS1/KI3DL1 absent/HLA-Bw4 present | 1 | 2 | 3 | 0.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bortolotti, D.; Gentili, V.; Bortoluzzi, A.; Govoni, M.; Schiuma, G.; Beltrami, S.; Rizzo, S.; Baldi, E.; Caselli, E.; Pugliatti, M.; et al. Herpesvirus Infections in KIR2DL2-Positive Multiple Sclerosis Patients: Mechanisms Triggering Autoimmunity. Microorganisms 2022, 10, 494. https://doi.org/10.3390/microorganisms10030494
Bortolotti D, Gentili V, Bortoluzzi A, Govoni M, Schiuma G, Beltrami S, Rizzo S, Baldi E, Caselli E, Pugliatti M, et al. Herpesvirus Infections in KIR2DL2-Positive Multiple Sclerosis Patients: Mechanisms Triggering Autoimmunity. Microorganisms. 2022; 10(3):494. https://doi.org/10.3390/microorganisms10030494
Chicago/Turabian StyleBortolotti, Daria, Valentina Gentili, Alessandra Bortoluzzi, Marcello Govoni, Giovanna Schiuma, Silvia Beltrami, Sabrina Rizzo, Eleonora Baldi, Elisabetta Caselli, Maura Pugliatti, and et al. 2022. "Herpesvirus Infections in KIR2DL2-Positive Multiple Sclerosis Patients: Mechanisms Triggering Autoimmunity" Microorganisms 10, no. 3: 494. https://doi.org/10.3390/microorganisms10030494
APA StyleBortolotti, D., Gentili, V., Bortoluzzi, A., Govoni, M., Schiuma, G., Beltrami, S., Rizzo, S., Baldi, E., Caselli, E., Pugliatti, M., Castellazzi, M., Fernández, M., Fainardi, E., & Rizzo, R. (2022). Herpesvirus Infections in KIR2DL2-Positive Multiple Sclerosis Patients: Mechanisms Triggering Autoimmunity. Microorganisms, 10(3), 494. https://doi.org/10.3390/microorganisms10030494