
The “Dark Side” of Picocyanobacteria: Life as We Do Not Know It (Yet)


Abstract
:1. Introduction: Understanding Picocyanobacteria
2. The Genus Synechococcus
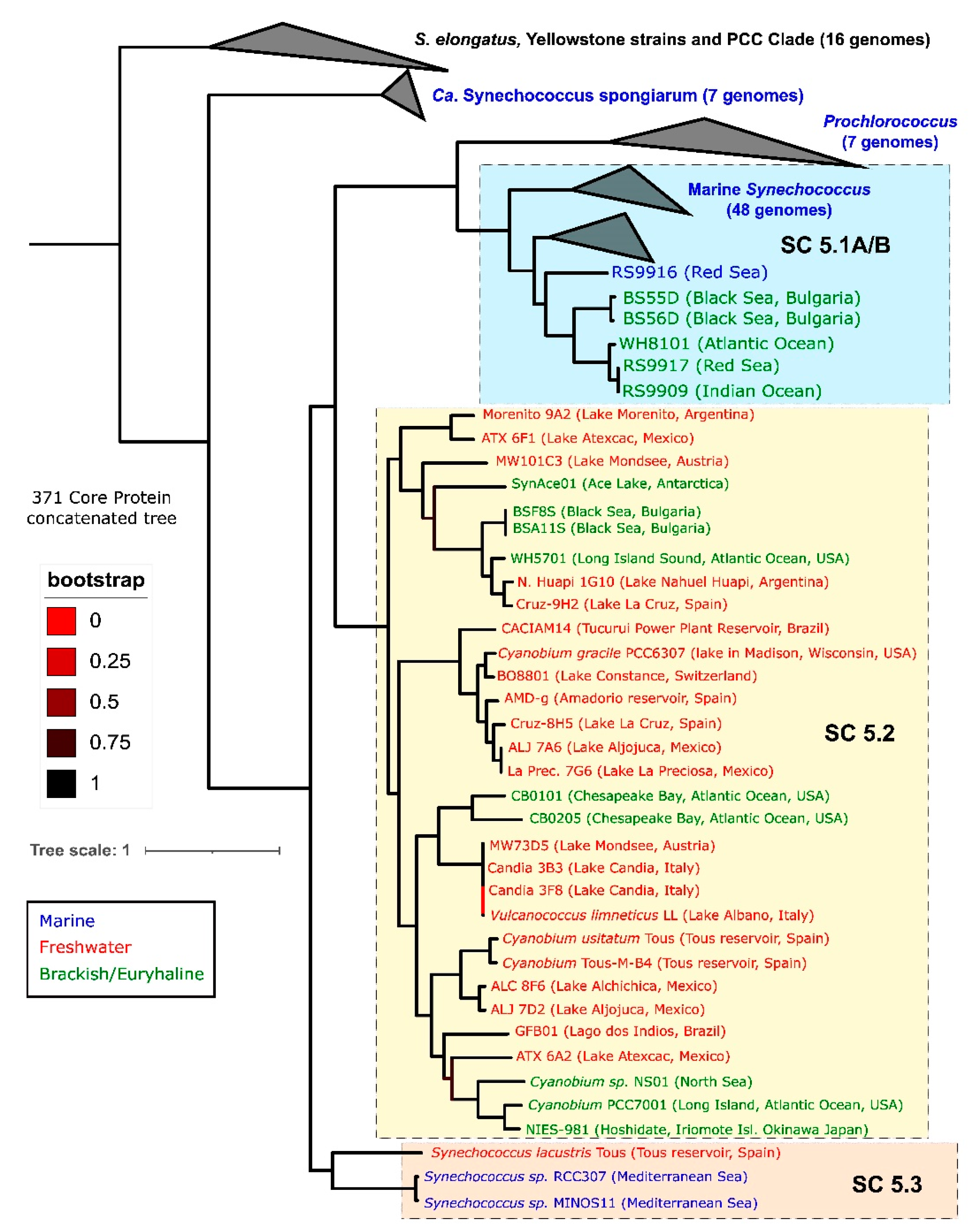
2.1. Phylogenetics: A Cryptic Genus?
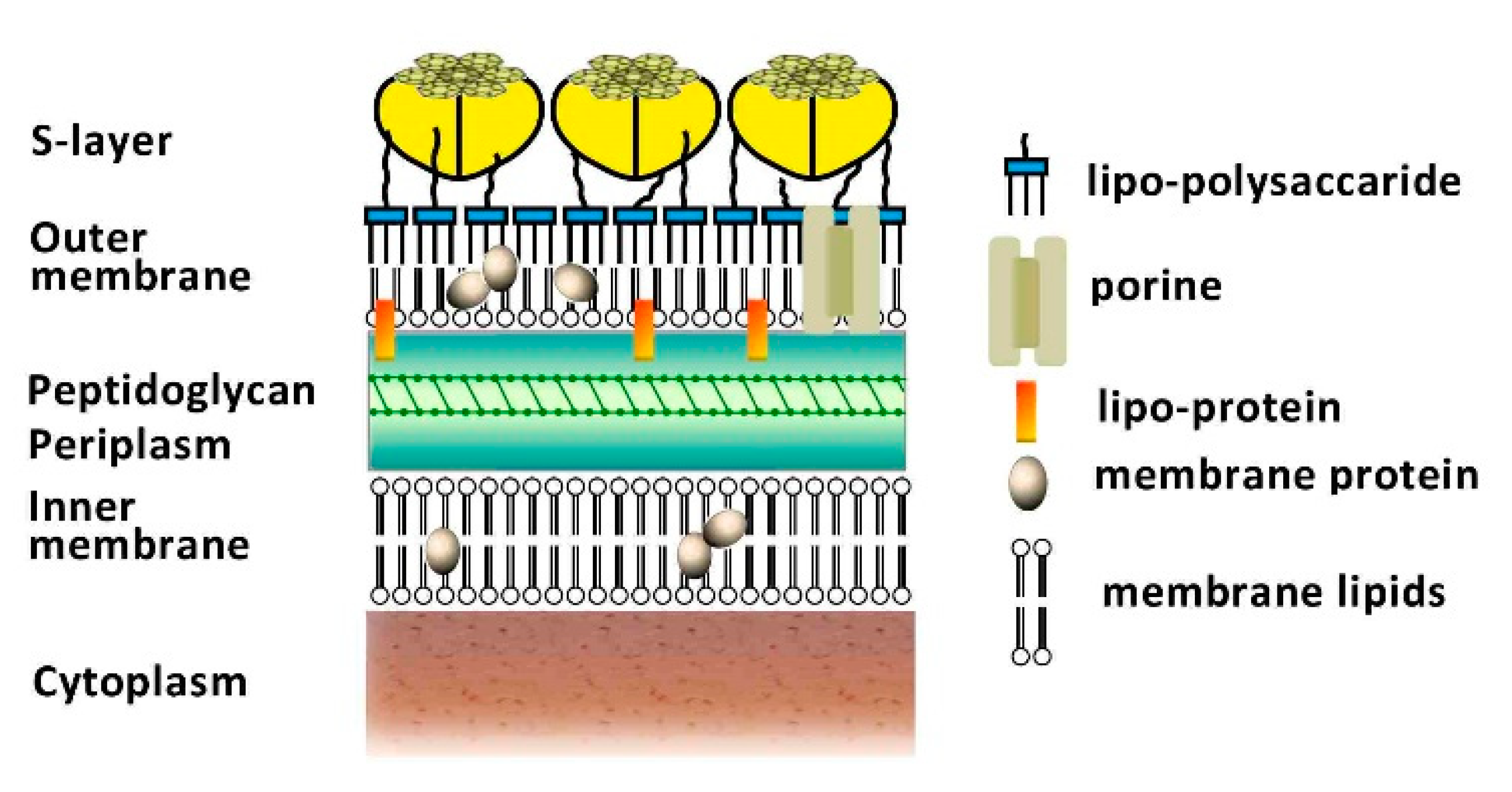
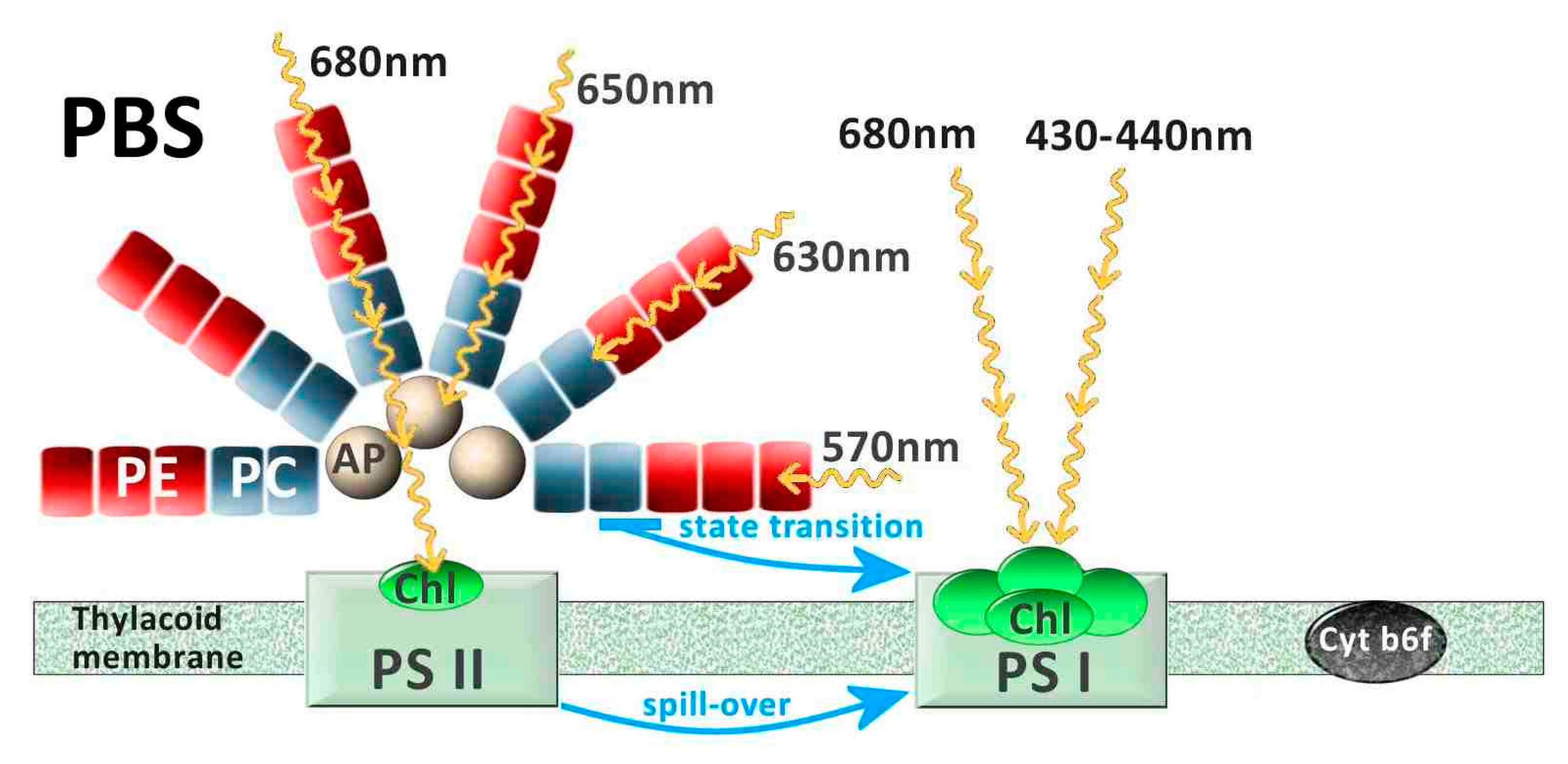
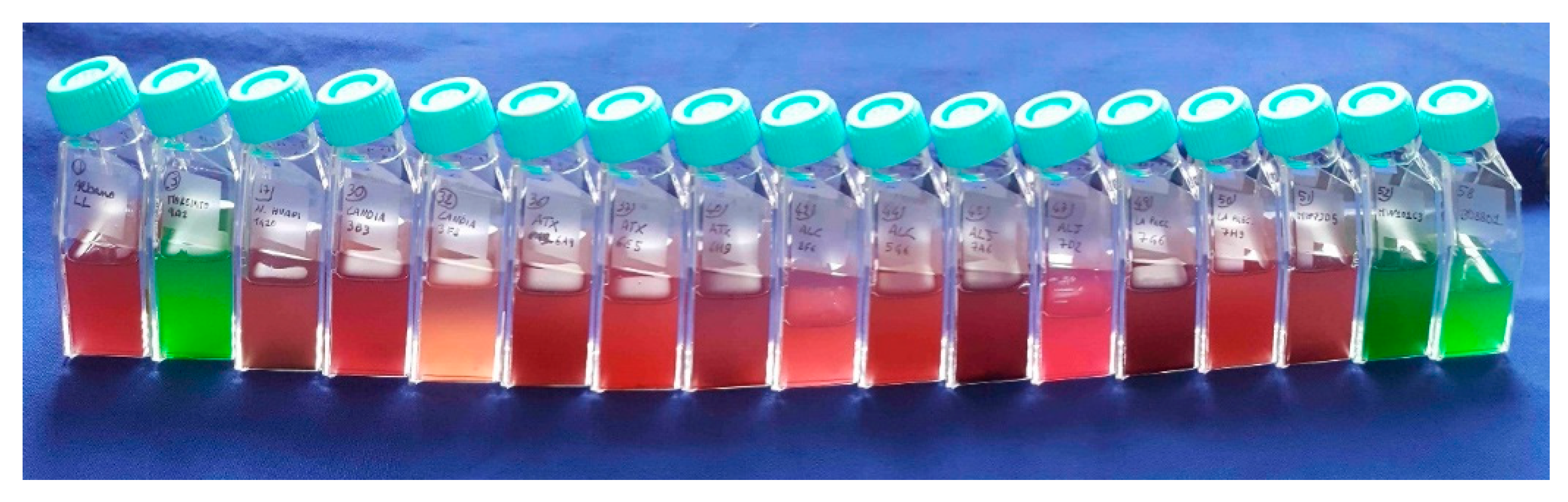
2.2. Physiological Features
2.3. Role and Distribution
2.3.1. Temperature
2.3.2. Nutrients
2.3.3. Primary Production
2.3.4. Community Composition
3. In the Dark: Synechococcus Surprising Adaptability
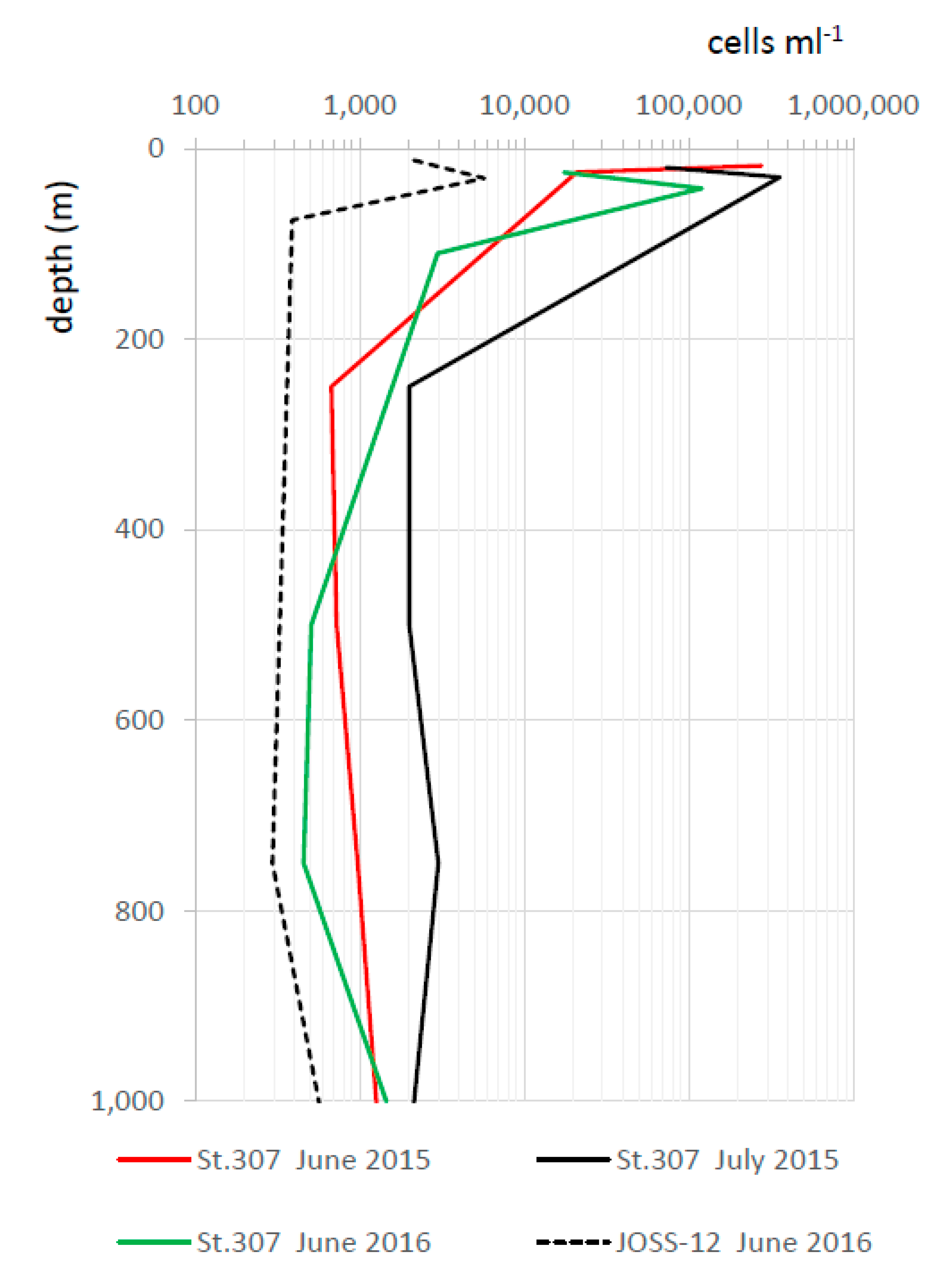
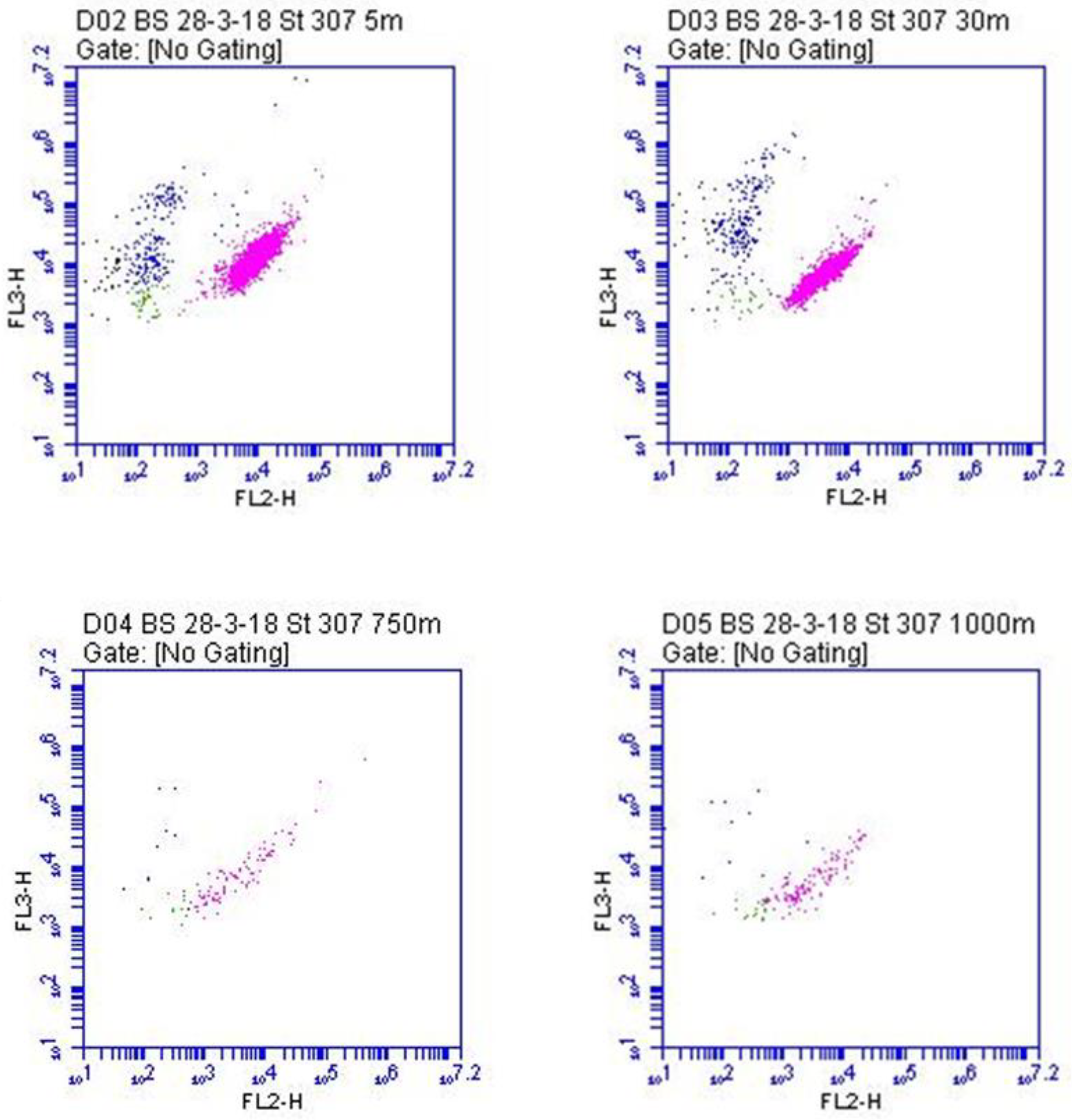
3.1. A Case Study: The Mesopelagic Zone of the Black Sea
3.2. Unexpected Metabolisms
3.3. Ecological Considerations
4. Conclusions: Learning from Life as We Do Not Know It
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnson, P.W.; Sieburth, J.M.N. Chrococcoid cyanobacteria in the sea: A ubiquitous and diverse phototrophic biomass. Limnol. Oceanogr. 1979, 24, 928–935. [Google Scholar] [CrossRef]
- Callieri, C. Synechococcus plasticity under environmental changes. FEMS Microbiol. Lett. 2017, 364, fnx229. [Google Scholar] [CrossRef] [PubMed]
- Callieri, C.; Cronberg, G.; Stockner, J.G. Freshwater picocyanobacteria- single cells, microcolonies and colonial forms. In Ecology of Cyanobacteria II: Their Diversity in Space and Time, 2nd ed.; Whitton, B., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 229–269. [Google Scholar]
- Scanlan, D.J. Marine Picocyanobacteria. In Ecology of Cyanobacteria II: Their Diversity in Space and Time, 2nd ed.; Whitton, B., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 503–533. [Google Scholar]
- Waterbury, J.B.; Watson, S.W.; Guillard, R.R.L.; Brand, L.E.E. Widespread occurrence of a unicellular, marine, planktonic cyanobacterium. Nature 1979, 227, 293–294. [Google Scholar] [CrossRef]
- Sánchez-Baracaldo, P. Origin of marine planktonic cyanobacteria. Sci. Rep. 2015, 5, 17418. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Baracaldo, P.; Bianchini, G.; Wilson, J.D.; Knoll, A.H. Cyanobacteria and biogeochemical cycles through Earth history. Trends Microbiol. 2022, 30, 143–157. [Google Scholar] [CrossRef]
- Archibald, J.M. Endosymbiosis and eukaryotic cell evolution. Curr. Biol. 2015, 25, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Honda, D.; Yokota, A.; Sugiyama, J. Detection of seven major evolutionary lineages in cyanobacteria based on 16S rRNA gene sequence analysis with new sequences of five marine Synechococcus strains. J. Mol. Evol. 1999, 48, 723–739. [Google Scholar] [CrossRef]
- Castenholz, R.W.; Phylum, B.X. Cyanobacteria, oxygenic photosynthetic bacteria. In Bergey’s Manual of Systematic Bacteriology; Boone, D.R., Castenholz, R.W., Eds.; Springer: New York, NY, USA, 2001; Volume 1, pp. 473–599. [Google Scholar]
- Dvorak, P.; Casamatta, D.A.; Poulickova, A.; Hasler, P.; Ondrej, V.; Sanges, R. Synechococcus: 3 billion years of global dominance. Mol. Ecol. 2014, 23, 5538–5551. [Google Scholar] [CrossRef]
- Callieri, C.; Slabakova, V.; Dzhembekova, N.; Slabakova, N.; Peneva, E.; Cabello-Yeves, P.J.; Di Cesare, A.; Eckert, E.M.; Bertoni, R.; Corno, G.; et al. The mesopelagic anoxic Black Sea as an unexpected habitat for Synechococcus challenges our understanding of global “deep red fluorescence”. ISME J. 2019, 13, 1676–1687. [Google Scholar] [CrossRef] [Green Version]
- Jardillier, L.; Zubkov, M.V.; Pearman, J.; Scanlan, D.J. Significant CO2 fixation by small prymnesiophytes in the subtropical and tropical northeast Atlantic Ocean. ISME J. 2010, 4, 1180–1192. [Google Scholar] [CrossRef]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Rincón, J.; Zabala, L.L.; Jiao, N.; Karl, D.M.; Li, W.K.W.; Lomas, M.W.; Veneziano, D.; et al. Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey-Watts, A.E.; Bindloss, M.E.; Belcher, J.H. Freshwater primary production by a blue-green alga of bacterial size. Nature 1968, 220, 1344–1345. [Google Scholar] [CrossRef] [PubMed]
- Komarek, J.; Kopecky, J.; Cepak, V. Generic characters of the simplest cyanoprokaryotes Cyanobium, Cyanobacterium and Synechococcus. Cryptogam. Algol. 1999, 20, 209–222. [Google Scholar] [CrossRef]
- Robertson, B.R.; Tezuka, N.; Watanabe, M.M. Phylogenetic analyses of Synechococcus strains (cyanobacteria) using sequences of the 16S rDNA and part of the phycocyanin operon reveal multiple evolutionary lines and reflect phycobilin content. Int. J. Syst. Evol. Microbiol. 2001, 51, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Crosbie, N.D.; Pöckl, M.; Weisse, T. Dispersal and phylogenetic diversity of nonmarine picocyanobacteria, inferred from 16S rRNA gene and cpcBA-intergenic spacer sequence analyses. Appl. Environ. Microbiol. 2003, 69, 5716–5721. [Google Scholar] [CrossRef] [Green Version]
- Callieri, C.; Coci, M.; Corno, G.; Macek, M.; Modenutti, B.; Balseiro, E.; Bertoni, R. Phylogenetic diversity of nonmarine picocyanobacteria. FEMS Microbiol. Ecol. 2013, 85, 293–301. [Google Scholar] [CrossRef]
- Scanlan, D.J.; Ostrowski, M.; Mazard, S.; Dufresne, A.; Garczarek, L.; Hess, W.R.; Post, A.F.; Hagemann, M.; Paulsen, I.; Partensky, F. Ecological genomics of marine picocyanobacteria. Microbiol. Mol. Biol. Rev. 2009, 73, 249–299. [Google Scholar] [CrossRef] [Green Version]
- Cabello-Yeves, P.J.; Haro-Moreno, J.M.; Martin-Cuadrado, A.B.; Ghai, R.; Picazo, A.; Camacho, A.; Rodriguez-Valera, F. Novel Synechococcus genomes reconstructed from freshwater reservoirs. Front. Microbiol. 2017, 8, 1151. [Google Scholar] [CrossRef] [Green Version]
- Cabello-Yeves, P.J.; Picazo, A.; Camacho, A.; Callieri, C.; Rosselli, R.; Roda-Garcia, J.J.; Coutinho, F.H.; Rodriguez-Valera, F. Ecological and genomic features of two widespread freshwater picocyanobacteria. Environ. Microbiol. 2018, 20, 3757–3771. [Google Scholar] [CrossRef]
- Doré, H.; Farrant, G.K.; Guyet, U.; Haguait, J.; Humily, F.; Ratin, M.; Pitt, F.D.; Ostrowski, M.; Six, C.; Brillet-Guéguen, L.; et al. Evolutionary mechanisms of long-term genome diversification associated with niche partitioning in marine picocyanobacteria. Front. Microbiol. 2020, 11, 2129. [Google Scholar] [CrossRef]
- Di Cesare, A.; Dzhembekova, N.; Cabello-Yeves, P.J.; Eckert, E.M.; Slabakova, V.; Slabakova, N.; Peneva, E.; Bertoni, R.; Corno, G.; Salcher, M.M.; et al. Genomic comparison and spatial distribution of different Synechococcus phylotypes in the Black Sea. Front. Microbiol. 2020, 11, 1979. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Baracaldo, P.; Bianchini, G.; Di Cesare, A.; Callieri, C.; Chrismas, N.A. Insights into the evolution of picocyanobacteria and phycoerythrin genes (mpeBA and cpeBA). Front. Microbiol. 2019, 10, 45. [Google Scholar] [CrossRef] [Green Version]
- Callieri, C.; Mandolini, E.; Bertoni, R.; Lauceri, R.; Picazo, A.; Camacho, A.; Cabello-Yeves, P.J. Atlas of picocyanobacteria monoclonal strains from the collection of CNR-IRSA, Italy. J. Limnol. 2021, 80, 2002. [Google Scholar] [CrossRef]
- Di Cesare, A.; Cabello-Yeves, P.J.; Chrismas, N.A.M.; Sánchez-Baracaldo, P.; Salcher, M.M.; Callieri, C. Genome analysis of the freshwater planktonic Vulcanococcus limneticus sp. nov. reveals horizontal transfer of nitrogenase operon and alternative pathways of nitrogen utilization. BMC Genom. 2018, 19, 259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabello-Yeves, P.J.; Scanlan, D.J.; Callieri, C.; Picazo, A.; Schallenberg, L.; Huber, P.; Roda-Garcia, J.J.; Bartosiewicz, M.; Belykh, O.I.; Tikhonova, I.V.; et al. α-cyanobacteria possessing form IA RuBisCO globally dominate aquatic habitats. ISME J. 2022. submitted. [Google Scholar]
- Hoiczyk, E.; Hansel, A. Cyanobacterial cell walls: News from an unusual prokaryotic envelope. J. Bacteriol. 2000, 182, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- Sleytr, U.B.; Messner, P.; Pum, D.; Sára, M. Crystalline bacterial cell surface layers (S-layers): From supramolecular cell structure to biomimetics and nanotechnology. Angew. Chem. Int. Ed. 1999, 38, 1034–1054. [Google Scholar] [CrossRef]
- Šmarda, J.; Šmajs, D.; Komrska, J.; Krzyzánek, V. S-layers on cell walls of cyanobacteria. Micron 2002, 33, 257–277. [Google Scholar] [CrossRef]
- Fagan, R.P.; Fairweather, N.F. Biogenesis and functions of bacterial S-layers. Nat. Rev. 2014, 12, 211. [Google Scholar] [CrossRef]
- Brahamsha, B. An abundant cell-surface polypeptide is required for swimming by the nonflagellated marine cyanobacterium Synechococcus. Proc. Natl. Acad. Sci. USA 1996, 93, 6504–6509. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, K.; Oster, G. On the Mysterious Propulsion of Synechococcus. PLoS ONE 2012, 7, e36081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarren, J.; Brahamsha, B. SwmB, a 1.12-megadalton protein that is required for nonflagellar swimming motility in Synechococcus. J. Bacteriol. 2007, 189, 1158–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCarren, J.; Brahamsha, B. Swimming motility mutants of marine Synechococcus affected in production localization of the S-layer protein SwmA. J. Bacteriol. 2009, 191, 1111–1114. [Google Scholar] [CrossRef] [Green Version]
- Giltner, C.L.; Nguyen, Y.; Burrows, L.L. Type IV pilin proteins: Versatile molecular modules. Microbiol. Mol. Biol. Rev. 2012, 76, 740–772. [Google Scholar] [CrossRef] [Green Version]
- Jezberová, J.; Komárková, J. Morphological transformation in a freshwater Cyanobium sp. induced by grazers. Environ. Microbiol. 2007, 9, 1858–1862. [Google Scholar] [CrossRef]
- Aguilo-Ferretjans, M.D.M.; Bosch, R.; Puxty, R.J.; Latva, M.; Zadjelovic, V.; Chhun, A.; Sousoni, D.; Polin, M.; Scanlan, D.; Christie-Oleza, J.A. Pili allow dominant marine cyanobacteria to avoid sinking and evade predation. Nat. Comun. 2021, 12, 1857. [Google Scholar] [CrossRef]
- Schultze-Lam, S.; Harauz, G.; Beveridge, T.J. Participation of a cyanobacterial Slayer in fine-grain mineral formation. J. Bacteriol. 1992, 174, 7971–7981. [Google Scholar] [CrossRef] [Green Version]
- De Yoreo, J.; Chung, S.; Nielsen, M.H. The dynamics and energetics of matrix assembly and mineralization. Calcif. Tissue Int. 2013, 93, 316–328. [Google Scholar] [CrossRef]
- Thompson, J.B.; Schultze-Lam, S.; Beveridge, T.J.; Des Marais, D. Whiting events: Biogenic origin due to the photosynthetic activity of cyanobacterial picoplankton. Limnol. Oceanogr. 1998, 42, 133–141. [Google Scholar]
- Dittrich, M.; Müller, B.; Mavrocordatos, D.; Wehrli, B. Induced calcite precipitation by cyanobacterium Synechococcus. Acta Hydroch. Hydrob. 2003, 31, 162–169. [Google Scholar] [CrossRef]
- Schultze-Lam, S.; Beveridge, T.J. Nucleation of celestite and strontianite on a cyanobacterial S-layer. Appl. Environ. Microbiol. 1994, 60, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Shirokova, L.S.; Mavromatis, V.; Bundelava, I.A.; Pokrovsky, O.S.; Bénézeth, P.; Gérard, E.; Pearce, C.R.; Oelkers, E.H. Using Mg isotopes to trace cyanobacterially mediated magnesium carbonate precipitation in alkaline lakes. Aquat. Geochem. 2013, 19, 1–24. [Google Scholar] [CrossRef]
- Lima, S.; Matinha-Cardoso, J.; Tamagnini, P.; Oliveira, P. Extracellular vescicles: An overlooked secretion system in cyanobacteria. Life 2020, 10, 129. [Google Scholar] [CrossRef] [PubMed]
- Farrant, G.K.; Doré, H.; Cornejo-Castillo, F.M.; Partensky, F.; Ratin, M.; Ostrowski, M.; Pitt, F.D.; Wincher, P.; Scanlan, D.J.; Iudicone, D.; et al. Delineating ecologically significant taxonomic units from global patterns of marine picocyanobacteria. Proc. Natl. Acad. Sci. USA 2016, 113, E3365–E3374. [Google Scholar] [CrossRef] [Green Version]
- Six, C.; Thomas, J.-C.; Garczarek, L.; Ostrowski, M.; Dufresne, A.; Blot, N.; Scanlan, D.J.; Partensky, F. Diversity and evolution of phycobilisomes in marine Synechococcus spp.: A comparative genomics study. Genome Biol. 2007, 8, R259. [Google Scholar] [CrossRef] [PubMed]
- Grébert, T.; Doré, H.; Partensky, F.; Farrant, G.K.; Boss, E.S.; Picheral, M.; Guidi, L.; Pesant, S.; Scanlan, D.J.; Winker, P.; et al. Light color acclimation is a key process in the global ocean distribution of Synechococcus cyanobacteria. Proc. Natl. Acad. Sci. USA 2018, 115, E2010–E2019. [Google Scholar] [CrossRef] [Green Version]
- Palenik, B.; Brahamsha, B.; Larimer, F.; Land, M.; Hauser, L.; Chain, P.; Lamerdin, J.; Regala, W.; Allen, E.E.; McCarren, J.; et al. The genome of a motile marine Synechococcus. Nature 2003, 424, 1037–1042. [Google Scholar] [CrossRef] [Green Version]
- Wood, A.M.; Horan, P.K.; Muirhead, K.; Phinney, D.A.; Yentsch, C.M.; Waterbury, J.B. Discrimination between types of pigments in marine Synechococcus spp. by scanning spectroscopy, epifluorescence microscopy and flow cytometry. Limnol. Oceanogr. 1985, 30, 1303–1315. [Google Scholar] [CrossRef]
- Vörös, L.; Callieri, C.; Balogh, K.V.; Bertoni, R. Freshwater picocyanobacteria along a trophic gradient and light quality range. Hydrobiologia 1998, 369/370, 117–125. [Google Scholar] [CrossRef]
- Callieri, C. Micro-players for macro-roles: Aquatic microbes in deep lakes. J. Limnol. 2016, 75, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Stomp, M.; Huisman, J.; Vörös, L.; Pick, F.R.; Laamanen, M.; Haverkamp, T.; Stal, L.J. Colorful coexistence of red and green picocyanobacteria in lakes and seas. Ecol. Lett. 2007, 10, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Fuller, N.J.; Marie, D.; Partensky, F.; Vaulot, D.; Post, A.F.; Scanlan, D.J. Clade-specific 16S ribosomal DNA oligonucleotides reveal the predominance of a single marine Synechococcus clade throughout a stratified water column in the Red Sea. Appl. Environ. Microb. 2003, 69, 2430–2443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Śliwińska-Wilczewska, S.; Konarzewska, Z.; Wiśniewska, K.; Konik, M. Photosynthetic pigments changes of three phenotypes of picocyanobacteria Synechococcus sp. under different light and temperature conditions. Cells 2020, 9, 2030. [Google Scholar] [CrossRef]
- Everroad, C.; Six, C.; Partensky, F.; Thomas, J.C.; Holtzendorff, J.; Wood, A.M. Biochemical bases of type IV chromatic adaptation in marine Synechococcus spp. J. Bacteriol. 2006, 188, 3345–3356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luimstra, V.M.; Schuurmans, J.M.; de Carvalho, C.F.M.; Matthijs, H.C.P.; Hellingwerf, K.J.; Huisman, J. Exploring the low photosynthetic efficiency of cyanobacteria in blue light using a mutant lacking phycobilisomes. Photosynth. Res. 2019, 141, 291–301. [Google Scholar] [CrossRef]
- Scanlan, D.J.; West, N.J. Molecular ecology of the marine cyanobacterial genera Prochlorococcus and Synechococcus. FEMS Microbiol. Ecol. 2002, 40, 1–12. [Google Scholar] [CrossRef]
- Lemasson, C.; Tandeau de Marsac, N.; Cohen-Bazire, G. Role of allophycocyanin as light-harvesting pigment in cyanobacteria. Proc. Natl. Acad. Sci. USA 1973, 70, 3130–3133. [Google Scholar] [CrossRef] [Green Version]
- Bruce, D.; Brimble, S.; Bryant, D.A. State transitions in a phycobilisome-less mutant of the cyanobacterium Synechococcus sp. PCC 7002. Biochim. Biophys. Acta Bioenerg. 1989, 974, 66–73. [Google Scholar] [CrossRef]
- Mullineaux, C.W. Phycobilisome-reaction centre interaction in cyanobacteria. Photosynth. Res. 2008, 95, 175–182. [Google Scholar] [CrossRef]
- Li, W.K.W. Annual average abundance of heterotrophic bacteria and Synechococcus in surface ocean waters. Limnol. Oceanogr. 1998, 43, 1746–1753. [Google Scholar] [CrossRef]
- Callieri, C.; Piscia, R. Photosynthetic efficiency and seasonality of autotrophic picoplankton in Lago Maggiore after its recovery. Freshw. Biol. 2002, 47, 941–956. [Google Scholar] [CrossRef]
- Weisse, T. Dynamics of autotrophic picoplankton in marine and freshwater ecosystems. Adv. Microb. Ecol. 1993, 13, 327–370. [Google Scholar]
- Callieri, C. Picophytoplankton in freshwater ecosystems: The importance of small-sized phototrophs. Freshw. Rev. 2008, 1, 1–28. [Google Scholar] [CrossRef]
- Camacho, A.; Miracle, M.R.; Vicente, E. Which factors determine the abundance and distribution of picocyanobacteria in inland waters? A comparison among different types of lakes and ponds. Arch. Hydrobiol. 2003, 157, 321–338. [Google Scholar] [CrossRef]
- Callieri, C.; Modenutti, B.; Queimaliños, C.; Bertoni, R.; Balseiro, E. Production and biomass of picophytoplankton and larger autotrophs in Andean ultraoligotrophic lakes: Differences in light harvesting efficiency in deep layers. Aquat. Ecol. 2007, 41, 511–523. [Google Scholar] [CrossRef]
- Belykh, O.I.; Ekaterina, G.; Sorokovikova, T.; Saphonova, A.; Tikhonova, I.V. Autotrophic picoplankton of Lake Baikal: Composition, abundance and structure. Hydrobiologia 2006, 568, 9–17. [Google Scholar] [CrossRef]
- Marchant, H.J.; Davidson, A.T.; Wright, S.W. The distribution and abundance of chroococcoid cyanobacteria in the Southern Ocean. Proc. NIPR Symp. Polar. Biol. 1987, 1, 1–9. [Google Scholar]
- Murphy, L.S.; Haugen, E.M. The distribution and abundance of phototrophic ultraplankton in the North Atlantic. Limnol. Oceanogr. 1985, 30, 47–58. [Google Scholar] [CrossRef]
- Waleron, M.; Waleron, K.; Vincent, W.F.; Wilmott, A. Allochthonous inputs of riverine picocyanobacteria to coastal waters in the Arctic Ocean. FEMS Microbiol. Ecol. 2007, 59, 356–365. [Google Scholar] [CrossRef]
- Cottrell, M.T.; Kirchman, D.L. Photoheterotrophic microbes in the Arctic Ocean in summer and winter. Appl. Environ. Microbiol. 2009, 75, 4958–4966. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, M.L.; Doré, H.; Garczarek, L.; Seuthe, L.; Müller, O.; Sandaa, R.-A.; Bratbak, G.; Larsen, A. Synechococcus in the Atlantic Gateway to the Arctic Ocean. Front. Mar. Sci. 2016, 3, 191. [Google Scholar] [CrossRef] [Green Version]
- Cabello-Yeves, P.J.; Callieri, C.; Picazo, A.; Mehrshad, M.; Haro-Moreno, J.M.; Roda-Garcia, J.J.; Dzhembekova, N.; Slabakova, V.; Slabakova, N.; Moncheva, S.; et al. Microbiome of the Black Sea water column analyzed by genome centric metagenomics. Environ. Microbiome 2021, 16, 5. [Google Scholar] [CrossRef]
- Andersson, A.; Haecky, P.; Hagström, A. Effect of temperature and light on the growth of micro- nano- and pico-plankton: Impact on algal succession. Mar. Biol. 1994, 120, 511–520. [Google Scholar] [CrossRef]
- Izaguirre, I.; Unrein, F.; Modenutti, B.; Allende, L. Photosynthetic picoplankton in Argentina lakes Advanc. Limnol. 2014, 65, 343–357. [Google Scholar]
- Felföldi, T.; Duleba, M.; Somogyi, B.; Vajna, B.; Nikolausz, M.; Présing, M.; Márialigeti, K.; Vörös, L. Diversity and seasonal dynamics of the photoautotrophic picoplankton in Lake Balaton (Hungary). Aquat. Microb. Ecol. 2011, 63, 273–287. [Google Scholar] [CrossRef] [Green Version]
- Schallenberg, L.; Pearman, J.K.; Burns, C.W.; Wood, S. Metabarcoding reveals lacustrine picocyanobacteria respond to environmental change through adaptive community structuring. Front. Microbiol. 2021, 12, 757929. [Google Scholar] [CrossRef]
- Pestova, D.; Macek, M.; Martinez Perez, M.E. Ciliates and their picophytoplankton-feeding activity in a high altitude warm-monomictic saline lake. Europ. J. Protist. 2008, 44, 13–25. [Google Scholar] [CrossRef]
- Picazo Mozo, A. Ecología del Picoplancton en Una Laguna Cárstica Estratificada. Ph.D. Thesis, Universidad de Valencia, Valencia, Spain, 2015; p. 246. [Google Scholar]
- Mills, M.M.; Ridame, C.; Davey, M.; La Roche, J. Iron and phosphorus co-limit nitrogen fixation in the eastern tropical North Atlantic. Nature 2004, 429, 292–294. [Google Scholar] [CrossRef]
- Schindler, D.W. Recent advantages in understanding and management of eutrophication. Limnol. Oceanogr. 2006, 51, 351–355. [Google Scholar] [CrossRef] [Green Version]
- Diaz, M.; Pedrozo, F.; Reynolds, C.; Temporetti, P. Chemical composition and the nitrogen-regulated trophic state of Patagonian lakes. Limnologica 2007, 37, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Moore, L.R.; Ostrowsky, M.; Scanlan, D.J.; Feren, K.; Sweetsir, T. Ecotypic variation in phosphorus-acquisition mechanisms within marine picocyanobacteria. Aquat. Microb. Ecol. 2005, 39, 257–269. [Google Scholar] [CrossRef] [Green Version]
- Mastala, Z.; Herodek, S.; Balogh, K.V.; Borbély, G.; Shafik, H.M.; Vörös, L. Nutrient requirement and growth of a Synechococcus species isolated from Lake Balaton. Int. Rev. Gesamten Hydrobiol. 1996, 81, 503–512. [Google Scholar] [CrossRef]
- Glibert, P.M.; Ray, R.T. Different patterns of growth and nitrogen uptake in two clones of marine Synechococcus spp. Mar. Biol. 1990, 107, 273–280. [Google Scholar] [CrossRef]
- Bird, C.; Wyman, M. Nitrate/nitrite assimilation system of the marine picoplanktonic cyanobacterium Synechococcus sp. strain WH8103: Effect of nitrogen source and availability on gene expression. Appl. Environ. Microbiol. 2003, 69, 7009–7018. [Google Scholar] [CrossRef] [Green Version]
- Saito, M.A.; McIlvin, M.R.; Moran, D.M.; Goepfert, T.J.; DiTullio, G.R.; Post, A.F.; Lamborg, C.H. Multiple nutrient stresses at intersecting Pacific Ocean biomes detected by protein biomarkers. Science 2014, 345, 1173–1177. [Google Scholar] [CrossRef]
- Wawrik, B.; Callaghan, A.V.; Bronk, D.A. Use of inorganic and organic nitrogen by Synechococcus spp. and diatoms on the west Florida shelf as measured using stable isotope probing. Appl. Environ. Microbiol. 2009, 75, 6662–6670. [Google Scholar] [CrossRef] [Green Version]
- Simon, R.D. Inclusion bodies in the cyanobacteria: Cyanophycin, polyphospate, polyhedral bodies. In The Cyanobacteria; Fay, P., Van Baalen, C., Eds.; Elsevier: Amsterdam, The Netherlands, 1987; pp. 199–225. [Google Scholar]
- Donald, K.M.; Scanlan, D.J.; Carr, N.G.; Mann, N.H.; Joint, I. Comparative phosphorus nutrition of the marine cyanobacterium Synechococcus WH7803 and the marine diatom Thalassiosira weissflogii. J. Plankton Res. 1997, 19, 1793–1813. [Google Scholar] [CrossRef] [Green Version]
- Stockner, J.G. Autotrophic picoplankton in freshwater ecosystems: The view from summit. Int. Rev. Gesamten Hydrobiol. 1991, 76, 483–492. [Google Scholar] [CrossRef]
- Agawin, N.S.R.; Duarte, C.M.; Agusti, S. Nutrient and temperature control of the contribution of picoplankton to phytoplankton biomass and production. Limnol. Oceanogr. 2000, 45, 591–600. [Google Scholar] [CrossRef]
- Bell, T.; Kalff, J. The contribution of picophytoplankton in marine and freshwater system of different trophic status and depth. Limnol. Oceanogr. 2001, 46, 1243–1248. [Google Scholar] [CrossRef]
- Callieri, C.; Stockner, J.G. Freshwater autotrophic picoplankton: A review. J. Limnol. 2002, 6, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Somogyi, B.; Felföldi, T.; Tóth, L.G.; Bernat, G.; Vörös, L. Photoautotrophic picoplankton- a review on their occurrence role and diversity in Lake Balaton. Biol. Futur. 2020, 71, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Christie-Oleza, J.A.; Armengaud, J.; Guerin, P.; Scanlan, D.J. Functional distinctness in the exoproteomes of marine Synechococcus. Environ. Microbiol. 2015, 17, 3781–3794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, R. Carbon-14 uptake by picoplankton and total phytoplankton in eight New Zealand lakes. Int. Rev. Gesamten Hydrobiol. 1991, 76, 631–641. [Google Scholar] [CrossRef]
- Nagata, T.; Takai, K.; Kawanobe, K.; Kim, D.; Nakazato, R.; Guselnikova, N.; Bondarenko, N.; Mologawaya, O.; Kostrnova, T.; Drucker, V.; et al. Autotrophic picoplankton in southern Lake Baikal: Abundance growth and grazing mortality during summer. J. Plankton Res. 1994, 16, 945–959. [Google Scholar] [CrossRef]
- Marañón, E.; Holligan, P.M.; Barciela, R.; Gonzalez, N.; Mourino, B.; Pazo, M.J.; Varela, M. Patterns of phytoplankton size structure and productivity in contrasting open-ocean environments. Mar. Ecol. Prog. Ser. 2001, 216, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Ostos, E.; Fernández, A.; Huete-Ortega, M.; Mouriño-Carballido, B.; Calvo-Díaz, A.; Morán, X.A.G.; Marañón, E. Size-fractionated phytoplankton biomass and production in the tropical Atlantic. Sci. Mar. 2011, 75, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Li, W.K.W. Primary productivity of prochlorophytes, cyanobacteria, and eucaryotic ultraphytoplankton: Measurements from flow cytometric sorting. Limnol. Oceanogr. 1994, 39, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Stockner, J.; Callieri, C.; Cronberg, G. Picoplankton and other nonbloom forming cyanobacteria in lakes. In The Ecology of Cyanobacteria: Their Diversity in Time and Space; Whitton, B.A., Potts, M., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 195–238. [Google Scholar]
- Ong, L.J.; Glazer, A.N.; Waterbury, J.B. An unusual phycoerythrin from a marine cyanobacterium. Science 1984, 224, 80–83. [Google Scholar] [CrossRef]
- Callieri, C.; Pinolini, M.L. Picoplankton in Lake Maggiore, Italy. Int. Rev. Gesamten Hydrobiol. 1995, 80, 491–501. [Google Scholar] [CrossRef]
- Six, C.; Thomas, J.C.; Thion, L.; Lemoine, Y.; Zal, F.; Partensky, F. Two novel phycoerythrin-associated linker proteins in the marine cyanobacterium Synechococcus sp. strain WH8102. J. Bacteriol. 2005, 87, 1685–1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahlgren, N.; Rocap, G. Culture isolation and culture independent clone libraries reveal new marine Synechococcus ecotypes with distinctive light and N physiologies. Appl. Environ. Microbiol. 2006, 72, 7193–7204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Song, Q.; Zheng, P.; Zhang, X.; Zou, S.; Li, Y.; Gao, X.; Zhao, Z.; Gong, J. Dynamics and distribution of marine Synechococcus abundance and genotypes during seasonal hypoxia in a coastal marine ranch. J. Mar. Sci. Eng. 2021, 9, 549. [Google Scholar] [CrossRef]
- Liu, H.; Jing, H.; Wong, T.H.C.; Chen, B. Co-occurrence of phycocyanin- and phycoerythrin-rich Synechococcus in subtropical estuarine and coastal waters of Hong Kong. Environ. Microbiol. Rep. 2014, 6, 90–99. [Google Scholar] [CrossRef] [Green Version]
- Jasser, I.; Krolicka, A.; Karnkowska-Ishikawa, A. A novel phylogenetic clade of picocyanobacteria from the Mazurian lakes (Poland) reflects the early ontogeny of glacial lakes. FEMS Microbiol. Ecol. 2011, 75, 89–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, V.; Palenik, B. Temporal variation of Synechococcus clades at a coastal Pacific Ocean monitoring site. ISME J. 2009, 3, 903–915. [Google Scholar] [CrossRef] [Green Version]
- Six, C.M.; Finkel, Z.V.; Irwin, A.J.; Campbell, D.A. Light variability illuminates niche-partitioning among marine picocyanobacteria. PLoS ONE 2007, 2, e1341. [Google Scholar] [CrossRef] [Green Version]
- Mazard, S.; Ostrowski, M.; Partensky, F.; Scanlan, D.J. Multi-locus sequence analysis, taxonomic resolution and biogeography of marine Synechococcus. Environ. Microbiol. 2012, 14, 372–386. [Google Scholar] [CrossRef]
- Pittera, J.; Humily, F.; Thorel, M.; Grulois, D.; Garczarek, L.; Six, C. Connecting thermal physiology and latitudinal niche partitioning in marine Synechococcus. ISME J. 2014, 8, 1221–1236. [Google Scholar] [CrossRef]
- Kim, Y.; Jeon, J.; Kwak, M.S.; Kim, G.H.; Koh, I.; Rho, M. Photosynthetic functions of Synechococcus in the ocean microbiomes of diverse salinity and seasons. PLoS ONE 2018, 13, e0190266. [Google Scholar] [CrossRef] [Green Version]
- Zwirglmaier, K.; Heywood, J.L.; Chamberlain, K.; Woodward, E.M.S.; Zubkov, M.V.; Scanlan, D.J. Basin-scale distribution patterns of picocyanobacterial lineages in the Atlantic Ocean. Environ. Microbiol. 2007, 9, 1278–1290. [Google Scholar] [CrossRef] [PubMed]
- Fuller, N.J.; Tarran, G.A.; Yallop, M.; Orcutt, K.M.; Scanlan, D.J. Molecular analysis of picocyanobacterial community structure along an Arabian Sea transect reveals distinct spatial separation of lineages. Limnol. Oceanogr. 2006, 51, 2515–2526. [Google Scholar] [CrossRef]
- Callieri, C.; Stockner, J. Picocyanobacteria success in oligotrophic lakes: Fact or fiction? J. Limnol. 2000, 59, 72–76. [Google Scholar] [CrossRef]
- Hawley, G.R.W.; Whitton, B.A. Seasonal changes in chlorophyll-containing picoplankton populations in ten lakes in northern England. Int. Rev. Gesamten Hydrobiol. 1991, 76, 545–554. [Google Scholar] [CrossRef]
- Sánchez-Baracaldo, P.; Handley, B.A.; Hayes, P.K. Picocyanobacterial community structure of freshwater lakes and the Baltic Sea revealed by phylogenetic analyses and clade specific quantitative PCR. Microbiology 2008, 154, 3347–3357. [Google Scholar] [CrossRef] [Green Version]
- Becker, S.; Rich, P.; Ernst, A. Seasonal and habitat-related distribution pattern of Synechococcus genotypes in Lake Constance. FEMS Microbiol. Ecol. 2007, 62, 64–77. [Google Scholar] [CrossRef]
- Ivanikova, N.V.; Popels, L.C.; McKay, M.L.; Bullerjahn, G.S. Lake Superior supports novel clusters of cyanobacterial picoplankton. Appl. Environ. Microbiol. 2007, 73, 4055–4065. [Google Scholar] [CrossRef] [Green Version]
- Callieri, C.; Caravati, E.; Corno, G.; Bertoni, R. Picocyanobacterial community structure and space-time dynamics in the subalpine Lake Maggiore (N. Italy). J. Limnol. 2012, 71, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Walker, T.D.; Marchant, H.J. The seasonal occurrence of chroococcoid cyanobacteria at an Antarctic Coastal Site. Polar Biol. 1989, 9, 193–196. [Google Scholar] [CrossRef]
- Sohrin, R.; Isaji, M.; Obara, Y.; Agostini, S.; Suzuki, Y.; Hiroe, Y.; Ichikawa, T.; Hidaka, K. Distribution of Synechococcus in the dark ocean. Aquat. Microb. Ecol. 2011, 64, 1–14. [Google Scholar] [CrossRef]
- Béjà, O.; Spudich, E.N.; Spudich, J.L.; Leclerc, M.; DeLong, E.F. Proteorhodopsin phototrophy in the ocean. Nature 2001, 411, 786–789. [Google Scholar] [CrossRef] [PubMed]
- Eiler, A. Evidence for the ubiquity of mixotrophic bacteria in the upper ocean: Implications and consequences. Appl. Environ. Microbiol. 2006, 7, 7431–7437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottrell, M.T.; Michelou, V.K.; Nemcek, N.; DiTullio, G.; Kirchman, D.L. Carbon cycling by microbes influenced by light in the northeast Atlantic Ocean. Aquat. Microb. Ecol. 2008, 50, 239–250. [Google Scholar] [CrossRef]
- Yelton, A.P.; Acinas, S.G.; Sunagawa, S.; Bork, P.; Pedrós-Alió, C.; Chisholm, S.W. Global genetic capacity for mixotrophy in marine picocyanobacteria. ISME J. 2016, 10, 2946–2957. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Marín, M.C.; Gómez-Baena, G.; López-Lozano, A.; Moreno-Cabezuelo, J.A.; Díez, J.; García-Fernández, J.M. Mixotrophy in marine picocyanobacteria: Use of organic compounds by Prochlorococcus and Synechococcus. ISME J. 2020, 14, 1065–1073. [Google Scholar] [CrossRef]
- Konovalov, S.K.; Murray, J.W.; Luther, G.W., III. Basic processes of Black Sea Biogeochemistry. Oceanography 2005, 18, 24–35. [Google Scholar] [CrossRef]
- Xing, X.; Claustre, H.; Boss, E.; Roesler, C.; Organelli, E.; Poteau, A.; Barbieux, M.; D’Ortenzio, F. Correction of profiles of in-situ chlorophyll fluorometry for the contribution of fluorescence originating from non-algal matter. Limnol. Oceanogr. Methods 2017, 15, 80–93. [Google Scholar] [CrossRef]
- Broenkow, W.W.; Yuen, M.A.; Yarbrough, M.A. VERTEX: Biological implications of total attenuation and chlorophyll and phycoerythrin fluorescence distributions along a 2000 m deep section in the Gulf of Alaska. Deep Sea Res. Part A Ocean. Res. Pap. 1992, 39, 417–437. [Google Scholar] [CrossRef]
- Pascoal, F.; Costa, R.; Magalhaes, C. The microbial rare biosphere: Current concepts, methods and ecological principles. FEMS Microb. Ecol. 2021, 97, fiaa227. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Callieri, C.; Cabello-Yeves, P.J.; Bertoni, F. The “Dark Side” of Picocyanobacteria: Life as We Do Not Know It (Yet). Microorganisms 2022, 10, 546. https://doi.org/10.3390/microorganisms10030546
Callieri C, Cabello-Yeves PJ, Bertoni F. The “Dark Side” of Picocyanobacteria: Life as We Do Not Know It (Yet). Microorganisms. 2022; 10(3):546. https://doi.org/10.3390/microorganisms10030546
Chicago/Turabian StyleCallieri, Cristiana, Pedro J. Cabello-Yeves, and Filippo Bertoni. 2022. "The “Dark Side” of Picocyanobacteria: Life as We Do Not Know It (Yet)" Microorganisms 10, no. 3: 546. https://doi.org/10.3390/microorganisms10030546
APA StyleCallieri, C., Cabello-Yeves, P. J., & Bertoni, F. (2022). The “Dark Side” of Picocyanobacteria: Life as We Do Not Know It (Yet). Microorganisms, 10(3), 546. https://doi.org/10.3390/microorganisms10030546