
Fe(II) Addition Drives Soil Bacterial Co-Ocurrence Patterns and Functions Mediated by Anaerobic and Chemoautotrophic Taxa


Abstract
:
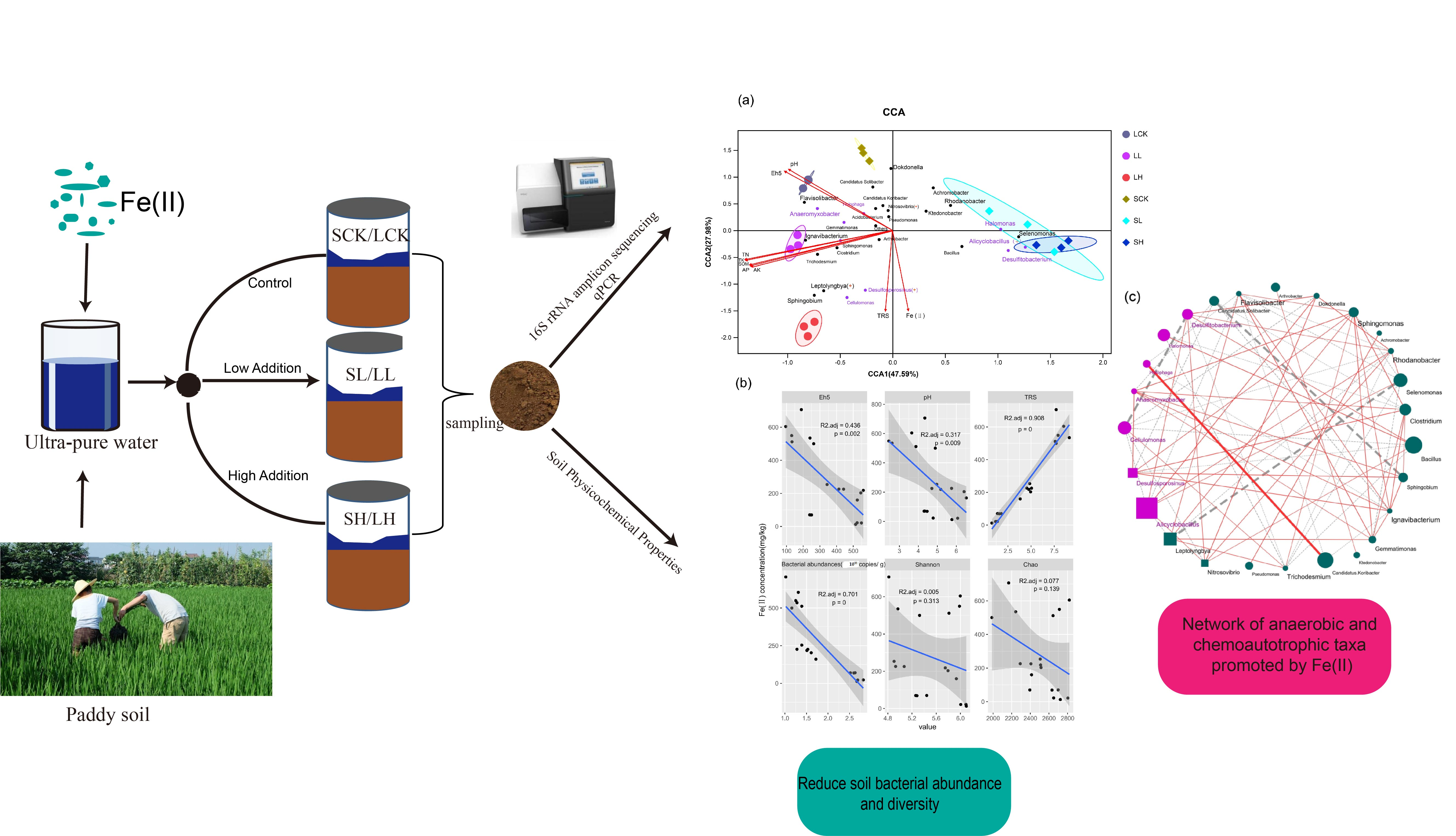{kind=link}
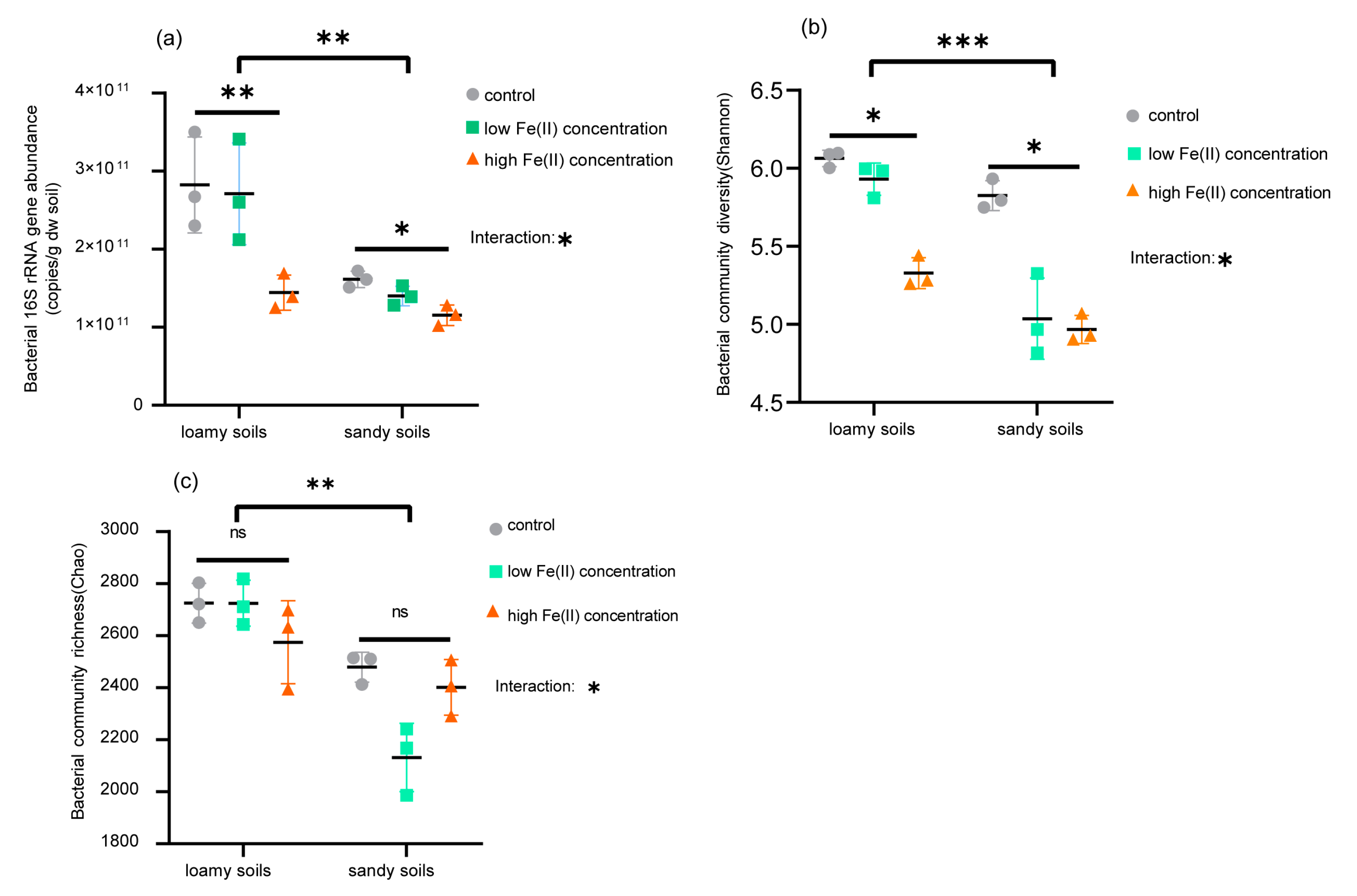{kind=link}
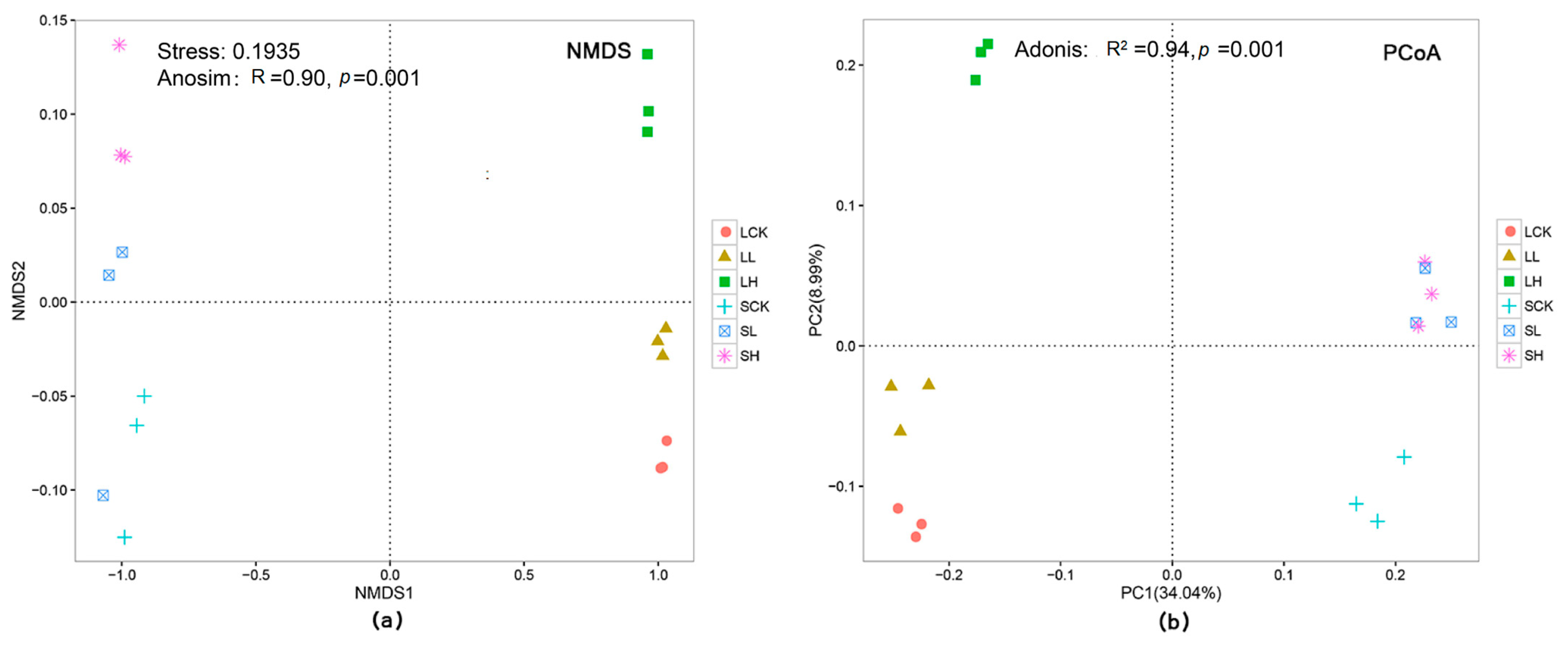{kind=link}
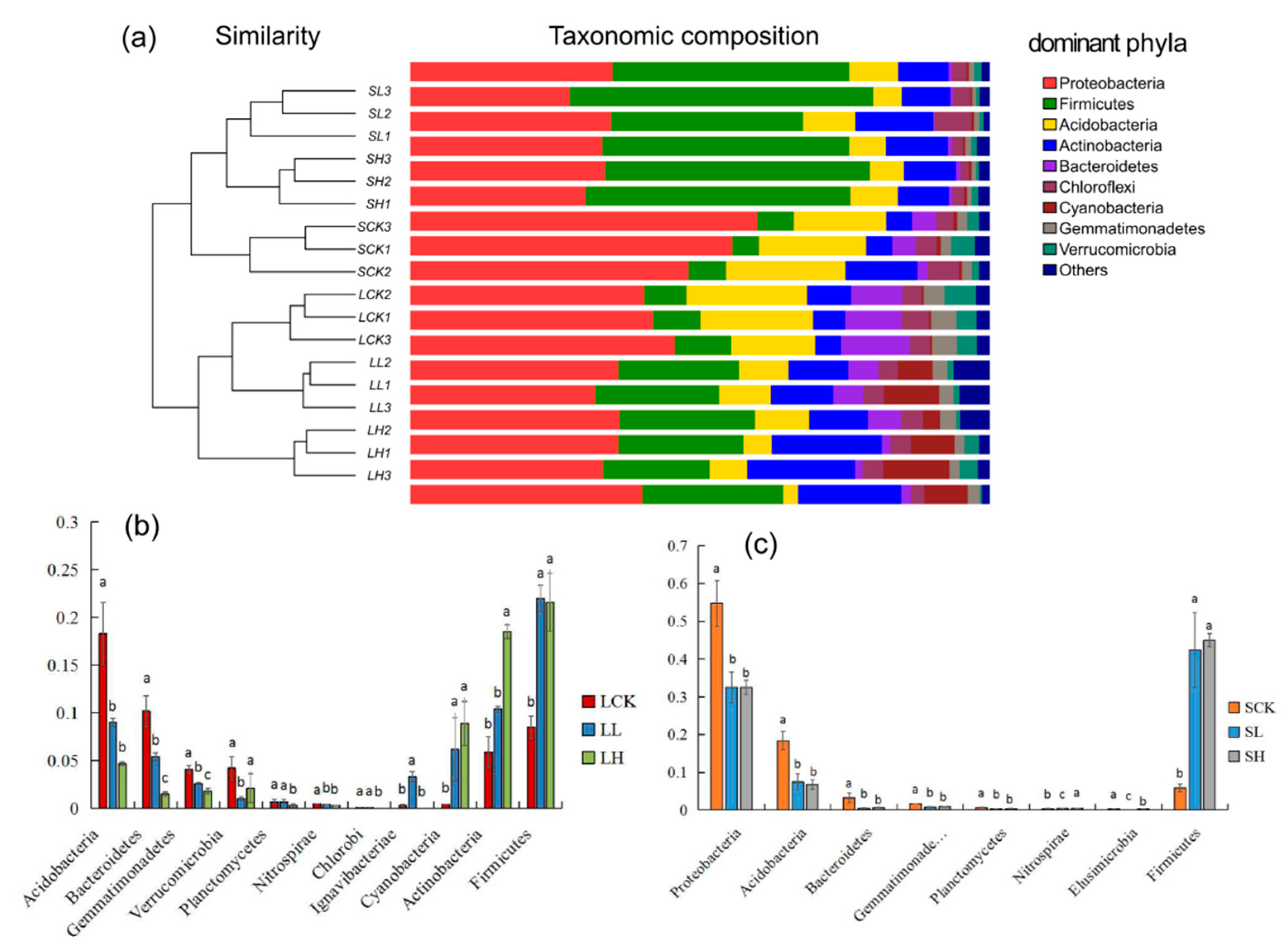{kind=link}
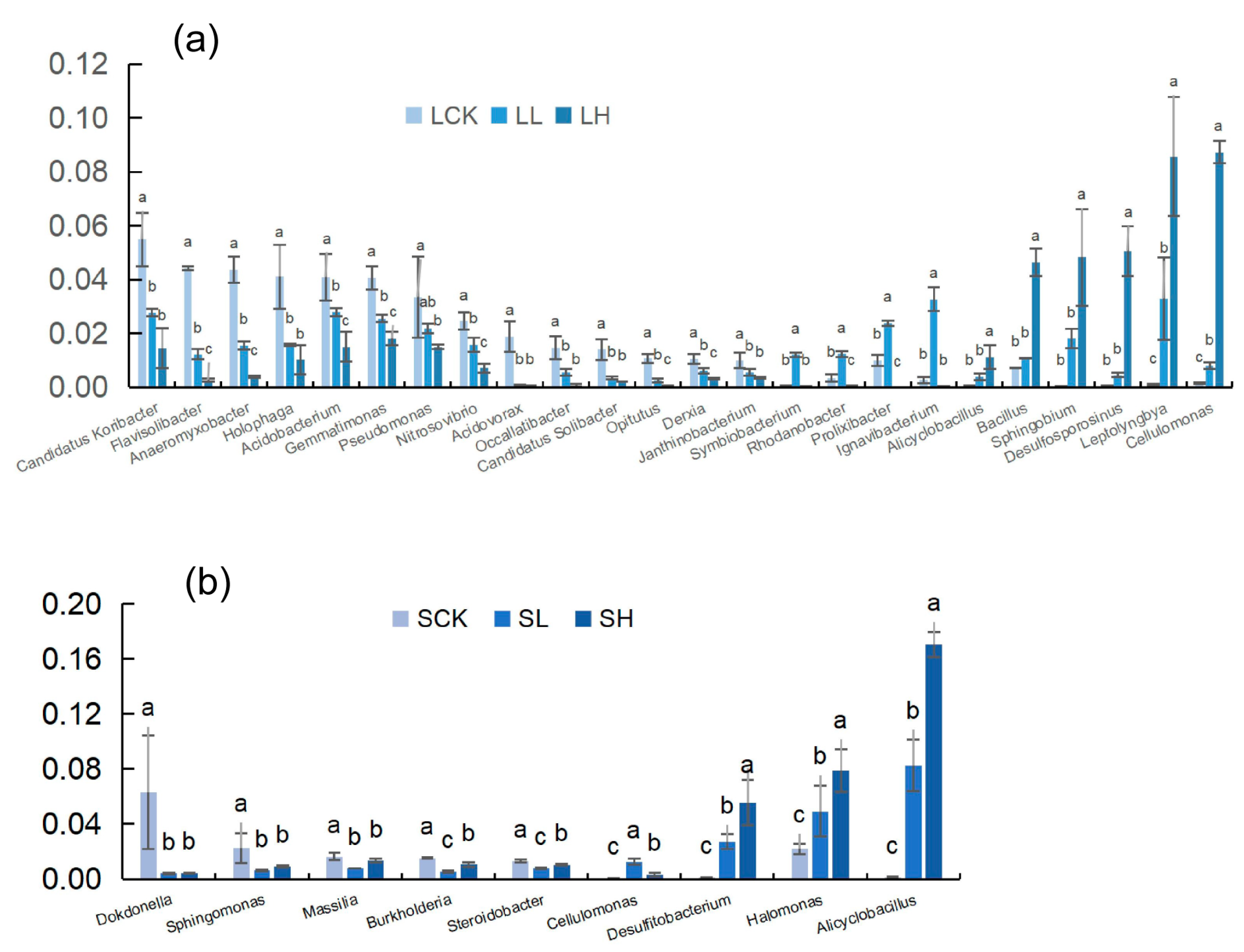{kind=link}
{kind=link}
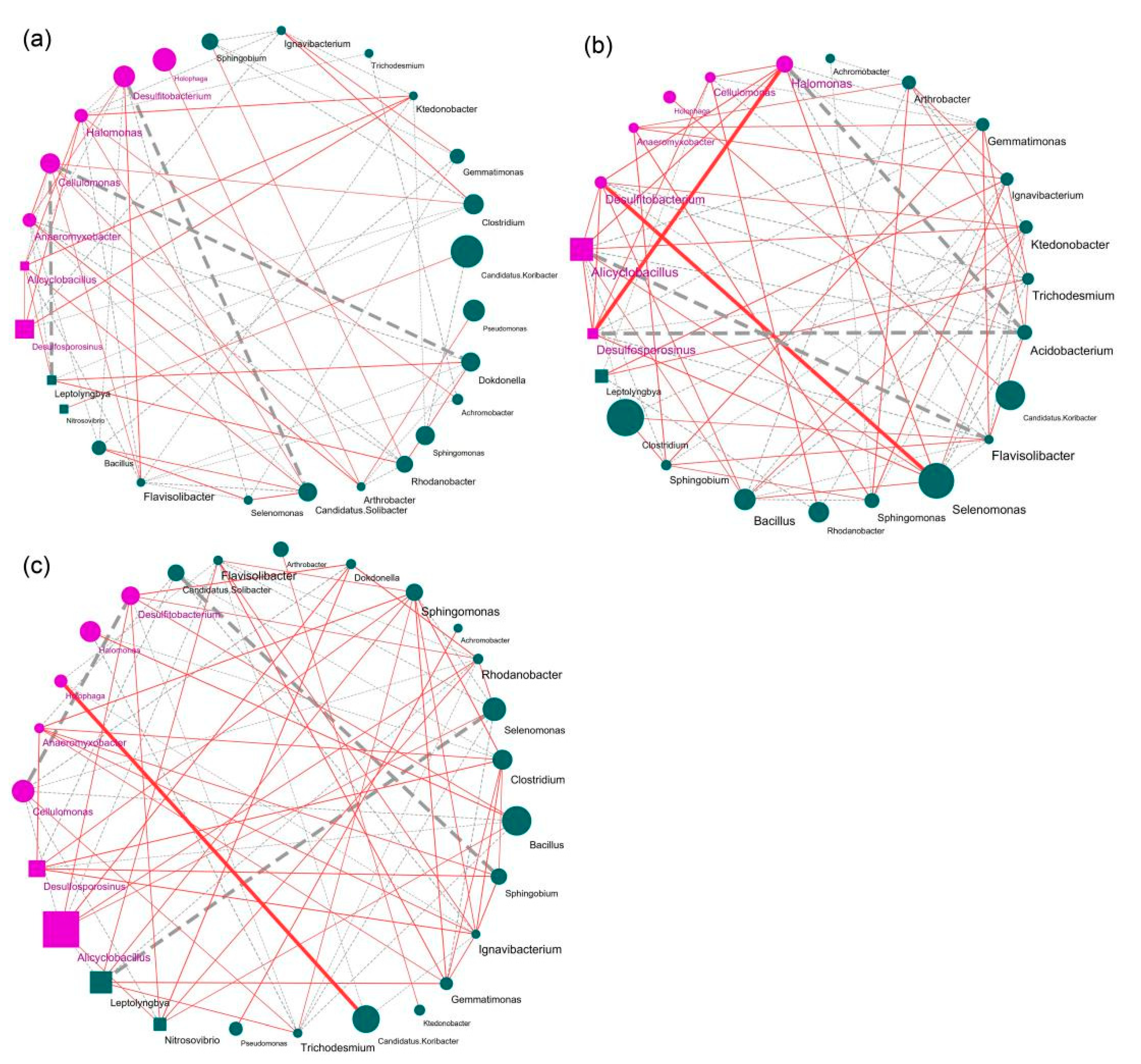{kind=link}
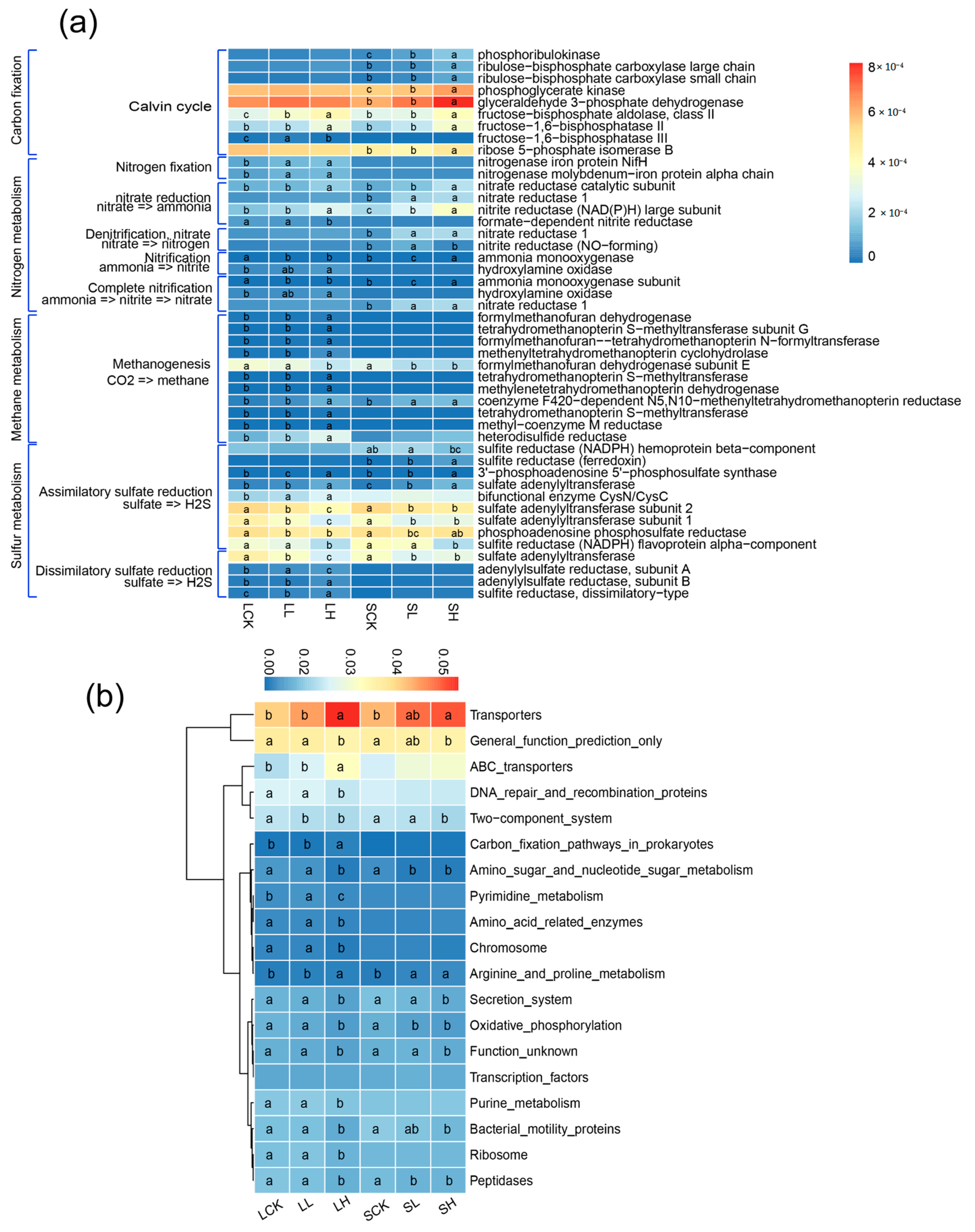{kind=link}
1. Introduction
2. Materials and Methods
2.1. Site Description and Soil Sampling
2.2. Experimental Design and Soil Hysicochemical Properties
2.3. Total Microbial DNA Extraction, PCR Amplification and Illumina Sequencing
2.4. Soil Bacterial Abundance Analysis
2.5. Microbiome Bioinformatics and Statistical Analysis
2.6. Co-Occurrence Network Analysis
3. Results
3.1. Soil Bacterial Abundance and Community Diversity
3.2. Analysis of Soil Bacterial Community Structure under Different Fe(II) Concentrations
3.3. Relationships between Bacterial Communities and Soil Physicochemical Properties
3.4. Potential Bacterial Co-Occurrence Networks of Fe(II)-Mediated Interactions
3.5. Prediction of Bacterial Metabolic Functions Based on PICRUSt
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tian, Z.; Niu, Y.; Fan, D.; Sun, L.; Ficsher, G.; Zhong, H.; Deng, J.; Tubiello, F.N. Maintaining rice production while mitigating methane and nitrous oxide emissions from paddy fields in China: Evaluating tradeoffs by using coupled agricultural systems models. Agric. Syst. 2018, 159, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Lv, Z.; Zhu, Y.; Liu, X.; Ye, H.; Tian, Y.; Li, F. Climate change impacts on regional rice production in China. Clim. Chang. 2018, 147, 523–537. [Google Scholar] [CrossRef]
- Sun, M.; Xiao, T.; Ning, Z.; Xiao, E.; Sun, W. Microbial community analysis in rice paddy soils irrigated by acid mine drainage contaminated water. Appl. Microbiol. Biotechnol. 2014, 99, 2911–2922. [Google Scholar] [CrossRef]
- Pezeshki, S.R.; Delaune, R.D. Soil Oxidation-Reduction in Wetlands and Its Impact on Plant Functioning. Biology 2012, 1, 196–221. [Google Scholar] [CrossRef] [Green Version]
- Bowles, J.F.W. The Iron Oxides: Structure, Properties Reactions Occurrence and Uses (Review). Mineral. Mag. 1997, 61, 740–741. [Google Scholar] [CrossRef]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen–host interface. Nat. Rev. Genet. 2012, 10, 525–537. [Google Scholar] [CrossRef]
- Schmidt, C.; Behrens, S.; Kappler, A. Ecosystem functioning from a geomicrobiological perspective—A conceptual framework for biogeochemical iron cycling. Environ. Chem. 2010, 7, 399–405. [Google Scholar] [CrossRef] [Green Version]
- Weber, K.; Achenbach, L.A.; Coates, J.D. Microorganisms pumping iron: Anaerobic microbial iron oxidation and reduction. Nat. Rev. Genet. 2006, 4, 752–764. [Google Scholar] [CrossRef] [Green Version]
- Straub, K.L.; Benz, M.; Schink, B.; Widdel, F. Anaerobic, nitrate-dependent microbial oxidation of ferrous iron. Appl. Environ. Microbiol. 1996, 62, 1458–1460. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.X.; Haudenshield, J.S.; Bowen, C.R.; Hartman, G.L. Metagenome-wide association study and machine learning prediction of bulk soil microbiome and crop productivity. Front. Microbiol. 2017, 8, 519. [Google Scholar] [CrossRef]
- Liesack, W.; Schnell, S.; Revsbech, N.P. Microbiology of flooded rice paddies. FEMS Microbiol. Rev. 2000, 24, 625–645. [Google Scholar] [CrossRef] [PubMed]
- Hegler, F.; Posth, N.R.; Jiang, J.; Kappler, A. Physiology of phototrophic iron (II)-oxidizing bacteria: Implications for modern and ancient environments. FEMS Microbiol. Ecol. 2008, 66, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo, D.M.; Pollard, A.G. Chemistry of waterlogged soils. I.—Changes in oxidation-reduction potentials of two soils on submergence; Influence of pH and organic matter. J. Sci. Food Agric. 1962, 13, 43–48. [Google Scholar] [CrossRef]
- Garnier, J.-M.; Travassac, F.; Lenoble, V.; Rose, J.; Zheng, Y.; Hossain, M.; Chowdhury, S.H.; Biswas, A.K.; Ahmed, K.M.; Cheng, Z.; et al. Temporal variations in arsenic uptake by rice plants in Bangladesh: The role of iron plaque in paddy fields irrigated with groundwater. Sci. Total Environ. 2010, 408, 4185–4193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widdel, F.; Schnell, S.; Heising, S.; Ehrenreich, A.; Assmus, B.; Schink, B. Ferrous iron oxidation by anoxygenic phototrophic bacteria. Nature 1993, 362, 834–836. [Google Scholar] [CrossRef]
- Colombo, C.; Palumbo, G.; He, J.-Z.; Pinton, R.; Cesco, S. Review on iron availability in soil: Interaction of Fe minerals, plants, and microbes. J. Soils Sediments 2013, 14, 538–548. [Google Scholar] [CrossRef]
- Zelmanowitz, S.; Boyle, W.C.; Armstrong, D.E.; Park, J.K. Ability of Subsoils to Buffer Extremely Acidic Simulated Coal-Pile Leachates. J. Environ. Eng. 1995, 121, 816–823. [Google Scholar] [CrossRef]
- Aharonov-Nadborny, R.; Tsechansky, L.; Raviv, M.; Graber, E.R. Impact of spreading olive mill waste water on agricultural soils for leaching of metal micronutrients and cations. Chemosphere 2017, 179, 213–221. [Google Scholar] [CrossRef]
- El-Naggar, A.; Lee, S.S.; Awad, Y.M.; Yang, X.; Ryu, C.; Rizwan, M.; Rinklebe, J.; Tsang, D.C.; Ok, Y.S. Influence of soil properties and feedstocks on biochar potential for carbon mineralization and improvement of infertile soils. Geoderma 2018, 332, 100–108. [Google Scholar] [CrossRef]
- Freilich, S.; Kreimer, A.; Meilijson, I.; Gophna, U.; Sharan, R.; Ruppin, E. The large-scale organization of the bacterial network of ecological co-occurrence interactions. Nucleic Acids Res. 2010, 38, 3857–3868. [Google Scholar] [CrossRef]
- Deng, Y.; Jiang, Y.-H.; Yang, Y.; He, Z.; Luo, F.; Zhou, J. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Luo, X.; Chen, Y.; Ye, X.; Wang, H.; Cao, Z.; Ran, W.; Cui, Z. Succession of Composition and Function of Soil Bacterial Communities During Key Rice Growth Stages. Front. Microbiol. 2019, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, S.; Nuccio, E.E.; Shi, Z.J.; He, Z.; Zhou, J.; Firestone, M.K. The interconnected rhizosphere: High network complexity dominates rhizosphere assemblages. Ecol. Lett. 2016, 19, 926–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Gu, J.; Wang, X.; Song, W.; Zhang, K.; Sun, W.; Zhang, X.; Zhang, Y.; Li, H. Effects of Copper Addition on Copper Resistance, Antibiotic Resistance Genes, and intl1 during Swine Manure Composting. Front. Microbiol. 2017, 8, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, S.; Min, Z.; Xiuxiu, Z.; Khan, M.H.; Lifeng, L.; Hui, C. Significance of Fe(II) and environmental factors on carbon-fixing bacterial community in two paddy soils. Ecotoxicol. Environ. Saf. 2019, 182, 109456. [Google Scholar] [CrossRef]
- Becking, L.G.M.B.; Kaplan, I.R.; Moore, D. Limits of the Natural Environment in Terms of pH and Oxidation-Reduction Potentials. J. Geol. 1960, 68, 243–284. [Google Scholar] [CrossRef]
- Gupta, P.K.; Gupta, P.K.; Gupta, P.K. Methods in Environmental Analysis: Water, Soil and Air; Agrobios: Jodhpur, India, 2007. [Google Scholar]
- Vilo, C.; Dong, Q. Evaluation of the RDP Classifier Accuracy Using 16S rRNA Gene Variable Regions. Metagenomics 2012, 1, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wu, F. Diversity and Co-occurrence Patterns of Soil Bacterial and Fungal Communities in Seven Intercropping Systems. Front. Microbiol. 2018, 9, 1521. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cui, Y.; Li, X.; Yao, M. Microeco: An R package for data mining in microbial community ecology. FEMS Microbiol. Ecol. 2020, 97. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Sun, J.; Chen, D.G. Statistical Analysis of Microbiome Data with R; Springer: Singapore, 2018; Volume 847. [Google Scholar]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Crampon, M.; Bodilis, J.; Portet-Koltalo, F. Linking initial soil bacterial diversity and polycyclic aromatic hydrocarbons (PAHs) degradation potential. J. Hazard. Mater. 2018, 359, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. Int. J. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Liu, S.; Khan, M.H.; Yuan, Z.; Hussain, S.; Cao, H.; Liu, Y. Response of soil microbiome structure and its network profiles to four soil amendments in monocropping strawberry greenhouse. PLoS ONE 2021, 16, e0245180. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Pérez-Jaramillo, J.E.; De Hollander, M.; Ramírez, C.A.; Mendes, R.; Raaijmakers, J.M.; Carrión, V.J. Deciphering rhizosphere microbiome assembly of wild and modern common bean (Phaseolus vulgaris) in native and agricultural soils from Colombia. Microbiome 2019, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Li, D.; Tang, Y.; Wu, F.; Wang, J. CytoCluster: A cytoscape plugin for cluster analysis and visualization of biological networks. Int. J. Mol. Sci. 2017, 18, 1880. [Google Scholar] [CrossRef] [Green Version]
- Kögel-Knabner, I.; Amelung, W.; Cao, Z.; Fiedler, S.; Frenzel, P.; Jahn, R.; Kalbitz, K.; Kölbl, A.; Schloter, M. Biogeochemistry of paddy soils. Geoderma 2010, 157, 1–14. [Google Scholar] [CrossRef]
- Gu, S.; Yang, T.; Shao, Z.; Wang, T.; Cao, K.; Jousset, A.; Friman, V.-P.; Mallon, C.; Mei, X.; Wei, Z.; et al. Siderophore-Mediated Interactions Determine the Disease Suppressiveness of Microbial Consortia. mSystems 2020, 5, e00811-19. [Google Scholar] [CrossRef] [PubMed]
- Tiedje, J.M.; Cho, J.C.; Murray, A.; Treves, D.; Xia, B.; Zhou, J. Soil teeming with life: New frontiers for soil science. In Sustainable Management of Soil Organic Matter; CABi: Wallingford, UK, 2001; pp. 393–425. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Zhang, G.; Li, J.; Lu, H.; Zhao, W. Effects of Fe2+ concentration on biomass accumulation and energy metabolism in photosynthetic bacteria wastewater treatment. Bioresour. Technol. 2012, 119, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Munro, S.; Potts, J.; Millard, P. Influence of grass species and soil type on rhizosphere microbial community structure in grassland soils. Appl. Soil Ecol. 2007, 36, 147–155. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, G.; Jin, J.; Liu, J.; Zhang, Q.; Liu, X. Bacterial communities in soybean rhizosphere in response to soil type, soybean genotype, and their growth stage. Soil Biol. Biochem. 2009, 41, 919–925. [Google Scholar] [CrossRef]
- Yang, G.; Han, L.; Wen, J.; Zhou, S. Pseudomonas guangdongensis sp. nov., isolated from an electroactive biofilm, and emended description of the genus Pseudomonas Migula 1894. Int. J. Syst. Evol. Microbiol. 2013, 63, 4599–4605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, D.C.; Sutton, S.D.; Ringelberg, D.B. The genus Sphingomonas: Physiology and ecology. Curr. Opin. Biotechnol. 1996, 7, 301–306. [Google Scholar] [CrossRef]
- Harms, H.; Koops, H.-P.; Wehrmann, H. An ammonia-oxidizing bacterium, Nitrosovibrio tenuis nov. gen. nov. sp. Arch. Microbiol. 1976, 108, 105–111. [Google Scholar] [CrossRef]
- Schramm, A.; Davidson, S.K.; Dodsworth, J.; Drake, H.L.; Stahl, D.A.; Dubilier, N. Acidovorax-like symbionts in the nephridia of earthworms. Environ. Microbiol. 2003, 5, 804–809. [Google Scholar] [CrossRef]
- Willems, A.; Goor, M.; Thielemans, S.; Gillis, M.; Kersters, K.; De Ley, J. Transfer of Several Phytopathogenic Pseudomonas Species to Acidovorax as Acidovorax avenae subsp. avenae subsp. nov., comb. nov., Acidovorax avenae subsp. citrulli, Acidovorax avenae subsp. cattleyae, and Acidovorax konjaci. Int. J. Syst. Bacteriol. 1992, 42, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Pakostova, E.; Johnson, D.B.; Bao, Z.; MacKenzie, P.M.; Ptacek, C.J.; Blowes, D.W. Bacterial and Archaeal Diversity in Sulfide-Bearing Waste Rock at Faro Mine Complex, Yukon Territory, Canada. Geomicrobiol. J. 2020, 37, 511–519. [Google Scholar] [CrossRef]
- Tsolcha, O.N.; Tekerlekopoulou, A.G.; Akratos, C.S.; Antonopoulou, G.; Aggelis, G.; Genitsaris, S.; Moustaka-Gouni, M.; Vayenas, D. A Leptolyngbya-based microbial consortium for agro-industrial wastewaters treatment and biodiesel production. Environ. Sci. Pollut. Res. 2018, 25, 17957–17966. [Google Scholar] [CrossRef]
- Spring, S.; Rosenzweig, F. The Genera Desulfitobacterium and Desulfosporosinus: Taxonomy. Prokaryotes 2006, 4, 771–786. [Google Scholar] [CrossRef]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Imlay, J.A. Pathways of oxidative damage. Annu. Rev. Microbiol. 2003, 57, 395–418. [Google Scholar] [CrossRef]
- Lee, C.; Kim, J.Y.; Lee, W.I.; Nelson, K.L.; Yoon, J.; Sedlak, D.L. Bactericidal Effect of Zero-Valent Iron Nanoparticles on Escherichia coli. Environ. Sci. Technol. 2008, 42, 4927–4933. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Park, H.-J.; Lee, C.; Nelson, K.L.; Sedlak, D.L.; Yoon, J. Inactivation of Escherichia coli by Nanoparticulate Zerovalent Iron and Ferrous Ion. Appl. Environ. Microbiol. 2010, 76, 7668–7670. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Dong, H.; Kukkadapu, R.; Agrawal, A.; Liu, D.; Zhang, J.; Edelmann, R.E. Biological oxidation of Fe(II) in reduced nontronite coupled with nitrate reduction by Pseudogulbenkiania sp. Strain 2002. Geochim. Cosmochim. Acta 2013, 119, 231–247. [Google Scholar] [CrossRef]
- Wei, Z.; Yang, T.; Friman, V.-P.; Xu, Y.; Shen, Q.; Jousset, A. Trophic network architecture of root-associated bacterial communities determines pathogen invasion and plant health. Nat. Commun. 2015, 6, 8413. [Google Scholar] [CrossRef] [Green Version]
- Villemur, R.; Lanthier, M.; Beaudet, R.; Lépine, F. The desulfitobacterium genus. FEMS Microbiol. Rev. 2006, 30, 706–733. [Google Scholar] [CrossRef] [Green Version]
- Gu, S.; Wei, Z.; Shao, Z.; Friman, V.-P.; Cao, K.; Yang, T.; Kramer, J.; Wang, X.; Li, M.; Mei, X.; et al. Competition for iron drives phytopathogen control by natural rhizosphere microbiomes. Nat. Microbiol. 2020, 5, 1002–1010. [Google Scholar] [CrossRef]
- Yuan, H.; Huang, S.; Yuan, J.; You, Y.; Zhang, Y. Characteristics of microbial denitrification under different aeration intensities: Performance, mechanism, and co-occurrence network. Sci. Total Environ. 2020, 754, 141965. [Google Scholar] [CrossRef]
- Emerson, D.; Roden, E.; Twining, B.S. The microbial ferrous wheel: Iron cycling in terrestrial, freshwater, and marine environments. Front. Microbiol. 2012, 3, 383. [Google Scholar] [CrossRef] [Green Version]
- Theodoulou, F.L.; Kerr, I.D. ABC transporter research: Going strong 40 years on. Biochem. Soc. Trans. 2015, 43, 1033–1040. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.-L.; Yao, J.; Wang, F.; Min, N.; Gu, J.-H.; Li, Z.-F.; Sunahara, G.; Duran, R.; Solevic-Knudsen, T.; Hudson-Edwards, K.A.; et al. Bacterial diversity in typical abandoned multi-contaminated nonferrous metal(loid) tailings during natural attenuation. Environ. Pollut. 2019, 247, 98–107. [Google Scholar] [CrossRef]
- Vigneron, A.; Cruaud, P.; Alsop, E.; de Rezende, J.R.; Head, I.M.; Tsesmetzis, N. Beyond the tip of the iceberg; a new view of the diversity of sulfite-and sulfate-reducing microorganisms. ISME J. 2018, 12, 2096–2099. [Google Scholar] [CrossRef]
- Huang, B.; Yu, K.; Gambrell, R.P. Effects of ferric iron reduction and regeneration on nitrous oxide and methane emissions in a rice soil. Chemosphere 2009, 74, 481–486. [Google Scholar] [CrossRef]
- Furukawa, Y.; Inubushi, K. Feasible suppression technique of methane emission from paddy soil by iron amendment. Nutr. Cycl. Agroecosystems 2002, 64, 193–201. [Google Scholar] [CrossRef]
- Bharali, A.; Baruah, K.K.; Gogoi, N. Methane emission from irrigated rice ecosystem: Relationship with carbon fixation, partitioning and soil carbon storage. Paddy Water Environ. 2016, 15, 221–236. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Liu, S.; Hussain, S.; Li, L.; Baiome, B.A.; Xiao, S.; Cao, H. Fe(II) Addition Drives Soil Bacterial Co-Ocurrence Patterns and Functions Mediated by Anaerobic and Chemoautotrophic Taxa. Microorganisms 2022, 10, 547. https://doi.org/10.3390/microorganisms10030547
Zhang C, Liu S, Hussain S, Li L, Baiome BA, Xiao S, Cao H. Fe(II) Addition Drives Soil Bacterial Co-Ocurrence Patterns and Functions Mediated by Anaerobic and Chemoautotrophic Taxa. Microorganisms. 2022; 10(3):547. https://doi.org/10.3390/microorganisms10030547
Chicago/Turabian StyleZhang, Chenyang, Senlin Liu, Sarfraz Hussain, Lifeng Li, Baiome Abdelmaguid Baiome, Shuiqing Xiao, and Hui Cao. 2022. "Fe(II) Addition Drives Soil Bacterial Co-Ocurrence Patterns and Functions Mediated by Anaerobic and Chemoautotrophic Taxa" Microorganisms 10, no. 3: 547. https://doi.org/10.3390/microorganisms10030547
APA StyleZhang, C., Liu, S., Hussain, S., Li, L., Baiome, B. A., Xiao, S., & Cao, H. (2022). Fe(II) Addition Drives Soil Bacterial Co-Ocurrence Patterns and Functions Mediated by Anaerobic and Chemoautotrophic Taxa. Microorganisms, 10(3), 547. https://doi.org/10.3390/microorganisms10030547