
Characterization of Bifidobacterium asteroides Isolates

 ,
,  ,
,  ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reference Strains and Culture Conditions
2.2. Identification of Isolates
2.3. Safety Assessment
2.3.1. Antimicrobial Susceptibility and MIC Determination
2.3.2. PCR Assay on Virulence Factors
2.3.3. Other Assessments: Hemolytic Activity and DNase and Gelatinase Activities
2.4. Hydrophobicity, Auto-Aggregation and Co-Aggregation Abilities
2.5. Adhesion on Abiotic Surface
2.6. Alignment for VanZ Putative Gene
2.7. Antagonistic Activity against Pathogens
2.7.1. Agar Diffusion Assay
2.7.2. Antibacterial Activity
2.8. Statistical Analysis
3. Results
3.1. Enzymatic Profile
3.2. Antibiotic Resistance Profile, Virulence Factors and Other Biochemical Properties
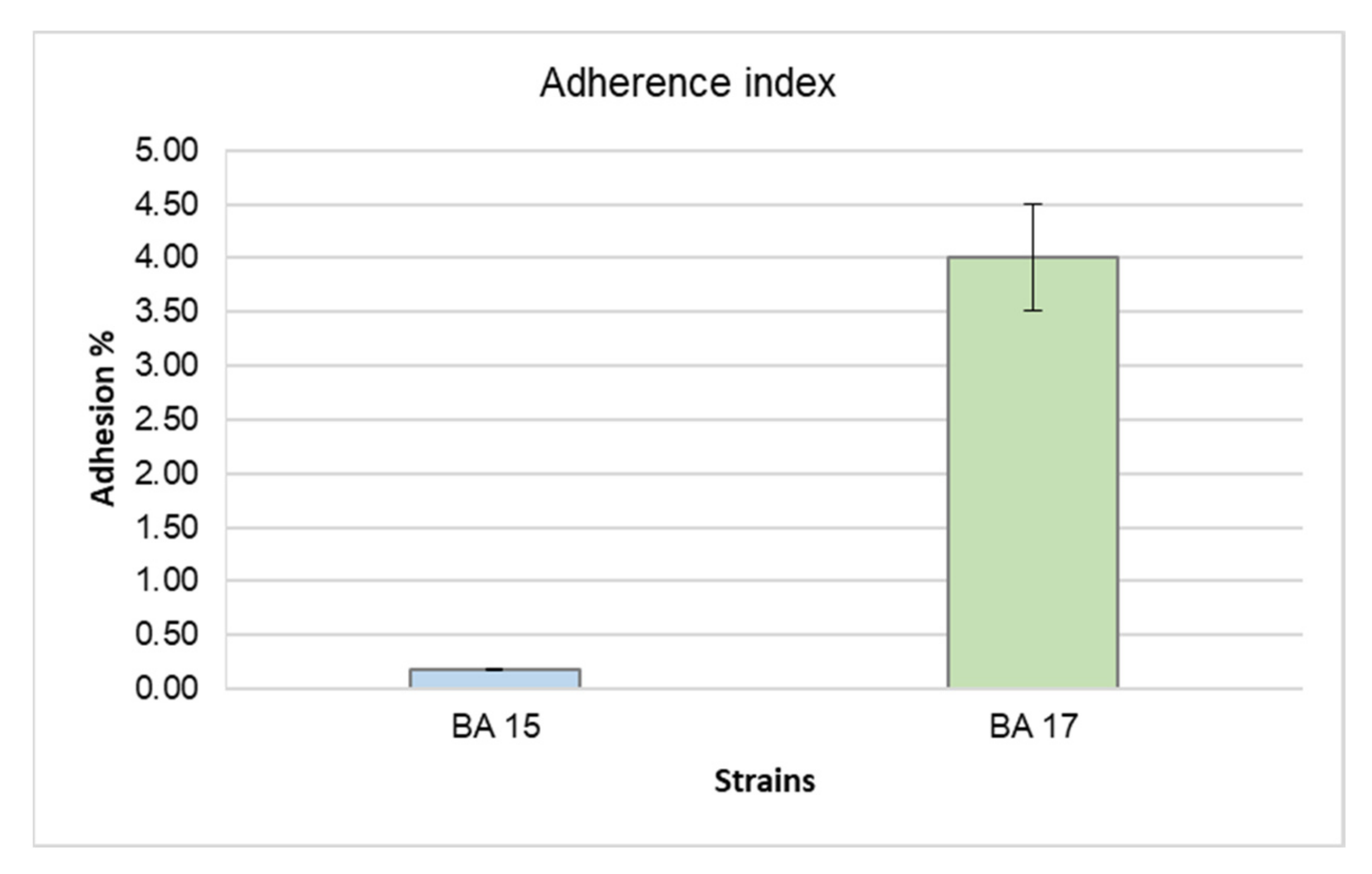
3.3. Adhesion to Abiotic Surfaces
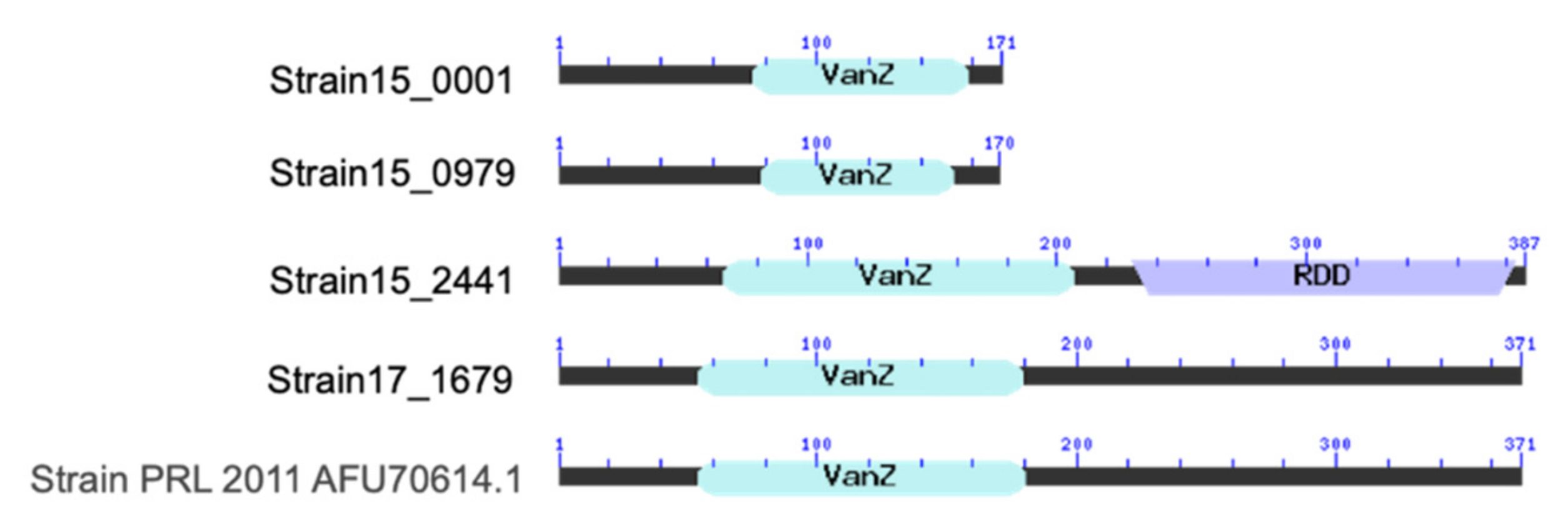
3.4. Alignment for VanZ Putative Gene
3.5. Antibacterial Activity
3.6. Auto-Aggregation, Co-Aggregation and Hydrophobicity Abilities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the Future of Probiotics and Prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Biavati, B.; Matterelli, P. The Family Bifidobacteriaceae in The Prokaryotes, 3rd ed.; Springer: New York, NY, USA, 2006; Volume 3, Chapter 1.1.2; pp. 331–336. [Google Scholar]
- Okamoto, M.; Benno, Y.; Leung, K.-P.; Maeda, N. Bifidobacterium tsurumiense sp. nov., from hamster dental plaque. Int. J. Syst. Evol. Microbiol. 2008, 58, 144–148. [Google Scholar] [CrossRef] [Green Version]
- Ventura, M.; Turroni, F.; Zomer, A.; Foroni, E.; Giubellini, V.; Bottacini, F.; Canchaya, C.; Claesson, M.; He, F.; Mantzourani, M.; et al. The Bifidobacterium dentium Bd1 Genome Sequence Reflects Its Genetic Adaptation to the Human Oral Cavity. PLoS Genet. 2009, 5, e1000785. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Makino, H.; Sasamoto, M.; Kudo, Y.; Fujimoto, J.; Demberel, S. Bifidobacterium mongoliense sp. nov., from airag, a traditional fermented mare’s milk product from Mongolia. Int. J. Syst. Evol. Microbiol. 2009, 59, 1535–1540. [Google Scholar] [CrossRef]
- Arzamasov, A.A.; Van Sinderen, D.; Rodionov, D.A. Comparative Genomics Reveals the Regulatory Complexity of Bifidobacterial Arabinose and Arabino-Oligosaccharide Utilization. Front. Microbiol. 2018, 9, 776. [Google Scholar] [CrossRef] [PubMed]
- Bottacini, F.; Milani, C.; Turroni, F.; Sánchez, B.; Foroni, E.; Duranti, S.; Serafini, F.; Viappiani, A.; Strati, F.; Ferrarini, A.; et al. Bifidobacterium asteroides PRL2011 Genome Analysis Reveals Clues for Colonization of the Insect Gut. PLoS ONE 2012, 7, e44229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberoni, D.; Gaggìa, F.; Baffoni, L.; Modesto, M.M.; Biavati, B.; Di Gioia, D. Bifidobacterium xylocopae sp. nov. and Bifidobacterium aemilianum sp. nov., from the carpenter bee (Xylocopa violacea) digestive tract. Syst. Appl. Microbiol. 2019, 42, 205–216. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Opinion of the Scientific Committee on a request from EFSA on the introduction of a Qualified Presumption of Safety (QPS) approach for assessment of selected microorganisms referred to EFSA. EFSA J. 2007, 587, 1–16. [Google Scholar] [CrossRef]
- Sornplang, P.; Piyadeatsoontorn, S. Probiotic isolates from unconventional sources: A review. J. Anim. Sci. Technol. 2016, 58, 26. [Google Scholar] [CrossRef] [Green Version]
- Pennacchia, C.; Vaughan, E.; Villani, F. Potential probiotic Lactobacillus strains from fermented sausages: Further investigations on their probiotic properties. Meat Sci. 2006, 73, 90–101. [Google Scholar] [CrossRef]
- Chang, J.-H.; Shim, Y.Y.; Cha, S.-K.; Chee, K.M. Probiotic characteristics of lactic acid bacteria isolated from kimchi. J. Appl. Microbiol. 2010, 109, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Vitali, B.; Minervini, G.; Rizzello, C.G.; Spisni, E.; Maccaferri, S.; Brigidi, P.; Gobbetti, M.; Di Cagno, R. Novel probiotic candidates for humans isolated from raw fruits and vegetables. Food Microbiol. 2012, 31, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daisley, B.A.; Chmiel, J.A.; Pitek, A.P.; Thompson, G.J.; Reid, G. Missing Microbes in Bees: How Systematic Depletion of Key Symbionts Erodes Immunity. Trends Microbiol. 2020, 28, 1010–1021. [Google Scholar] [CrossRef] [PubMed]
- Bonilla-Rosso, G.; Engel, P. Functional roles and metabolic niches in the honey bee gut microbiota. Curr. Opin. Microbiol. 2018, 43, 69–76. [Google Scholar] [CrossRef] [Green Version]
- European Safety Authority Home Page. Available online: https://www.efsa.europa.eu/en/science/scientific-committee-and-panels/feedap (accessed on 7 February 2022).
- Janashia, I.; Carminati, D.; Rossetti, L.; Zago, M.; Fornasari, M.E.; Haertlé, T.; Chanishvili, N.; Giraffa, G. Characterization of fructophilic lactic microbiota of Apis mellifera from the Caucasus Mountains. Ann. Microbiol. 2016, 66, 1387–1395. [Google Scholar] [CrossRef]
- Mancino, W.; Duranti, S.; Mancabelli, L.; Longhi, G.; Anzalone, R.; Milani, C.; Lugli, G.A.; Carnevali, L.; Statello, R.; Sgoifo, A.; et al. Bifidobacterial Transfer from Mother to Child as Examined by an Animal Model. Microorganisms 2019, 7, 293. [Google Scholar] [CrossRef] [Green Version]
- Caggia, C.; De Angelis, M.; Pitino, I.; Pino, A.; Randazzo, C. Probiotic features of Lactobacillus strains isolated from Ragusano and Pecorino Siciliano cheeses. Food Microbiol. 2015, 50, 109–117. [Google Scholar] [CrossRef]
- Inturri, R.; Stivala, A.; Blandino, G. Microbiological characteristics of the probiotic strains B. longum BB536 and L. rhamnosus HN001 used in combination. Minerva Gastroenterol. Dietol. 2015, 61, 191–197. [Google Scholar]
- Blandino, G.; Fazio, D.; Petronio, G.P.; Inturri, R.; Tempera, G.; Furneri, P.M. Labeling quality and molecular characterization studies of products containing Lactobacillus spp. strains. Int. J. Immunopathol. Pharmacol. 2016, 29, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Inturri, R.; Stivala, A.; Furneri, P.M.; Blandino, G. Growth and adhesion to HT-29 cells inhibition of Gram-negatives by Bifidobacterium longum BB536 e Lactobacillus rhamnosus HN001 alone and in combination. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4943–4949. [Google Scholar] [PubMed]
- Inturri, R.; Trovato, L.; Volti, G.L.; Oliveri, S.; Blandino, G. In vitro inhibitory activity of Bifidobacterium longum BB536 and Lactobacillus rhamnosus HN001 alone or in combination against bacterial and Candida reference strains and clinical isolates. Heliyon 2019, 5, e02891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruas-Madiedo, P.; Gueimonde, M.; Margolles, A.; de los Reyes-Gavilan, C.; Salminen, S. Exopolysaccharides Produced by Probiotic Strains Modify the Adhesion of Probiotics and Enteropathogens to Human Intestinal Mucus. J. Food Prot. 2006, 69, 2011–2015. [Google Scholar] [CrossRef]
- Foroni, E.; Serafini, F.; Amidani, D.; Turroni, F.; He, F.; Bottacini, F.; O’Connell Motherway, M.; Viappiani, A.; Zhang, Z.; Rivetti, C.; et al. Genetic analysis and morphological identification of pilus-like structures in members of the genus Bifidobacterium. Microb. Cell Fact. 2011, 10, S16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killer, J.; Kopečný, J.; Mrázek, J.; Rada, V.; Dubná, S.; Marounek, M. Bifidobacteria in the digestive tract of bumblebees. Anaerobe 2010, 16, 165–170. [Google Scholar] [CrossRef]
- Pino, A.; Bartolo, E.; Caggia, C.; Cianci, A.; Randazzo, C.L. Detection of vaginal lactobacilli as probiotic candidates. Sci. Rep. 2019, 9, 3355. [Google Scholar] [CrossRef]
- Ruas-Madiedo, P.; Hernández-Barranco, A.; Margolles, A.; de los Reyes-Gavilán, C.G. A Bile Salt-Resistant Derivative of Bifidobacterium animalis Has an Altered Fermentation Pattern When Grown on Glucose and Maltose. Appl. Environ. Microbiol. 2005, 71, 6564–6570. [Google Scholar] [CrossRef] [Green Version]
- Arboleya, S.; Ruas-Madiedo, P.; Margolles, A.; Solís, G.; Salminen, S.; de los Reyes-Gavilán, C.G.; Gueimonde, M. Characterization and in vitro properties of potentially probiotic Bifidobacterium strains isolated from breast-milk. Int. J. Food Microbiol. 2011, 149, 28–36. [Google Scholar] [CrossRef] [Green Version]
- ISO 10932/IDF 233; Milk and Milk Products—Determination of the Minimal Inhibitory Concentration (MIC) of Antibiotics Applicable to Bifidobacteria and Non-Enterococcal Lactic Acid Bacteria (LAB). ISO: International Organization for Standardization: Geneva, Switzerland, 2010.
- Russo, N.; Caggia, C.; Pino, A.; Coque, T.M.; Arioli, S.; Randazzo, C.L. Enterococcus spp. in Ragusano PDO and Pecorino Siciliano cheese types: A snapshot of their antibiotic resistance distribution. Food Chem. Toxicol. 2018, 120, 277–286. [Google Scholar] [CrossRef]
- Wattam, A.R.; Abraham, D.; Dalay, O.; Disz, T.L.; Driscoll, T.; Gabbard, J.L.; Gillespie, J.J.; Gough, R.; Hix, D.; Kenyon, R.; et al. PATRIC, the bacterial bioinformatics database and analysis resource. Nucleic Acids Res. 2014, 42, D581–D591. [Google Scholar] [CrossRef] [Green Version]
- Vankerckhoven, V.; Van Autgaerden, T.; Vael, C.; Lammens, C.; Chapelle, S.; Rossi, R.; Jabes, D.; Goossens, H. Development of a Multiplex PCR for the Detection of asa1, gelE, cylA, esp, and hyl Genes in Enterococci and Survey for Virulence Determinants among European Hospital Isolates of Enterococcus faecium. J. Clin. Microbiol. 2004, 42, 4473–4479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depardieu, F.; Perichon, B.; Courvalin, P. Detection of the van Alphabet and Identification of Enterococci and Staphylococci at the Species Level by Multiplex PCR. J. Clin. Microbiol. 2004, 42, 5857–5860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pino, A.; Russo, N.; Van Hoorde, K.; De Angelis, M.; Sferrazzo, G.; Randazzo, C.L.; Caggia, C. Piacentinu Ennese PDO Cheese as Reservoir of Promising Probiotic Bacteria. Microorganisms 2019, 7, 254. [Google Scholar] [CrossRef] [Green Version]
- Pino, A.; Rapisarda, A.M.C.; Vitale, S.G.; Cianci, S.; Caggia, C.; Randazzo, C.L.; Cianci, A. A clinical pilot study on the effect of the probiotic Lacticaseibacillus rhamnosus TOM 22.8 strain in women with vaginal dysbiosis. Sci. Rep. 2021, 11, 2592. [Google Scholar] [CrossRef]
- Gross, M.; Cramton, S.E.; Götz, F.; Peschel, A. Key Role of Teichoic Acid Net Charge in Staphylococcus aureus Colonization of Artificial Surfaces. Infect. Immun. 2001, 69, 3423–3426. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Randazzo, C.L.; Russo, N.; Pino, A.; Mazzaglia, A.; Ferrante, M.; Conti, G.O.; Caggia, C. Effects of selected bacterial cultures on safety and sensory traits of Nocellara Etnea olives produced at large factory scale. Food Chem. Toxicol. 2018, 115, 491–498. [Google Scholar] [CrossRef]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.A.G.; Tsakalidou, E.; Nychas, G.-J.E.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 7th ed.; CLSI Document M7-A7; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2006; ISBN 1-56238-587-9. [Google Scholar]
- EFSA. Technical guidance prepared by the Panel on Additives and Products or Substances used in Animal Feed (FEEDAP) on the update of the criteria used in the assessment of bacterial resistance to antibiotics of human or veterinary importance. EFSA J. 2008, 732, 1–15. [Google Scholar] [CrossRef]
- Alessandri, G.; Ossiprandi, M.C.; Mac Sharry, J.; Van Sinderen, D.; Ventura, M. Bifidobacterial Dialogue with Its Human Host and Consequent Modulation of the Immune System. Front. Immunol. 2019, 10, 2348. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.; Nethery, M.A.; Hidalgo-Cantabrana, C.; Barrangou, R. Comprehensive Mining and Characterization of CRISPR-Cas Systems in Bifidobacterium. Microorganisms 2020, 8, 720. [Google Scholar] [CrossRef] [PubMed]
- Odamaki, T.; Bottacini, F.; Kato, K.; Mitsuyama, E.; Yoshida, K.; Horigome, A.; Xiao, J.-Z.; Van Sinderen, D. Genomic diversity and distribution of Bifidobacterium longum subsp. longum across the human lifespan. Sci. Rep. 2018, 8, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turroni, F.; Duranti, S.; Milani, C.; Lugli, G.A.; Van Sinderen, D.; Ventura, M. Bifidobacterium bifidum: A Key Member of the Early Human Gut Microbiota. Microorganisms 2019, 7, 544. [Google Scholar] [CrossRef] [Green Version]
- Lugli, G.A.; Alessandri, G.; Milani, C.; Mancabelli, L.; Ruiz, L.; Fontana, F.; Borragán, S.; González, A.; Turroni, F.; Ossiprandi, M.C.; et al. Evolutionary development and co-phylogeny of primate-associated bifidobacteria. Environ. Microbiol. 2020, 22, 3375–3393. [Google Scholar] [CrossRef] [PubMed]
- Kešnerová, L.; Emery, O.; Troilo, M.; Liberti, J.; Erkosar, B.; Engel, P. Gut microbiota structure differs between honeybees in winter and summer. ISME J. 2020, 14, 801–814. [Google Scholar] [CrossRef] [Green Version]
- Kwong, W.K.; Moran, N.A. Gut microbial communities of social bees. Nat. Rev. Microbiol. 2016, 14, 374–384. [Google Scholar] [CrossRef]
- Leahy, S.; Higgins, D.; Fitzgerald, G.; Van Sinderen, D. Getting better with bifidobacteria. J. Appl. Microbiol. 2005, 98, 1303–1315. [Google Scholar] [CrossRef]
- Nowak, A.; Szczuka, D.; Górczyńska, A.; Motyl, I.; Kręgiel, D. Characterization of Apis mellifera Gastrointestinal Microbiota and Lactic Acid Bacteria for Honeybee Protection—A Review. Cells 2021, 10, 701. [Google Scholar] [CrossRef]
- FAO; WHO. Probiotics in food. In Health and Nutritional Properties and Guidelines for Evaluation; Food and Agriculture Organization: Rome, Italy, 2006; ISBN 92-5-105513-0. [Google Scholar]
- EFSA. Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Guidance on the Assessment of Bacterial Susceptibility to Antimicrobials of Human and Veterinary Importance. EFSA J. 2012, 10, 2740. [Google Scholar] [CrossRef]
- Inturri, R.; Ventura, M.; Ruas-Madiedo, P.; Lugli, G.A.; Blandino, G. Complete Genome Sequence of Bifidobacterium longum W11 (LMG P-21586), Used as a Probiotic Strain. Genome Announc. 2017, 5, e01659-16. [Google Scholar] [CrossRef] [Green Version]
- Inturri, R.; Mangano, K.; Santagati, M.; Intrieri, M.; Di Marco, R.; Blandino, G. Immunomodulatory Effects of Bifidobacterium longum W11 Produced Exopolysaccharide on Cytokine Production. Curr. Pharm. Biotechnol. 2017, 18, 883–889. [Google Scholar] [CrossRef] [PubMed]
- Marras, L.; Caputo, M.; Bisicchia, S.; Soato, M.; Bertolino, G.; Vaccaro, S.; Inturri, R. The Role of Bifidobacteria in Predictive and Preventive Medicine: A Focus on Eczema and Hypercholesterolemia. Microorganisms 2021, 9, 836. [Google Scholar] [CrossRef] [PubMed]
- Gueimonde, M.; Delgado, S.; Mayo, B.; Ruas-Madiedo, P.; Margolles, A.; de los Reyes-Gavilán, C.G. Viability and diversity of probiotic Lactobacillus and Bifidobacterium populations included in commercial fermented milks. Food Res. Int. 2004, 37, 839–850. [Google Scholar] [CrossRef]
- Awasti, N.; Tomar, S.; Pophaly, S.D.; Poonam; Lule, V.K.; Singh, T.P.; Anand, S. Probiotic and functional characterization of bifidobacteria of Indian human origin. J. Appl. Microbiol. 2016, 120, 1021–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ku, S.; Yang, S.; Lee, H.H.; Choe, D.; Johnston, T.V.; Ji, G.E.; Park, M.S. Biosafety assessment of Bifidobacterium animalis subsp. lactis AD011 used for human consumption as a probiotic microorganism. Food Control 2019, 117, 106985. [Google Scholar] [CrossRef]
- Gueimonde, M.; Sánchez, B.; de Los Reyes-Gavilán, C.G.; Margolles, A. Antibiotic resistance in probiotic bacteria. Front. Microbiol. 2013, 4, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campedelli, I.; Mathur, H.; Salvetti, E.; Clarke, S.; Rea, M.C.; Torriani, S.; Ross, R.P.; Hill, C.; O’Toole, P.W. Genus-Wide Assessment of Antibiotic Resistance in Lactobacillus spp. Appl. Environ. Microbiol. 2019, 85, e01738-18. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.J.; Ku, S.; Kim, S.Y.; Lee, H.H.; Jin, H.; Kang, S.; Li, R.; Johnston, T.V.; Park, M.S.; Ji, G.E. Safety Evaluations of Bifidobacterium bifidum BGN4 and Bifidobacterium longum BORI. Int. J. Mol. Sci. 2018, 19, 1422. [Google Scholar] [CrossRef] [Green Version]
- Sirichoat, A.; Flórez, A.B.; Vázquez, L.; Buppasiri, P.; Panya, M.; Lulitanond, V.; Mayo, B. Antibiotic Susceptibility Profiles of Lactic Acid Bacteria from the Human Vagina and Genetic Basis of Acquired Resistances. Int. J. Mol. Sci. 2020, 21, 2594. [Google Scholar] [CrossRef] [Green Version]
- Fouhy, F.; O’Connell Motherway, M.; Fitzgerald, G.F.; Ross, R.; Stanton, C.; Van Sinderen, D.; Cotter, P.D. In Silico Assigned Resistance Genes Confer Bifidobacterium with Partial Resistance to Aminoglycosides but Not to Β-Lactams. PLoS ONE 2013, 8, e82653. [Google Scholar] [CrossRef] [Green Version]
- European Committee on Antimicrobial Susceptibility Testing. EUCAST 2019. The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 9.0, 2019. Last Accession MIC Distributions for Bifidobacterium spp. 2021. Available online: http://www.eucast.org (accessed on 7 February 2022).
- Gueimonde, M.; Flórez, A.B.; van Hoek, A.H.A.M.; Stuer-Lauridsen, B.; Strøman, P.; de los Reyes-Gavilán, C.G.; Margolles, A. Genetic Basis of Tetracycline Resistance in Bifidobacterium animalis subsp. lactis. Appl. Environ. Microbiol. 2010, 76, 3364–3369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatahi-Bafghi, M. Antibiotic resistance genes in the Actinobacteria phylum. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1599–1624. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, A.; Robertson, K.; Yung, A.; Que, M.; Randall, H.; Wellalagodage, D.; Cox, T.; Robertson, D.; Chi, C.; Sun, J. Efficacy of Probiotics in Patients of Cardiovascular Disease Risk: A Systematic Review and Meta-analysis. Curr. Hypertens. Rep. 2020, 22, 74. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Meriluoto, J.; Salminen, S. Adhesion and aggregation properties of probiotic and pathogen strains. Eur. Food Res. Technol. 2008, 226, 1065–1073. [Google Scholar] [CrossRef]
- Begum, S.B.; Roobia, R.R.; Karthikeyan, M.; Murugappan, R. Validation of nutraceutical properties of honey and probiotic potential of its innate microflora. LWT 2015, 60, 743–750. [Google Scholar] [CrossRef]
- Rahman, M.; Kim, W.-S.; Kumura, H.; Shimazaki, K.-I. Autoaggregation and surface hydrophobicity of bifidobacteria. World J. Microbiol. Biotechnol. 2008, 24, 1593–1598. [Google Scholar] [CrossRef]
- Kesen, M.A.; Aiyegoro, O.A. Beneficial Characteristics and Evaluation Criteria of Probiotics. Int. J. Food Biosci. 2018, 1, 19–26. [Google Scholar]
- Collado, M.; Meriluoto, J.; Salminen, S. Role of commercial probiotic strains against human pathogen adhesion to intestinal mucus. Lett. Appl. Microbiol. 2007, 45, 454–460. [Google Scholar] [CrossRef]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [Green Version]
- De Vuyst, L.; Avonts, L.; Makras, L. Probiotics, prebiotics and gut health. In Functional Foods, Ageing and Degenerative Disease; Remacle, C., Reusens, B., Eds.; Woodhead Publishing: Cambridge, UK, 2004; Chapter 17; pp. 416–482. [Google Scholar]
- Cheikhyoussef, A.; Pogori, N.; Chen, W.; Zhang, H. Antimicrobial proteinaceous compounds obtained from bifidobacteria: From production to their application. Int. J. Food Microbiol. 2008, 125, 215–222. [Google Scholar] [CrossRef]
- Igbafe, J.; Kilonzo-Nthenge, A.; Nahashon, S.; Mafiz, A.; Nzomo, M. Probiotics and Antimicrobial Effect of Lactiplantibacillus plantarum, Saccharomyces cerevisiae, and Bifidobacterium longum against Common Foodborne Pathogens in Poultry. Agriculture 2020, 10, 368. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Pino, A.; Ricciardi, L.; Romano, C.; Comito, D.; Arena, E.; Saitta, S.; Caggia, C. Probiotic supplementation in systemic nickel allergy syndrome patients: Study of its effects on lactic acid bacteria population and on clinical symptoms. J. Appl. Microbiol. 2015, 118, 202–211. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Virulence Factor | Primer Name | Oligonucleotide Sequence (5′ to 3′) | Product Size (bp) |
|---|---|---|---|---|
| asa1 | aggregation substance | ASAfw ASArw | GCACGCTATTACGAACTATGA TAAGAAAGAACATCACCACGA | 375 |
| cylA | cytolysin | CYTfw CYTrw | TATGACAATGCTTTTTGGGAT AGATGCACCCGAAATAATATA | 213 |
| gelE | gelatinase | GELfw GELrw | ATAGACAATGCTTTTTGGGAT AGATGCACCCGAAATAATATA | 213 |
| hyl | hyaluronidase | HYLfw HYLrw | ACAGAAGAGCTGCAGGAAATG GACTGACGTCCAAGTTTCCAA | 276 |
| esp | surface protein | SPfw SPrw | AGATTTCATCTTTGATTCTTGG AATTGATTCTTTAGCATCTGG | 688 |
| Reaction/Enzyme | BA15 | BA17 | Bifidobacterium spp. (% of Positive Reaction) |
|---|---|---|---|
| Urease | − | − | 0 |
| Arginine dehydrolase | + | + | 100 |
| α-galactosidase | + | + | 100 |
| β-galactosidase | − | − | 9 |
| β-galactosidase-6-phosphate | + | + | 100 |
| α-glucosidase | + | + | 91 |
| β-glucosidase | + | − | 45 |
| α-arabinosidase | − | − | 0 |
| β-glucuronidase | + | + | 64 |
| N-acetyl-β-glucosaminidase | + | + | 99 |
| Mannose fermentation | + | + | 93 |
| Raffinose fermentation | − | − | 0 |
| Glutamic acid decarboxylase | − | − | 0 |
| α-fucosidase | − | − | 9 |
| Reduction of nitrates | − | − | 1 |
| Indole production | − | − | 5 |
| Alkaline phosphatase | + | + | 100 |
| Arginine arylamidase | + | + | 99 |
| Proline arylamidase | − | − | 27 |
| Leucyl glycine arylamidase | + | + | 99 |
| Phenylalanine arylamidase | + | + | 91 |
| Leucine arylamidase | − | − | 9 |
| Pyroglutamic acid arylamidase | + | + | 99 |
| Tyrosine arylamidase | + | − | 64 |
| Alanine arylamidase | + | + | 99 |
| Glycine arylamidase | + | + | 91 |
| Histidine arylamidase | − | − | 1 |
| Glutamyl Glutamic Acid Arylaminidase | + | + | 91 |
| Serine arylaminidase | − | − | 0 |
| Biochemical Reactions | BA15 | BA17 |
|---|---|---|
| NH3 from arginine | − | − |
| Gelatin liquefaction | − | − |
| Indole production | − | − |
| Glucosidase | + | + |
| Xylose | − | − |
| D-Fructose | + | + |
| D-Galactose | + | + |
| Maltose | − | − |
| Trehalose | − | − |
| D-Melibiose | + | + |
| Mannitol | − | − |
| Salicin | − | − |
| Sorbitol | − | − |
| L-Arabinose | − | + |
| Raffinose | − | − |
| D-Ribose | − | − |
| Lactose | + | + |
| Inulin | − | − |
| Cellobiose | − | − |
| Melezitose | − | − |
| AMP (4) * | VAN (2) * | GEN (16) * | STRE (32) * | ERY (1) * | CLI (1) * | TET (8) * | CHL (4) * | |
|---|---|---|---|---|---|---|---|---|
| Tested Range (µg/mL) | ||||||||
| STRAINS | (0.5–16) | (0.5–16) | (4–128) | (8–256) | (0.25–8) | (0.25–8) | (2–64) | (1–64) |
| BB12 | <0.5 | 0.5 | 128 R | 128 R | 0.25 | <0.25 | 16 R | 2 |
| BA15 | 16 R | >16 R | 4 | 8 | 0.25 | 0.25 | 2 | 64 R |
| BA17 | 16 R | >16 R | <4 | <8 | <0.25 | <0.25 | <2 | 64 R |
| Strains | E. coli ATCC 25922 | E. coli ATCC 9637 | S. typhimurium ATCC 14028 | S. aureus ATCC 6538 | S. aureus ATCC 29213 |
|---|---|---|---|---|---|
| BB12 | ++ | ++ | ++ | ++ | ++ |
| BA15 | ++ | + | ++ | - | + |
| BA17 | ++ | + | ++ | ++ | + |
| Strains | % Identity | % Gaps | Identical | Gap Count | Gap Length | Score | Length | |
|---|---|---|---|---|---|---|---|---|
| PRL2011 | Strain15_0001 | 46.5 | 36.9 | 303 | 55 | 240 | 348 | 651 |
| Strain15_0979 | 49.0 | 32.8 | 329 | 62 | 220 | 379 | 671 | |
| Strain15_2441 | 49.8 | 31.5 | 649 | 109 | 409 | 872 | 1297 | |
| Strain17_1679 | 94.0 | 0.0 | 1048 | 0 | 0 | 4972 | 1115 |
| Auto-A% | Co-A% | H% | |||||
|---|---|---|---|---|---|---|---|
| Strains | S. aureus ATCC 6538 | S. typhimurium ATCC 14028 | E. coli ATCC 25922 | S. enteritidis ATCC 13076 | Listeria monocytogenes ATCC 12466 | ||
| BB 12 | 36.70 ± 0.11 a | 40.05 ± 0.17 b | 10.00 ± 0.19 a | 23.50 ± 0.13 b | 34.50 ± 0.12 c | 15.60 ± 0.13 a | 84.50 ± 0.13 c |
| BA 15 | 34.13 ± 0.13 a | 14.22 ± 0.12 a | 19.25 ± 0.17 b | 15.97 ± 0.12 a | 15.97 ± 0.15 a | 19.18 ± 0.15 b | 59.67 ± 0.14 a |
| BA 17 | 33.11 ± 0.17 a | 16.67 ± 0.11 a | 13.33 ± 0.18 a | 28.31 ± 0.18 b | 26.25 ± 0.17 b | 19.73 ± 0.12 b | 79.15 ± 0.11 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pino, A.; Benkaddour, B.; Inturri, R.; Amico, P.; Vaccaro, S.C.; Russo, N.; Vaccalluzzo, A.; Agolino, G.; Caggia, C.; Miloud, H.; et al. Characterization of Bifidobacterium asteroides Isolates. Microorganisms 2022, 10, 655. https://doi.org/10.3390/microorganisms10030655
Pino A, Benkaddour B, Inturri R, Amico P, Vaccaro SC, Russo N, Vaccalluzzo A, Agolino G, Caggia C, Miloud H, et al. Characterization of Bifidobacterium asteroides Isolates. Microorganisms. 2022; 10(3):655. https://doi.org/10.3390/microorganisms10030655
Chicago/Turabian StylePino, Alessandra, Bachir Benkaddour, Rosanna Inturri, Pietro Amico, Susanna C. Vaccaro, Nunziatina Russo, Amanda Vaccalluzzo, Gianluigi Agolino, Cinzia Caggia, Hadadji Miloud, and et al. 2022. "Characterization of Bifidobacterium asteroides Isolates" Microorganisms 10, no. 3: 655. https://doi.org/10.3390/microorganisms10030655
APA StylePino, A., Benkaddour, B., Inturri, R., Amico, P., Vaccaro, S. C., Russo, N., Vaccalluzzo, A., Agolino, G., Caggia, C., Miloud, H., & Randazzo, C. L. (2022). Characterization of Bifidobacterium asteroides Isolates. Microorganisms, 10(3), 655. https://doi.org/10.3390/microorganisms10030655