
Harnessing the Algal Chloroplast for Heterologous Protein Production




Abstract
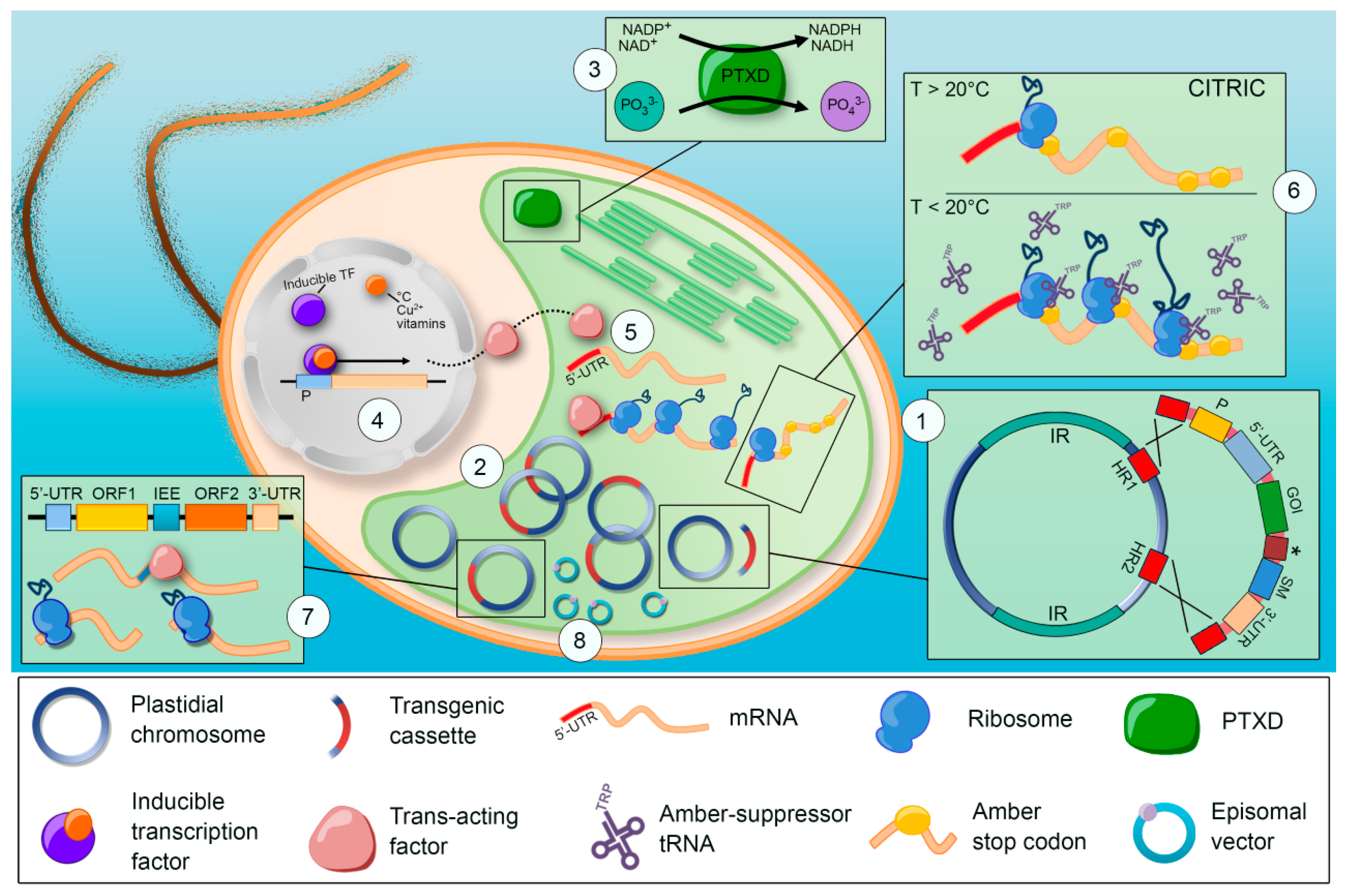
:1. Introduction
2. Chloroplast Genetics in a Nutshell
3. Plastome Engineering in a Model Green Alga
3.1. Selection Strategies
3.2. Cis-Acting Regulatory Elements
3.3. Reporter Genes

4. Examples of Recombinant Products
4.1. Microalgal Molecular Pharming
4.2. Enzymes, Metabolites, and Valorization of Lignocellulose

{kind=link}
| Expressed Product | Category/Application | Promoters and Cis-Acting Elements | Selection System | Highlights | References |
|---|---|---|---|---|---|
| Biopharmaceuticals/green biologics for human health | |||||
| Bovine mammary-associated serum amyloid protein (M-SAA) | Prophylaxis of bacterial and viral infections | PpsbA psbA 3′-UTR | aadA1 gene (aminoglycoside adenyltransferase)—Spectinomycin | Accumulation of 5% of TSP | |
| Bioactive peptides from milk proteins connected | Antihypertensive, antimicrobial, immunomodulatory, antioxidant, and hypocholesterolemic activities | PrbcL or PatpA rbcL 3′-UTR | aadA1 gene—Spectinomycin | Chimeric peptides linked by gastrointestinal proteases cleavage sites | [124] |
| Bovine milk amyloid A protein (mammary-associated serum amyloid A, M-SAA) | Prophylaxis of bacterial and viral infections | Combinations of various endogenous and heterologous promoters (psbA, atpA, tufA, and psbD) | Phototrophic rescue of ΔpsbA strain and aphA6 gene (aminoglycoside 3′-phosphotransferase)—Kanamycin | Avoidance of psbA auto-attenuation and photoautotrophic growth in 100 L | [99] |
| Tenth human fibronectin type III domain (10FN3) | Extracellular matrix glycoprotein with roles in cell adhesion, migration, growth, and differentiation | PpsbA psbA 3′-UTR | aphA6 gene—Kanamycin | A carboxy-terminal fusion to the M-SAA protein enabled synthesis of otherwise non detectable products | [126,127] |
| Fourteenth human fibronectin type III domain (14FN3) | Antibody mimic | ||||
| Human vascular endothelial growth factor (VEGF) isoform 121 | Treatment of pulmonary emphysema | ||||
| High mobility group protein B1 (HMGB1) | Mediator of wound healing | ||||
| Human growth hormone (hGH) | Growth hormone deficiency | PpsaA and PatpA rbcL 3′-UTR | Phototrophic rescue of TN72 (ΔpsbH) strain | The purified hGH has biological activity in vitro | [70] |
| Chimeric antihypertensive peptides (angiotensin-converting enzyme ACE-inhibitory peptides) | Treatment of hypertension | PrbcL rbcL 3′-UTR | aadA1 gene—Spectinomycin | Antihypertensive and ACE-inhibitory effects of the recombinant protein demonstrated in vivo in murine models | [129] |
| Chimeric antihypertensive peptides (angiotensin-converting enzyme ACE-inhibitory peptides) | Treatment of hypertension | PrbcL rbcL 3′-UTR | aadA1 gene—Spectinomycin | Antihypertensive effect of the recombinant protein demonstrated in vivo in murine models | [130] |
| Phosphorylated human osteopontin | Bone regenerative therapy | Not disclosed | Not disclosed | Successful specific folding and PTMs | [128] |
| Full-length IgG1 human monoclonal antibody against anthrax protective antigen 83 (PA83) (heavy and light chains, HC, LC) | Blocker of anthrax toxin | PpsbA rbcL 3′-UTR (HC) PpsbA psbA 3′-UTR (LC) | Mutated 16S-rRNA gene—Spectinomycin | The antibody binds its target antigen, PA83, at levels similar to the same antibody expressed in mammalian cells | [131] |
| Variable domains of camelid heavy chain-only antibodies (VH H) binding and neutralizing botulinum neurotoxin | Antitoxin | PpsbA psbA 3′-UTR | aphA6 gene—Kanamycin | Accumulation of 5% of TSP Prevention of neuron intoxication in vitro Stable in gastric environment | [134] |
| Single-chain fragment variable (scFv) antibody | Proof-of-concept production of bioactive recombinant protein | PpsaA rbcL 3′-UTR | Phototrophic rescue of TN72 (ΔpsbH) strain | Fusion to the Tat export signal peptide-enabled targeting of the recombinant product in the thylakoid lumen | [135] |
| Bacteriophage Cpl-1 and Pal endolysins | Antibacterial effectors against Streptococcus pneumoniae | PpsaA-exon 1 rbcL 3′-UTR | Phototrophic rescue of TN72 (ΔpsbH) strain | Demonstrated antibacterial activity against different serotypes of S. pneumoniae, including clinical isolates | [125] |
| PlyGBS bacterial lysin | Antibacterial effector against Streptococcus | P16S rRNA atpA 5′- and 3′-UTRs | aadA1 gene—Spectinomycin | The effect of light period and intensity on recombinant protein expression was investigated, revealing optimal conditions with mixotrophy under constant illumination at moderately low light (35 μE m−2 s−1) | [107] |
| Single-chain antibody (scFv) targeting the B-cell surface antigen CD22 fused to the enzymatic domain of exotoxin A from Pseudomonas aeruginosa (immunotoxin) | Treatment of B-cell lymphomas | PpsbA psbA 3′-UTR | aphA6 gene—Kanamycin | The expressed proteins specifically bind and reduce the viability of B-cell lymphomas in vitro | [133] |
| Single-chain antibody (scFv) targeting the B-cell surface antigen CD22 fused to the eukaryotic ribosome inactivating protein, gelonin (immunotoxin) | Treatment of B-cell lymphomas | PpsbA psbA 3′-UTR | aphA6 gene—Kanamycin | The expressed proteins specifically bind and reduce the viability of B-cell lymphomas in vitro | [132] |
| Major birch pollen allergen Bet v 1 | Allergen immunotherapy (AIT) for the the treatment of allergic diseases | PpsaA rbcL 3′-UTR | Phototrophic rescue of FUD50 (ΔatpB) strain | The Bet v 1 antigen from algae showed similar binding to human IgE and murine Bet v 1-specific IgG | [137] |
| Foot-and-mouth disease virus VP1 protein fused with cholera toxin B | Antigen Adjuvant | PatpA rbcL 3′-UTR | aadA1 gene—Spectinomycin | The fusion protein displayed GM1-ganglioside-binding affinity and antigenicity | [138] |
| Human glutamic acid decarboxylase (hGAD65) (diabetes-associated autoantigen) | Diagnostic marker/antigen for immunotherapy | PrbcL rbcL 3′-UTR | aadA1 gene—Spectinomycin | Antigenicity of algal derived product verified by ELISA and in vivo assays | [140] |
| D2 fibronectin-binding domain of Staphylococcus aureus fused with the cholera toxin B subunit (CTB) | Oral vaccine | PrbcL rbcL 3′-UTR | aadA1 gene—Spectinomycin | Induction of specific mucosal and systemic immune responses in mice | [139] |
| p210 epitope from apolipoprotein ApoB100 fused to the β subunit of the cholera toxin (CtxB) | Oral vaccine/immunotherapy for atherosclerosis | PatpA rbcL 3′-UTR | aadA1 gene—Spectinomycin | In vivo immunogenic activity of the chimera when orally administered in mice and detection of anti-p210 serum antibodies | [141] |
| Plasmodium falciparum surface protein 25 (Pfs25) and 28 (Pfs28) | Malaria antigen/subunit vaccines | PpsbA psbA 3′-UTRs | aphA6 gene—Kanamycin | The two proteins are immunogenic in mice and Pfs25 antibodies bind in vitro to P. falciparum, exhibiting transmission-blocking activity | [142] |
| C-terminal antigenic domain of the Plasmodium falciparum surface protein Pfs48 and 45 | Malaria antigen/transmission-blocking vaccine | PpsbD psbA-3′-UTR PpsbA psbA-3′-UTR | Phototrophic rescue of psbH—strain | The purified peptides are recognized by specific transmission-blocking antibodies | [112] |
| Plasmodium falciparum surface protein 25 (Pfs25) fused to the β subunit of the cholera toxin (CtxB) | Malaria antigen/transmission-blocking vaccine | PpsbA psbA 3′-UTR | aphA6 gene—Kanamycin | Orally vaccinated mice with freeze-dried algae containing CtxB-Pfs25-elicited CtxB-specific serum IgG antibodies, and both CtxB- and Pfs25-specific secretory IgA antibodies | [143,144] |
| Single-chain fragment antibody molecule (αCD22 scFv) and Plasmodium falciparum surface protein 25 (Pfs25) | Malaria antigen/transmission-blocking vaccine | PpsbA psbA 3′-UTR | aphA6 gene—Kanamycin | Optimization of light intensity (300 μmol m−2 s−1) resulted in six-fold increase in protein accumulation | [113] |
| Plasmodium falciparum PfCelTOS antigen (cell traversal protein for ookinetes and sporozoites) alone and fused to human interleukin-2 (IL-2) | Malaria antigen/transmission-blocking vaccine fused to adjuvant | PatpA rbcL 3′-UTR | Phototrophic rescue of TN72 (ΔpsbH) strain | Protein accumulation is promoted by mixotrophic cultivation in low light | [145,146] |
| Biopharmaceuticals and enzymes for aquaculture, animal health, and pest control | |||||
| vapA and acrV proteins from the fish pathogen Aeromonas salmonicida | Antigen/immunization | Various combinations of promoters, 5′- and 3′-UTRs | Phototrophic rescue of FUD50 (ΔatpB) strain aadA1 gene—Spectinomycin | Strongest expression with the PpsaA-exon1/5′-UTR element | [95] |
| VP28 protein of the white spot syndrome virus | Oral vaccine | PpsbA psbA 3′-UTR | aadA1 gene—Spectinomycin | Recombinant product accumulated to 21% of TCP | [58] |
| Classical swine fever virus (CSFV) structural protein | Antigen/immunization | PatpA rbcL 3′-UTR | aadA1 gene—Spectinomycin | Accumulation of 1.5–1% of TSP, antigenicity verified by ELISA | [151] |
| Escherichia coli AppA phytase enzyme | Feed additive for poultry | PatpA rbcL 3′-UTR | Mutated 16S-rRNA gene—Spectinomycin | Fecal phytate excretion is reduced following feeding with whole-cell algal lysate | [152] |
| Cry (1A, 4A, 4B and 11A) cytotoxic proteins of Bacillus thuringiensis subsp. israelensis (Bti) | Mosquito control | PpsbD psbA 3′-UTR | Mutated 16S-rRNA gene—Spectinomycin | Live cell bioassays demonstrated toxicity of the cry transformants to larvae of Aedes aegypti and Culex quinquefasciatus | [168,169] |
| VP28 protein of the white spot syndrome virus (WSSV) | Oral delivery system to control WSSV disease in shrimp | PatpA rbcL 3′-UTR | Phototrophic rescue of TN72 (ΔpsbH) strain | Feeding of algal biomass exressing the VP28 antigen improved shrimp survival upon infection with WSSV | [149] |
| Enzymes with industrial applications | |||||
| Alcohol dehydrogenase (ADH1) from Saccharomyces cerevisiae | Ethanol production | PrbcL rbcL 3′-UTR | Phototrophic rescue of rbcL (CC2653) mutant | Algal cultivation in low oxygen partial pressure or anoxia promoted ADH1 accumulation and ethanol production | [65] |
| β-carotene hydroxylase (crtR-B) from Haematococcus pluvialis (lacustris) | Metabolic engineering/heterologous synthesis of astaxanthin | PatpA rbcL 3′-UTR | aadA1 geneSpectinomycin | Total carotenoid content is increased in the crtR-B transformants following high light treatment compared to wild type cells | [154] |
| Plant cytochrome P450 (CYP79A1) | Metabolic engineering/heterologous synthesis of diterpenoids | PatpA rbcL 3′-UTR | Phototrophic rescue of TN72 (ΔpsbH) strain | The enzyme is targeted to the organelle membrane via its endogenous N-terminal region, and converts tyrosine to p-hydroxyphenylacetaldoxime | [159] |
| Bifunctional diterpene synthase (cis-abienol synthase, TPS4) | Metabolic engineering/heterologous synthesis of diterpenoids | PatpA rbcL 3′-UTR | Phototrophic rescue of TN72 (ΔpsbH) strain | The expression of this enzyme is compatible with pilot-scale algal cultivation [160] | [158] |
| Glycohydrolase family 5 endoglucanase from Paenibacillus sp. KCTC8848P (CelK1) | Hydrolytic enzyme/saccharification of lignocellulosic biomass | PpsaA rbcL 3′-UTR | Phototrophic rescue of FUD50 (ΔatpB) strain | [162] | |
| Cel6A endoglucanase from Thermobifida fusca | Hydrolytic enzyme/saccharification of lignocellulosic biomass | P16S rRNA atpA 5′-UTR rbcL 3′-UTR | aadA1 gene—Spectinomycin | A fusion to the downstream box (DB) of the TetC (tetracycline cyclase) gene improved protein accumulation | [163] |
| CelB endoglucanase from Thermotoga neapolitana | Hyperthermophilic hydrolytic enzyme/saccharification of lignocellulosic biomass | PpsaA rbcL 3′-UTR | aadA1 gene—Spectinomycin | The cellulolytic blend enabled the conversion of alkaline-treated lignocellulose into glucose Hydrolysates boosted the biogas production by methanogenic bacteria | [164] |
| Cellobiohydrolase portion of the CelB cellulosome (C-CBH) from Caldicellulosiruptor saccharolyticus | |||||
| β-glucosidase from Pyrococcus furiosus | |||||
| Xylanase from Thermotoga neapolitana | |||||
| CelB endoglucanase from Thermotoga neapolitana | Hyperthermophilic hydrolytic enzyme/saccharification of lignocellulosic biomass | PpsaA rbcL 3′-UTR | Phototrophic rescue of FUD50 (ΔatpB) strain | Expression of the hydrolytic enzyme is coupled to the PTXD growth selector, enabling selective growth in non-sterile, phosphite-fertilized medium | [79] |
5. Multigenic Engineering
6. Inducible Expression Systems
7. Beyond Model Species
| Species | Class, Order, and Family | Physiological Characteristics | Transformation Method and Selection System | Plastome Integration Site(s) | Expressed Product and Cis-Regulatory Elements | Reference |
|---|---|---|---|---|---|---|
| Green algae (Chlorophyta) | ||||||
| Tetraselmis subcordiformis | Chlorodendrophyceace, Chlorodendrales, Chlorodendraceae | Halophilic | Biolistics Bar gene (phosphinothricin N-acetyltransferase)—Bialaphos | Silent site between rrn16S/tRNA-I and tRNA-A/rrn23S | Enhanced green fluorescent protein (eGFP) Endogenous PatpA C. reinhardtii rbcL 3′-UTR | [200] |
| Tetraselmis subcordiformis | Chlorodendrophyceace, Chlorodendrales, Chlorodendraceae | Halophilic | Biolistics Bar gene—Bialaphos | Silent site between rrn16S/tRNA-I and tRNA-A/rrn23S | Enhanced green fluorescent protein (eGFP) Combination of endogenous elements from rrn16S, rbcL, psbA, and psbC Prrn-psbA-5′UTR–psbA 3′-UTR produced highest expression | [201] |
| Dunaliella salina | Chlorophyceae, Chlamydomonadales, Dunaliellaceae | Halophilic | Biolistics hptll gene (hygromycin phosphotransferase)—Hygromycin | Silent site between rrn16S/tRNA-I and tRNA-A/rrn23S | Enhanced green fluorescent protein (eGFP) Oryza sativa plastid ribosomal RNA operon promoter (Prrn) and psbA 5′-UTR | [202] |
| Dunaliella tertiolecta | Chlorophyceae, Chlamydomonadales, Dunaliellaceae | Halophilic | Biolistics ereB gene (erythromycin esterase)—Erythromycin | Silent site between psbB/psbH | Hydrolytic enzymes: xylanase, α-galactosidase, phytase, phosphate anhydrolase, and β-mannanase Endogenous PpsbD Endogenous psbA 3′-UTR | [161] |
| Desmodesmus armatus | Chlorophyceae, Sphaeropleales, Scenedesmaceae | Resistant to high light and cold temperatures | Biolistics Mutated PsbA gene (S264K) conferring atrazine resistance—Atrazine | Endogenous psbA locus | - | [203] |
| Haematococcus pluvialis (lacustris) | Chlorophyceae, Chlamydomonadales, Haematococcaceae | Natural producer of high-value compound astaxanthin | Biolistics Aad1 gene (aminoglycoside adenyltransferase)—Spectinomycin | Silent site between rrn16S/rrn23S | Endogenous PrbcL Endogenous rbcL 3′-UTR | [204] |
| Haematococcus pluvialis (lacustris) | Chlorophyceae, Chlamydomonadales, Haematococcaceae | Natural producer of high-value compound astaxanthin | Biolistics Aad1 gene–Spectinomycin | Silent site between rrn16S/rrn23S | Phytoene desaturase (pds) to enhance endogenous astaxanthin accumulation Endogenous PpsbA Endogenous rbcL 3′-UTR | [205] |
| Haematococcus pluvialis (lacustris) | Chlorophyceae, Chlamydomonadales, Haematococcaceae | Natural producer of high-value compound astaxanthin | Biolistics Bar gene–Bialaphos | Silent site between rrn16S/tRNA-I and tRNA-A/rrn23S | Antimicrobial peptide piscidin-4 (ant1) Endogenous PrbcL Endogenous psbA 3′-UTR | [206] |
| Chlorella vulgaris | Trebouxiophyceae, Chlorellales, Chlorellaceace | Thermo- and high-light-tolerant and high biomass producer | Biolistics Aad1 gene—Spectinomycin | Silent site between rrn16S/tRNA-I and tRNA-A/rrn23S | Antimicrobial peptide NZ2114 and piscidin-4 (ant1, ant2) Endogenous P16S Endogenous rbcL 3′-UTR (Aad1) Endogenous PrbcL Endogenous psbA 3′-UTR (ant1, ant2) | [211] |
| Parachlorella kessleri | Trebouxiophyceae, Chlorellales, Chlorellaceace | Thermo- and high-light-tolerant and high biomass producer | Biolistics Aad1 gene—Spectinomycin | Silent site between rrn16S/tRNA-I and tRNA-A/rrn23S | Endogenous PpsbA Endogenous psbA 3′-UTR | [213] |
| Picochlorum renovo | Trebouxiophyceae, Chlorellales, Incertae sedis | Halophilic, thermotolerant, and high biomass producer | Biolistics ereB gene—Erythromycin | Silent site between rrn16S/tRNA-I and tRNA-A/rrn23S | Super folder GFP (sfGFP) Endogenous P16S Endogenous 16S 3′-UTR | [173] |
| Picochlorum renovo and celeri | Trebouxiophyceae, Chlorellales, Incertae sedis | Halophilic, thermotolerant, and high biomass producer | Biolistics Chloroplast optimized ptxD isoform [77]—Phosphite | Silent site between rrn16S/tRNA-I and tRNA-A/rrn23S | PTXD Endogenous P16S Synthetic terminator | [78] |
| Stramenopiles | ||||||
| Nannochloropsis oceanica | Eustigmatophyceae, Eustigmatales, Monodopsidaceae | Halophilic, high-lipid-accumulating | Electroporation Sh ble gene—Zeocyn | Endogenous chlL locus | Green Fluorescent protein (GFP) Endogenous PrbcL Endogenous psbA 3′-UTR | [48] |
| Nannochloropsis gaditana | Eustigmatophyceae, Eustigmatales, Monodopsidaceae | Halophilic, high-lipid-accumulating | Biolistics bar gene—Bialaphos | Silent site between rrn16S/tRNA-I and tRNA-A/rrn23S | Antimicrobial peptides ant1, ant2 Endogenous PpsbA Endogenous rbcL 3′-UTR (bar) Endogenous PrbcL Endogenous psbA 3′-UTR (ant1, ant2) | [214] |
| Phaeodactylum tricornutum | Bacillariophyae, Bacillariales, Phaeodactilaceae | Halophilic, high-lipid-accumulating | Electroporation cat gene (chloramphenicol acetyltransferase)—chloramphenicol | Silent site between rrn16S/tRNA-I and tRNA-A/rrn23S | Enhanced green fluorescent protein (eGFP) Endogenous PrbcL Endogenous rbcS 3′-UTR | [215] |
8. What Next?
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Jackson, D.A.; Symons, R.H.; Berg, P. Biochemical Method for Inserting New Genetic Information into DNA of Simian Virus 40: Circular SV40 DNA Molecules Containing Lambda Phage Genes and the Galactose Operon of Escherichia coli. Proc. Natl. Acad. Sci. USA 1972, 69, 2904–2909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.N.; Chang, A.C.; Boyer, H.W.; Helling, R.B. Construction of biologically functional bacterial plasmids in vitro. Proc. Natl. Acad. Sci. USA 1973, 70, 3240–3244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itakura, K.; Hirose, T.; Crea, R.; Riggs, A.D.; Heyneker, H.L.; Bolivar, F.; Boyer, H.W. Expression in Escherichia coli of a Chemically Synthesized Gene for the Hormone Somatostatin. Science 1977, 198, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Sproles, A.E.; Fields, F.J.; Smalley, T.N.; Le, C.H.; Badary, A.; Mayfield, S.P. Recent advancements in the genetic engineering of microalgae. Algal Res. 2021, 53, 102158. [Google Scholar] [CrossRef]
- Gutiérrez, S.; Lauersen, K. Gene Delivery Technologies with Applications in Microalgal Genetic Engineering. Biology 2021, 10, 265. [Google Scholar] [CrossRef]
- Nickelsen, J.; Kück, U. The unicellular green alga Chlamydomonas reinhardtii as an experimental system to study chloroplast RNA metabolism. Naturwissenschaften 2000, 87, 97–107. [Google Scholar] [CrossRef]
- Rochaix, J.-D. Functional Analysis of Plastid Genes through Chloroplast Reverse Genetics in Chlamydomonas. In Photosynthesis in Algae; Larkum, A.W.D., Douglas, S.E., Raven, J.A., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 83–94. [Google Scholar]
- Keeling, P.J. The endosymbiotic origin, diversification and fate of plastids. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2010, 365, 729–748. [Google Scholar] [CrossRef] [Green Version]
- Franklin, S.; Ngo, B.; Efuet, E.; Mayfield, S.P. Development of a GFP reporter gene for Chlamydomonas reinhardtii chloroplast. Plant J. 2002, 30, 733–744. [Google Scholar] [CrossRef] [Green Version]
- Gallaher, S.D.; Fitz-Gibbon, S.T.; Strenkert, D.; Purvine, S.O.; Pellegrini, M.; Merchant, S.S. High-throughput sequencing of the chloroplast and mitochondrion of Chlamydomonas reinhardtii to generate improved de novo assemblies, analyze expression patterns and transcript speciation, and evaluate diversity among laboratory strains and wild isolates. Plant J. 2018, 93, 545–565. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, K.; Beligni, M.V.; Prieto, S.; Haynes, P.A.; McDonald, W.H.; Yates, J.R.; Mayfield, S.P. Proteomic Characterization of the Chlamydomonas reinhardtii Chloroplast Ribosome: Identification of proteins unique to the 70 s ribosome. J. Biol. Chem. 2003, 278, 33774–33785. [Google Scholar] [CrossRef] [Green Version]
- Harris, E.H.; Burkhart, B.D.; Gillham, N.W.; Boynton, J.E. Antibiotic resistance mutations in the chloroplast 16S and 23S rRNA genes of Chlamydomonas reinhardtii: Correlation of genetic and physical maps of the chloroplast genome. Genetics 1989, 123, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Newman, S.M.; Boynton, J.E.; Gillham, N.W.; Randolph-Anderson, B.L.; Johnson, A.M.; Harris, E.H. Transformation of chloroplast ribosomal RNA genes in Chlamydomonas: Molecular and genetic characterization of integration events. Genetics 1990, 126, 875–888. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Stoebe, B.; Goremykin, V.; Hansmann, S.; Hasegawa, M.; Kowallik, K.V.; Hapsmann, S. Gene transfer to the nucleus and the evolution of chloroplasts. Nature 1998, 393, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Fong, S.E.; Surzycki, S.J. Chloroplast RNA polymerase genes of Chlamydomonas reinhardtii exhibit an unusual structure and arrangement. Curr. Genet. 1992, 21, 485–497. [Google Scholar] [CrossRef]
- Terashima, M.; Specht, M.; Hippler, M. The chloroplast proteome: A survey from the Chlamydomonas reinhardtii perspective with a focus on distinctive features. Curr. Genet. 2011, 57, 151–168. [Google Scholar] [CrossRef]
- Shi, L.-X.; Theg, S.M. The chloroplast protein import system: From algae to trees. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2013, 1833, 314–331. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Bakhtiari, S.; Valente-Paterno, M.; Wu, Y.; Law, C.; Dai, D.; Dhaliwal, J.; Bui, K.H.; Zerges, W. Chloroplast-localized translation for protein targeting in Chlamydomonas reinhardtii. bioRxiv 2021. [Google Scholar] [CrossRef]
- Maul, J.; Lilly, J.W.; Cui, L.; DePamphilis, C.W.; Miller, W.; Harris, E.H.; Stern, D.B. The Chlamydomonas reinhardtii Plastid Chromosome: Islands of Genes in a Sea of Repeats. Plant Cell 2002, 14, 2659–2679. [Google Scholar] [CrossRef]
- Karcher, D.; Köster, D.; Schadach, A.; Klevesath, A.; Bock, R. The Chlamydomonas Chloroplast HLP Protein Is Required for Nucleoid Organization and Genome Maintenance. Mol. Plant 2009, 2, 1223–1232. [Google Scholar] [CrossRef]
- Adams, G. Chloroplast gene transmission in Chlamydomonas reinhardtii: A random choice model. Plasmid 1978, 1, 522–535. [Google Scholar] [CrossRef]
- Simpson, C.L.; Stern, D.B. The Treasure Trove of Algal Chloroplast Genomes. Surprises in Architecture and Gene Content, and Their Functional Implications. Plant Physiol. 2002, 129, 957–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turmel, M.; Lemieux, C. Chapter Six—Evolution of the Plastid Genome in Green Algae. In Advances in Botanical Research; Chaw, S.-M., Jansen, R.K., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 157–193. [Google Scholar]
- Jiao, H.S.; Hicks, A.; Simpson, C.; Stern, D.B. Short dispersed repeats in the Chlamydomonas chloroplast genome are collocated with sites for mRNA 3’ end formation. Curr. Genet. 2004, 45, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Odom, O.W.; Baek, K.-H.; Dani, R.N.; Herrin, D.L. Chlamydomonas chloroplasts can use short dispersed repeats and multiple pathways to repair a double-strand break in the genome. Plant J. 2008, 53, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Nakazato, E.; Fukuzawa, H.; Tabata, S.; Takahashi, H.; Tanaka, K. Identification and Expression Analysis of cDNA Encoding a Chloroplast Recombination Protein REC1, the Chloroplast RecA Homologue in Chlamydomonas reinhardtii. Biosci. Biotechnol. Biochem. 2003, 67, 2608–2613. [Google Scholar] [CrossRef] [Green Version]
- Gumpel, N.J.; Rochaix, J.-D.; Purton, S. Studies on homologous recombination in the green alga Chlamydomonas reinhardtii. Curr. Genet. 1994, 26, 438–442. [Google Scholar] [CrossRef]
- Durrenberger, F. Double strand break-induced recombination in Chlamydomonas reinhardtii chloroplasts. Nucleic Acids Res. 1996, 24, 3323–3331. [Google Scholar] [CrossRef] [Green Version]
- Bohne, A.-V.; Irihimovitch, V.; Weihe, A.; Stern, D.B. Chlamydomonas reinhardtii encodes a single sigma70-like factor which likely functions in chloroplast transcription. Curr. Genet. 2006, 49, 333–340. [Google Scholar] [CrossRef]
- Eberhard, S.; Drapier, D.; Wollman, F.-A. Searching limiting steps in the expression of chloroplast-encoded proteins: Relations between gene copy number, transcription, transcript abundance and translation rate in the chloroplast of Chlamydomonas reinhardtii. Plant J. 2002, 31, 149–160. [Google Scholar] [CrossRef]
- Minai, L.; Wostrikoff, K.; Wollman, F.-A.; Choquet, Y. Chloroplast Biogenesis of Photosystem II Cores Involves a Series of Assembly-Controlled Steps That Regulate Translation. Plant Cell 2006, 18, 159–175. [Google Scholar] [CrossRef]
- Stern, D.B.; Goldschmidt-Clermont, M.; Hanson, M.R. Chloroplast RNA Metabolism. Annu. Rev. Plant Biol. 2010, 61, 125–155. [Google Scholar] [CrossRef]
- Macedo-Osorio, K.S.; Martínez-Antonio, A.; Badillo-Corona, J.A. Pas de Trois: An Overview of Penta-, Tetra-, and Octo-Tricopeptide Repeat Proteins From Chlamydomonas reinhardtii and Their Role in Chloroplast Gene Expression. Front. Plant Sci. 2021, 12, 775366. [Google Scholar] [CrossRef] [PubMed]
- Gutmann, B.; Royan, S.; Schallenberg-Rüdinger, M.; Lenz, H.; Castleden, I.R.; McDowell, R.; Vacher, M.A.; Tonti-Filippini, J.; Bond, C.S.; Knoop, V.; et al. The Expansion and Diversification of Pentatricopeptide Repeat RNA-Editing Factors in Plants. Mol. Plant 2020, 13, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Cahoon, A.B.; Nauss, J.A.; Stanley, C.D.; Qureshi, A. Deep Transcriptome Sequencing of Two Green Algae, Chara vulgaris and Chlamydomonas reinhardtii, Provides No Evidence of Organellar RNA Editing. Genes 2017, 8, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royan, S.; Gutmann, B.; des Francs-Small, C.C.; Honkanen, S.; Schmidberger, J.; Soet, A.; Sun, Y.; Sanglard, L.V.P.; Bond, C.; Small, I. A synthetic RNA editing factor edits its target site in chloroplasts and bacteria. Commun. Biol. 2021, 4, 545. [Google Scholar] [CrossRef] [PubMed]
- Holloway, S.P.; Deshpande, N.N.; Herrin, D.L. The catalytic group-I introns of the psbA gene of Chlamydomonas reinhardtii: Core structures, ORFs and evolutionary implications. Curr. Genet. 1999, 36, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Kück, U.; Schmitt, O. The Chloroplast Trans-Splicing RNA–Protein Supercomplex from the Green Alga Chlamydomonas reinhardtii. Cells 2021, 10, 290. [Google Scholar] [CrossRef] [PubMed]
- Cavaiuolo, M.; Kuras, R.; Wollman, F.; Choquet, Y.; Vallon, O. Small RNA profiling in Chlamydomonas: Insights into chloroplast RNA metabolism. Nucleic Acids Res. 2017, 45, 10783–10799. [Google Scholar] [CrossRef] [Green Version]
- Schroda, M. The Chlamydomonas genome reveals its secrets: Chaperone genes and the potential roles of their gene products in the chloroplast. Photosynth. Res. 2004, 82, 221–240. [Google Scholar] [CrossRef]
- Kim, J.; Mayfield, S.P. Protein Disulfide Isomerase as a Regulator of Chloroplast Translational Activation. Science 1997, 278, 1954–1957. [Google Scholar] [CrossRef]
- Vallon, O. Chlamydomonas Immunophilins and Parvulins: Survey and Critical Assessment of Gene Models. Eukaryot. Cell 2005, 4, 230–241. [Google Scholar] [CrossRef] [Green Version]
- Boynton, J.E.; Gillham, N.W.; Harris, E.H.; Hosler, J.P.; Johnson, A.M.; Jones, A.R.; Randolph-Anderson, B.L.; Robertson, D.; Klein, T.M.; Shark, K.B.; et al. Chloroplast Transformation in Chlamydomonas with High Velocity Microprojectiles. Science 1988, 240, 1534–1538. [Google Scholar] [CrossRef]
- Blowers, A.D.; Bogorad, L.; Shark, K.B.; Sanford, J.C. Studies on Chlamydomonas Chloroplast Transformation: Foreign DNA Can Be Stably Maintained in the Chromosome. Plant Cell 1989, 1, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán-Zapata, D.; Macedo-Osorio, K.S.; Almaraz-Delgado, A.L.; Durán-Figueroa, N.; Badillo-Corona, J.A. Production of Recombinant Proteins in the Chloroplast of the Green Alga Chlamydomonas reinhardtii. Methods Mol. Biol. 2016, 1385, 69–85. [Google Scholar] [PubMed]
- Economou, C.; Wannathong, T.; Szaub, J.; Purton, S. A Simple, Low-Cost Method for Chloroplast Transformation of the Green Alga Chlamydomonas reinhardtii. In Chloroplast Biotechnology: Methods and Protocols; Maliga, P., Ed.; Humana Press: Totowa, NJ, USA, 2014; pp. 401–411. [Google Scholar]
- Larrea-Alvarez, M.; Young, R.; Purton, S. A Simple Technology for Generating Marker-Free Chloroplast Transformants of the Green Alga Chlamydomonas reinhardtii. Methods Mol. Biol. 2021, 2317, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Gan, Q.; Jiang, J.; Han, X.; Wang, S.; Lu, Y. Engineering the Chloroplast Genome of Oleaginous Marine Microalga Nannochloropsis oceanica. Front. Plant Sci. 2018, 9, 439. [Google Scholar] [CrossRef] [Green Version]
- Weiner, I.; Feldman, Y.; Shahar, N.; Yacoby, I.; Tuller, T. CSO—A sequence optimization software for engineering chloroplast expression in Chlamydomonas reinhardtii. Algal Res. 2020, 46, 101788. [Google Scholar] [CrossRef]
- Dauvillee, D.; Hilbig, L.; Preiss, S.; Johanningmeier, U. Minimal Extent of Sequence Homology Required for Homologous Recombination at the psbA Locus in Chlamydomonas reinhardtii Chloroplasts using PCR-generated DNA Fragments. Photosynth. Res. 2004, 79, 219–224. [Google Scholar] [CrossRef]
- Neupert, J.; Gallaher, S.D.; Lu, Y.; Strenkert, D.; Segal, N.A.; Barahimipour, R.; Fitz-Gibbon, S.T.; Schroda, M.; Merchant, S.S.; Bock, R. An epigenetic gene silencing pathway selectively acting on transgenic DNA in the green alga Chlamydomonas. Nat. Commun. 2020, 11, 6269. [Google Scholar] [CrossRef]
- Neupert, J.; Karcher, D.; Bock, R. Generation of Chlamydomonas strains that efficiently express nuclear transgenes. Plant J. 2009, 57, 1140–1150. [Google Scholar] [CrossRef]
- Schroda, M. Good News for Nuclear Transgene Expression in Chlamydomonas. Cells 2019, 8, 1534. [Google Scholar] [CrossRef] [Green Version]
- Shahar, N.; Landman, S.; Weiner, I.; Elman, T.; Dafni, E.; Feldman, Y.; Tuller, T.; Yacoby, I. The Integration of Multiple Nuclear-Encoded Transgenes in the Green Alga Chlamydomonas reinhardtii Results in Higher Transcription Levels. Front. Plant Sci. 2020, 10, 1784. [Google Scholar] [CrossRef] [PubMed]
- Angstenberger, M.; de Signori, F.; Vecchi, V.; Dall’Osto, L.; Bassi, R. Cell Synchronization Enhances Nuclear Transformation and Genome Editing via Cas9 Enabling Homologous Recombination in Chlamydomonas reinhardtii. ACS Synth. Biol. 2020, 9, 2840–2850. [Google Scholar] [CrossRef] [PubMed]
- Baier, T.; Jacobebbinghaus, N.; Einhaus, A.; Lauersen, K.J.; Kruse, O. Introns mediate post-transcriptional enhancement of nuclear gene expression in the green microalga Chlamydomonas reinhardtii. PLoS Genet. 2020, 16, e1008944. [Google Scholar] [CrossRef] [PubMed]
- Kurniasih, S.D.; Yamasaki, T.; Kong, F.; Okada, S.; Widyaningrum, D.; Ohama, T. UV-mediated Chlamydomonas mutants with enhanced nuclear transgene expression by disruption of DNA methylation-dependent and independent silencing systems. Plant Mol. Biol. 2016, 92, 629–641. [Google Scholar] [CrossRef]
- Surzycki, R.; Greenham, K.; Kitayama, K.; Dibal, F.; Wagner, R.; Rochaix, J.D.; Ajam, T.; Surzycki, S. Factors effecting expression of vaccines in microalgae. Biologicals 2009, 37, 133–138. [Google Scholar] [CrossRef]
- Goldschmidt-Clermont, M. Transgenic expression of aminoglycoside adenine transferase in the chloroplast: A selectable marker for site-directed transformation of chlamydomonas. Nucleic Acids Res. 1991, 19, 4083–4089. [Google Scholar] [CrossRef] [Green Version]
- Bateman, J.M.; Purton, S. Tools for chloroplast transformation in Chlamydomonas: Expression vectors and a new dominant selectable marker. Mol. Gen. Genet. 2000, 263, 404–410. [Google Scholar] [CrossRef]
- Lee, J.W. Protocol measuring horizontal gene transfer from algae to non-photosynthetic organisms. MethodsX 2019, 6, 1564–1574. [Google Scholar] [CrossRef]
- Fischer, N.; Stampacchia, O.; Redding, K.; Rochaix, J.D. Selectable marker recycling in the chloroplast. Mol. Gen. Genet. 1996, 251, 373–380. [Google Scholar] [CrossRef]
- Woessner, J.P.; Masson, A.; Harris, E.H.; Bennoun, P.; Gillham, N.W.; Boynton, J.E. Molecular and genetic analysis of the chloroplast ATPase ofChlamydomonas. Plant Mol. Biol. 1985, 4, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Bertalan, I.; Munder, M.; Weiß, C.; Kopf, J.; Fischer, D.; Johanningmeier, U. A rapid, modular and marker-free chloroplast expression system for the green alga Chlamydomonas reinhardtii. J. Biotechnol. 2015, 195, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-C.; Melis, A. Marker-free genetic engineering of the chloroplast in the green microalga Chlamydomonas reinhardtii. Plant Biotechnol. J. 2013, 11, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Day, A.; Dowson-Day, M.; Shen, G.-F.; Dixon, R. The Klebsiella pneumoniae nitrogenase Fe protein gene (nifH) functionally substitutes for the chlL gene in Chlamydomonas reinhardtii. Biochem. Biophys. Res. Commun. 2005, 329, 966–975. [Google Scholar] [CrossRef] [PubMed]
- Kindle, K.L.; Richards, K.L.; Stern, D.B. Engineering the chloroplast genome: Techniques and capabilities for chloroplast transformation in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 1991, 88, 1721–1725. [Google Scholar] [CrossRef] [Green Version]
- Balczun, C.; Bunse, A.; Hahn, D.; Bennoun, P.; Nickelsen, J.; Kück, U.; Nickelsen, J. Two adjacent nuclear genes are required for functional complementation of a chloroplast trans-splicing mutant from Chlamydomonas reinhardtii. Plant J. 2005, 43, 636–648. [Google Scholar] [CrossRef]
- O’Connor, H.E.; Ruffle, S.V.; Cain, A.J.; Deak, Z.; Vass, I.; Nugent, J.H.; Purton, S. The 9-kDa phosphoprotein of photosystem: II. Generation and characterisation of Chlamydomonas mutants lacking PSII-H and a site-directed mutant lacking the phosphorylation site. Biochim. Biophys. Acta 1998, 1364, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Wannathong, T.; Waterhouse, J.C.; Young, R.E.B.; Economou, C.K.; Purton, S. New tools for chloroplast genetic engineering allow the synthesis of human growth hormone in the green alga Chlamydomonas reinhardtii. Appl. Microbiol. Biotechnol. 2016, 100, 5467–5477. [Google Scholar] [CrossRef] [Green Version]
- Costas, A.M.G.; White, A.K.; Metcalf, W.W. Purification and Characterization of a Novel Phosphorus-oxidizing Enzyme from Pseudomonas stutzeri WM88. J. Biol. Chem. 2001, 276, 17429–17436. [Google Scholar] [CrossRef] [Green Version]
- Loera-Quezada, M.M.; González, M.A.L.; Velazquez, G.; Sanchez-Calderón, L.; Nascimento, M.D.; López-Arredondo, D.; Herrera-Estrella, L. A novel genetic engineering platform for the effective management of biological contaminants for the production of microalgae. Plant Biotechnol. J. 2016, 14, 2066–2076. [Google Scholar] [CrossRef] [Green Version]
- Loera-Quezada, M.M.; González, M.A.L.; López-Arredondo, D.; Herrera-Estrella, L. Phosphite cannot be used as a phosphorus source but is non-toxic for microalgae. Plant Sci. 2015, 231, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Sandoval, J.; Macedo-Osorio, K.S.; Durán-Figueroa, N.V.; Garibay-Orijel, C.; Badillo-Corona, J.A. Chloroplast engineering of Chlamydomonas reinhardtii to use phosphite as phosphorus source. Algal Res. 2018, 33, 291–297. [Google Scholar] [CrossRef]
- Sandoval, J.; Jiménez-Clemente, L.A.; Macedo-Osorio, K.S.; Oliver-Salvador, M.C.; Fernández-Linares, L.C.; Durán-Figueroa, N.V.; Badillo-Corona, J.A. Use of the ptxD gene as a portable selectable marker for chloroplast transformation in Chlamydomonas reinhardtii. Mol. Biotechnol. 2019, 61, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Changko, S.; Rajakumar, P.D.; Young, R.E.B.; Purton, S. The phosphite oxidoreductase gene, ptxD as a bio-contained chloroplast marker and crop-protection tool for algal biotechnology using Chlamydomonas. Appl. Microbiol. Biotechnol. 2020, 104, 675–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutolo, E.; Tosoni, M.; Barera, S.; Herrera-Estrella, L.; Dall’Osto, L.; Bassi, R. A Phosphite Dehydrogenase Variant with Promiscuous Access to Nicotinamide Cofactor Pools Sustains Fast Phosphite-Dependent Growth of Transplastomic Chlamydomonas reinhardtii. Plants 2020, 9, 473. [Google Scholar] [CrossRef] [Green Version]
- Dahlin, L.R.; Guarnieri, M.T. Heterologous expression of phosphite dehydrogenase in the chloroplast or nucleus enables phosphite utilization and genetic selection in Picochlorum spp. Algal Res. 2022, 62, 102604. [Google Scholar] [CrossRef]
- Cutolo, E.; Tosoni, M.; Barera, S.; Herrera-Estrella, L.; Dall’Osto, L.; Bassi, R. A chimeric hydrolase-PTXD transgene enables chloroplast-based heterologous protein expression and non-sterile cultivation of Chlamydomonas reinhardtii. Algal Res. 2021, 59, 102429. [Google Scholar] [CrossRef]
- Specht, E.A.; Mayfield, S.P. Synthetic Oligonucleotide Libraries Reveal Novel Regulatory Elements in Chlamydomonas Chloroplast mRNAs. ACS Synth. Biol. 2013, 2, 34–46. [Google Scholar] [CrossRef]
- Blowers, A.D.; Klein, U.; Ellmore, G.S.; Bogorad, L. Functional in vivo analyses of the 3’ flanking sequences of the Chlamydomonas chloroplast rbcL and psaB genes. Mol. Gen. Genet. 1993, 238, 339–349. [Google Scholar] [CrossRef]
- Klinkert, B.; Elles, I.; Nickelsen, J. Translation of chloroplast psbD mRNA in Chlamydomonas is controlled by a secondary RNA structure blocking the AUG start codon. Nucleic Acids Res. 2006, 34, 386–394. [Google Scholar] [CrossRef] [Green Version]
- Rahim, M.M.A.; Vigneault, F.; Zerges, W. The RNA Structure of cis-acting Translational Elements of the Chloroplast psbC mRNA in Chlamydomonas reinhardtii. Front. Plant Sci. 2016, 7, 828. [Google Scholar] [CrossRef] [Green Version]
- Rott, R.; Liveanu, V.; Drager, R.G.; Stern, D.B.; Schuster, G. The sequence and structure of the 3′-untranslated regions of chloroplast transcripts are important determinants of mRNA accumulation and stability. Plant Mol. Biol. 1998, 36, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Suay, L.; Salvador, M.L.; Abesha, E.; Klein, U. Specific roles of 5′ RNA secondary structures in stabilizing transcripts in chloroplasts. Nucleic Acids Res. 2005, 33, 4754–4761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldschmidt-Clermont, M.; Rahire, M.; Rochaix, J.-D. Redundant cis-acting determinants of 3′ processing and RNA stability in the chloroplast rbcL mRNA of Chlamydomonas. Plant J. 2008, 53, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Boutanaev, A.; Zucchi, P.; Bogorad, L. Gene elements that affect the longevity of rbcL sequence-containing transcripts in Chlamydomonas reinhardtii chloroplasts. Proc. Natl. Acad. Sci. USA 2001, 98, 2289–2294. [Google Scholar] [CrossRef] [Green Version]
- Salvador, M.L.; Suay, L.; Anthonisen, I.L.; Klein, U. Changes in the 5’-untranslated region of the rbcL gene accelerate transcript degradation more than 50-fold in the chloroplast of Chlamydomonas reinhardtii. Curr. Genet. 2004, 45, 176–182. [Google Scholar] [CrossRef]
- Mayfield, S.P.; Cohen, A.; Danon, A.; Yohn, C. Translation of the psbA mRNA of Chlamydomonas reinhardtii requires a structured RNA element contained within the 5’ untranslated region. J. Cell Biol. 1994, 127, 1537–1545. [Google Scholar] [CrossRef] [Green Version]
- Barnes, D.; Franklin, S.; Schultz, J.; Henry, R.; Brown, E.; Coragliotti, A.; Mayfield, S.P. Contribution of 5′- and 3′-untranslated regions of plastid mRNAs to the expression of Chlamydomonas reinhardtii chloroplast genes. Mol. Genet. Genom. 2005, 274, 625–636. [Google Scholar] [CrossRef]
- Baecker, J.J.; Sneddon, J.C.; Hollingsworth, M.J. Efficient translation in chloroplasts requires element(s) upstream of the putative ribosome binding site from atpI. Am. J. Bot. 2009, 96, 627–636. [Google Scholar] [CrossRef] [Green Version]
- Coragliotti, A.T.; Beligni, M.V.; Franklin, S.E.; Mayfield, S.P. Molecular Factors Affecting the Accumulation of Recombinant Proteins in the Chlamydomonas reinhardtii Chloroplast. Mol. Biotechnol. 2011, 48, 60–75. [Google Scholar] [CrossRef] [Green Version]
- Blowers, A.D.; Ellmore, G.S.; Klein, U.; Bogorad, L. Transcriptional Analysis of Endogenous and Foreign Genes in Chloroplast Transformants of Chlamydomonas. Plant Cell 1990, 2, 1059. [Google Scholar] [CrossRef] [Green Version]
- Ishikura, K.; Takaoka, Y.; Kato, K.; Sekine, M.; Yoshida, K.; Shinmyo, A. Expression of a foreign gene in Chlamydomonas reinhardtii chloroplast. J. Biosci. Bioeng. 1999, 87, 307–314. [Google Scholar] [CrossRef]
- Michelet, L.; Lefebvre-Legendre, L.; Burr, S.; Rochaix, J.-D.; Goldschmidt-Clermont, M.P. Enhanced chloroplast transgene expression in a nuclear mutant of Chlamydomonas. Plant Biotechnol. J. 2011, 9, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasala, B.A.; Muto, M.; Sullivan, J.; Mayfield, S.P. Improved heterologous protein expression in the chloroplast of Chlamydomonas reinhardtii through promoter and 5′ untranslated region optimization. Plant Biotechnol. J. 2011, 9, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Bruick, R.K.; Mayfield, S.P. Processing of the psbA 5′ Untranslated Region in Chlamydomonas reinhardtii Depends upon Factors Mediating Ribosome Association. J. Cell Biol. 1998, 143, 1145–1153. [Google Scholar] [CrossRef] [Green Version]
- Gimpel, J.A.; Mayfield, S.P. Analysis of heterologous regulatory and coding regions in algal chloroplasts. Appl. Microbiol. Biotechnol. 2013, 97, 4499–4510. [Google Scholar] [CrossRef] [PubMed]
- Gimpel, J.A.; Hyun, J.S.; Schoepp, N.G.; Mayfield, S.P. Production of recombinant proteins in microalgae at pilot greenhouse scale. Biotechnol. Bioeng. 2015, 112, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Shahar, N.; Elman, T.; Williams-Carrier, R.; Ben-Zvi, O.; Yacoby, I.; Barkan, A. Use of plant chloroplast RNA-binding proteins as orthogonal activators of chloroplast transgenes in the green alga Chlamydomonas reinhardtii. Algal Res. 2021, 60, 102535. [Google Scholar] [CrossRef]
- Kasai, S.; Yoshimura, S.; Ishikura, K.; Takaoka, Y.; Kobayashi, K.; Kato, K.; Shinmyo, A. Effect of coding regions on chloroplast gene expression in Chlamydomonas reinhardtii. J. Biosci. Bioeng. 2003, 95, 276–282. [Google Scholar] [CrossRef]
- Crozet, P.; Navarro, F.J.; Willmund, F.; Mehrshahi, P.; Bakowski, K.; Lauersen, K.; Pérez-Pérez, M.-E.; Auroy, P.; Rovira, A.G.; Sauret-Gueto, S.; et al. Birth of a Photosynthetic Chassis: A MoClo Toolkit Enabling Synthetic Biology in the Microalga Chlamydomonas reinhardtii. ACS Synth. Biol. 2018, 7, 2074–2086. [Google Scholar] [CrossRef] [Green Version]
- Oey, M.; Ross, I.L.; Hankamer, B. Gateway-Assisted Vector Construction to Facilitate Expression of Foreign Proteins in the Chloroplast of Single Celled Algae. PLoS ONE 2014, 9, e86841. [Google Scholar] [CrossRef]
- Mayfield, S.P.; Schultz, J. Development of a luciferase reporter gene, luxCt, for Chlamydomonas reinhardtii chloroplast. Plant J. 2004, 37, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Braun-Galleani, S.; Baganz, F.; Purton, S. Improving recombinant protein production in the Chlamydomonas reinhardtii chloroplast using vivid Verde Fluorescent Protein as a reporter. Biotechnol. J. 2015, 10, 1289–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Kim, K.W.; Kwon, Y.M.; Kim, J.Y.H. mCherry Protein as an In Vivo Quantitative Reporter of Gene Expression in the Chloroplast of Chlamydomonas reinhardtii. Mol. Biotechnol. 2020, 62, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, S.E.C.; Hankamer, B.; Oey, M. Optimising light conditions increases recombinant protein production in Chlamydomonas reinhardtii chloroplasts. Algal Res. 2018, 32, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Geisler, K.; Leontidou, T.; Young, R.E.; Vonlanthen, S.E.; Purton, S.; Abell, C.; Smith, A.G. Droplet-based microfluidic screening and sorting of microalgal populations for strain engineering applications. Algal Res. 2021, 56, 102293. [Google Scholar] [CrossRef] [PubMed]
- Rochaix, J.-D.; Surzycki, R.; Ramundo, S. Regulated Chloroplast Gene Expression in Chlamydomonas. Methods Mol. Biol. 2021, 2317, 305–318. [Google Scholar] [CrossRef]
- Young, R.; Purton, S. CITRIC: Cold-inducible translational readthrough in the chloroplast of Chlamydomonas reinhardtii using a novel temperature-sensitive transfer RNA. Microb. Cell Fact. 2018, 17, 186. [Google Scholar] [CrossRef] [Green Version]
- Macedo-Osorio, K.S.; Pérez-España, V.H.; Garibay-Orijel, C.; Guzmán-Zapata, D.; Durán-Figueroa, N.V.; Badillo-Corona, J.A. Intercistronic expression elements (IEE) from the chloroplast of Chlamydomonas reinhardtii can be used for the expression of foreign genes in synthetic operons. Plant Mol. Biol. 2018, 98, 303–317. [Google Scholar] [CrossRef]
- Jones, C.S.; Luong, T.; Hannon, M.; Tran, M.; Gregory, J.A.; Shen, Z.; Briggs, S.P.; Mayfield, S.P. Heterologous expression of the C-terminal antigenic domain of the malaria vaccine candidate Pfs48/45 in the green algae Chlamydomonas reinhardtii. Appl. Microbiol. Biotechnol. 2013, 97, 1987–1995. [Google Scholar] [CrossRef]
- Munjal, N.; Garzon-Sanabria, A.J.; Quinones, K.W.; Gregory, J.; Nikolov, Z.L. Light-Induced Production of An Antibody Fragment and Malaria Vaccine Antigen from Chlamydomonas reinhardtii. Processes 2014, 2, 625–638. [Google Scholar] [CrossRef] [Green Version]
- ‘t Lam, G.P.; van der Kolk, J.A.; Chordia, A.; Vermuë, M.H.; Olivieri, G.; Eppink, M.H.M.; Wijffels, R.H. Mild and Selective Protein Release of Cell Wall Deficient Microalgae with Pulsed Electric Field. ACS Sustain. Chem. Eng. 2017, 5, 6046–6053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoffels, L.; Finlan, A.; Mannall, G.; Purton, S.; Parker, B. Downstream Processing of Chlamydomonas reinhardtii TN72 for Recombinant Protein Recovery. Front. Bioeng. Biotechnol. 2019, 7, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murbach, T.S.; Glávits, R.; Endres, J.R.; Hirka, G.; Vértesi, A.; Béres, E.; Szakonyiné, I.P. A Toxicological Evaluation of Chlamydomonas reinhardtii, a Green Algae. Int. J. Toxicol. 2018, 37, 53–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niccolai, A.; Zittelli, G.C.; Rodolfi, L.; Biondi, N.; Tredici, M.R. Microalgae of interest as food source: Biochemical composition and digestibility. Algal Res. 2019, 42, 101617. [Google Scholar] [CrossRef]
- Kwon, K.-C.; Lamb, A.; Fox, D.; Jegathese, S.J.P. An evaluation of microalgae as a recombinant protein oral delivery platform for fish using green fluorescent protein (GFP). Fish Shellfish Immunol. 2019, 87, 414–420. [Google Scholar] [CrossRef]
- Taunt, H.N.; Stoffels, L.; Purton, S. Green biologics: The algal chloroplast as a platform for making biopharmaceuticals. Bioengineered 2018, 9, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Yefremova, Y.; Purton, S. The algal chloroplast as a synthetic biology platform for production of therapeutic proteins. Microbiology 2018, 164, 113–121. [Google Scholar]
- Brooks, S.A. Appropriate Glycosylation of Recombinant Proteins for Human Use: Implications of Choice of Expression System. Mol. Biotechnol. 2004, 28, 241–256. [Google Scholar] [CrossRef]
- Mathieu-Rivet, E.; Kiefer-Meyer, M.C.; Vanier, G.; Ovide, C.; Burel, C.; Lerouge, P.; Bardor, M. Protein N-glycosylation in eukaryotic microalgae and its impact on the production of nuclear expressed biopharmaceuticals. Front. Plant Sci. 2014, 5, 359. [Google Scholar] [CrossRef]
- Manuell, A.L.; Beligni, M.V.; Elder, J.H.; Siefker, D.T.; Tran, M.; Weber, A.; McDonald, T.L.; Mayfield, S.P. Robust expression of a bioactive mammalian protein in Chlamydomonas chloroplast. Plant Biotechnol. J. 2007, 5, 402–412. [Google Scholar] [CrossRef]
- Campos-Quevedo, N.; Rosales-Mendoza, S.; Paz-Maldonado, L.M.T.; Martínez-Salgado, L.; Guevara-Arauza, J.C.; Soria-Guerra, R.E. Production of milk-derived bioactive peptides as precursor chimeric proteins in chloroplasts of Chlamydomonas reinhardtii. Plant Cell Tissue Organ Cult. (PCTOC) 2013, 113, 217–225. [Google Scholar] [CrossRef]
- Stoffels, L.; Taunt, H.N.; Charalambous, B.; Purton, S. Synthesis of bacteriophage lytic proteins against Streptococcus pneumoniae in the chloroplast of Chlamydomonas reinhardtii. Plant Biotechnol. J. 2017, 15, 1130–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasala, B.A.; Muto, M.; Lee, P.A.; Jager, M.; Cardoso, R.M.; Behnke, C.A.; Kirk, P.; Hokanson, C.A.; Crea, R.; Mendez, M.; et al. Production of therapeutic proteins in algae, analysis of expression of seven human proteins in the chloroplast of Chlamydomonas reinhardtii. Plant Biotechnol. J. 2010, 8, 719–733. [Google Scholar] [CrossRef] [Green Version]
- Rasala, B.A.; Mayfield, S.P. The microalga Chlamydomonas reinhardtii as a platform for the production of human protein therapeutics. Bioeng. Bugs. 2011, 2, 50–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravi, A.; Guo, S.; Rasala, B.; Tran, M.; Mayfield, S.; Nikolov, Z.L. Separation Options for Phosphorylated Osteopontin from Transgenic Microalgae Chlamydomonas reinhardtii. Int. J. Mol. Sci. 2018, 19, 585. [Google Scholar] [CrossRef] [Green Version]
- Ochoa-Méndez, C.E.; Lara-Hernández, I.; González, L.M.; Aguirre-Bañuelos, P.; Ibarra-Barajas, M.; Castro-Moreno, P.; Gonzalez-Ortega, O.; Soria-Guerra, R.E. Bioactivity of an antihypertensive peptide expressed in Chlamydomonas reinhardtii. J. Biotechnol. 2016, 240, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Carrizalez-López, C.; Gonzalez-Ortega, O.; Ochoa-Méndez, C.E.; Galván-Moreno, F.U.; Rosales-Mendoza, S.; Monreal-Escalante, E.; Bañuelos-Hernandez, B.; Paz-Maldonado, L.M.T.; Castro-Moreno, P.; Ibarra-Barajas, M.; et al. Expression of multiple antihypertensive peptides as a fusion protein in the chloroplast of Chlamydomonas reinhardtii. J. Appl. Phycol. 2018, 30, 1701–1709. [Google Scholar] [CrossRef]
- Tran, M.; Zhou, B.; Pettersson, P.L.; Gonzalez, M.J.; Mayfield, S.P. Synthesis and assembly of a full-length human monoclonal antibody in algal chloroplasts. Biotechnol. Bioeng. 2009, 104, 663–673. [Google Scholar] [CrossRef]
- Tran, M.; Henry, R.E.; Siefker, D.; Van, C.; Newkirk, G.; Kim, J.; Bui, J.; Mayfield, S.P. Production of anti-cancer immunotoxins in algae: Ribosome inactivating proteins as fusion partners. Biotechnol. Bioeng. 2013, 110, 2826–2835. [Google Scholar] [CrossRef]
- Tran, M.; Van, C.; Barrera, D.J.; Pettersson, P.L.; Peinado, C.D.; Bui, J.; Mayfield, S.P. Production of unique immunotoxin cancer therapeutics in algal chloroplasts. Proc. Natl. Acad. Sci. USA 2013, 110, E15–E22. [Google Scholar] [CrossRef] [Green Version]
- Barrera, D.J.; Rosenberg, J.N.; Chiu, J.G.; Chang, Y.-N.; Debatis, M.; Ngoi, S.-M.; Chang, J.T.; Shoemaker, C.B.; Oyler, G.A.; Mayfield, S.P. Algal chloroplast produced camelid VH H antitoxins are capable of neutralizing botulinum neurotoxin. Plant Biotechnol. J. 2015, 13, 117–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zedler, J.A.; Mullineaux, C.W.; Robinson, C. Efficient targeting of recombinant proteins to the thylakoid lumen in Chlamydomonas reinhardtii using a bacterial Tat signal peptide. Algal Res. 2016, 19, 57–62. [Google Scholar] [CrossRef] [Green Version]
- Specht, E.A.; Mayfield, S.P. Algae-based oral recombinant vaccines. Front. Microbiol. 2014, 5, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschl, S.; Ralser, C.; Asam, C.; Gangitano, A.; Huber, S.; Ebner, C.; Bohle, B.; Wolf, M.; Briza, P.; Ferreira, F.; et al. Expression and Characterization of Functional Recombinant Bet v 1.0101 in the Chloroplast of Chlamydomonas reinhardtii. Int. Arch. Allergy Immunol. 2017, 173, 44–50. [Google Scholar] [CrossRef]
- Sun, M.; Qian, K.; Su, N.; Chang, H.; Liu, J.; Shen, G. Foot-and-mouth disease virus VP1 protein fused with cholera toxin B subunit expressed in Chlamydomonas reinhardtii chloroplast. Biotechnol. Lett. 2003, 25, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Dreesen, I.A.; Hamri, G.C.-E.; Fussenegger, M. Heat-stable oral alga-based vaccine protects mice from Staphylococcus aureus infection. J. Biotechnol. 2010, 145, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Brandsma, M.; Tremblay, R.; Maxwell, D.; Jevnikar, A.M.; Huner, N.; Ma, S. A novel expression platform for the production of diabetes-associated autoantigen human glutamic acid decarboxylase (hGAD65). BMC Biotechnol. 2008, 8, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beltrán-López, J.I.; Romero-Maldonado, A.; Monreal-Escalante, E.; Bañuelos-Hernández, B.; Paz-Maldonado, L.M.; Rosales-Mendoza, S. Chlamydomonas reinhardtii chloroplasts express an orally immunogenic protein targeting the p210 epitope implicated in atherosclerosis immunotherapies. Plant Cell Rep. 2016, 35, 1133–1141. [Google Scholar] [CrossRef]
- Gregory, J.; Li, F.; Tomosada, L.M.; Cox, C.J.; Topol, A.B.; Vinetz, J.M.; Mayfield, S. Algae-Produced Pfs25 Elicits Antibodies That Inhibit Malaria Transmission. PLoS ONE 2012, 7, e37179. [Google Scholar] [CrossRef] [Green Version]
- Gregory, J.A.; Topol, A.B.; Doerner, D.Z.; Mayfield, S. Alga-Produced Cholera Toxin-Pfs25 Fusion Proteins as Oral Vaccines. Appl. Environ. Microbiol. 2013, 79, 3917–3925. [Google Scholar] [CrossRef] [Green Version]
- Patra, K.P.; Li, F.; Carter, D.; Gregory, J.A.; Baga, S.; Reed, S.G.; Mayfield, S.P.; Vinetz, J.M. Alga-Produced Malaria Transmission-Blocking Vaccine Candidate Pfs25 Formulated with a Human Use-Compatible Potent Adjuvant Induces High-Affinity Antibodies That Block Plasmodium falciparum Infection of Mosquitoes. Infect. Immun. 2015, 83, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Shamriz, S.; Ofoghi, H. Engineering the chloroplast of Chlamydomonas reinhardtii to express the recombinant PfCelTOS-Il2 antigen-adjuvant fusion protein. J. Biotechnol. 2018, 266, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Shamriz, S.; Ofoghi, H. Expression of Recombinant PfCelTOS Antigen in the Chloroplast of Chlamydomonas reinhardtii and its Potential Use in Detection of Malaria. Mol. Biotechnol. 2019, 61, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Berndt, A.J.; Smalley, T.N.; Ren, B.; Simkovsky, R.; Badary, A.; Sproles, A.E.; Fields, F.J.; Torres-Tiji, Y.; Heredia, V.; Mayfield, S.P. Recombinant production of a functional SARS-CoV-2 spike receptor binding domain in the green algae Chlamydomonas reinhardtii. PLoS ONE 2021, 16, e0257089. [Google Scholar] [CrossRef] [PubMed]
- Charoonnart, P.; Purton, S.; Saksmerprome, V. Applications of Microalgal Biotechnology for Disease Control in Aquaculture. Biology 2018, 7, 24. [Google Scholar] [CrossRef] [Green Version]
- Kiataramgul, A.; Maneenin, S.; Purton, S.; Areechon, N.; Hirono, I.; Brocklehurst, T.W.; Unajak, S. An oral delivery system for controlling white spot syndrome virus infection in shrimp using transgenic microalgae. Aquaculture 2020, 521, 735022. [Google Scholar] [CrossRef]
- Charoonnart, P.; Tuant, H.; Webb, C.; Purton, S.; Robinson, C.; Saksmerprom, E.V. Enhanced production of antiviral dsRNA in the Chlamydomonas reinhardtii chloroplast via a novel convergent promoter expression system. bioRxiv 2022. [Google Scholar] [CrossRef]
- He, D.-M.; Qian, K.-X.; Shen, G.-F.; Zhang, Z.-F.; Li, Y.-N.; Su, Z.-L.; Shao, H.-B. Recombination and expression of classical swine fever virus (CSFV) structural protein E2 gene in Chlamydomonas reinhardtii chroloplasts. Colloids Surf. B Biointerfaces 2007, 55, 26–30. [Google Scholar] [CrossRef]
- Yoon, S.-M.; Kim, S.Y.; Li, K.F.; Yoon, B.H.; Choe, S.; Kuo, M.M.-C. Transgenic microalgae expressing Escherichia coli AppA phytase as feed additive to reduce phytate excretion in the manure of young broiler chicks. Appl. Microbiol. Biotechnol. 2011, 91, 553–563. [Google Scholar] [CrossRef]
- Jackson, H.O.; Taunt, H.N.; Mordaka, P.M.; Smith, A.G.; Purton, S. The Algal Chloroplast as a Testbed for Synthetic Biology Designs Aimed at Radically Rewiring Plant Metabolism. Front. Plant Sci. 2021, 12, 708370. [Google Scholar] [CrossRef]
- Tan, C.-P.; Zhao, F.Q.; Su, Z.L.; Liang, C.W.; Qin, S. Expression of β-carotene hydroxylase gene (crtR-B) from the green alga Haematococcus pluvialis in chloroplasts of Chlamydomonas reinhardtii. J. Appl. Phycol. 2007, 19, 347–355. [Google Scholar] [CrossRef]
- Agrawal, S.; Karcher, D.; Ruf, S.; Erban, A.; Hertle, A.P.; Kopka, J.; Bock, R. Riboswitch-mediated inducible expression of an astaxanthin biosynthetic operon in plastids. Plant Physiol. 2021, 188, 637–652. [Google Scholar] [CrossRef] [PubMed]
- Lauersen, K.J. Eukaryotic microalgae as hosts for light-driven heterologous isoprenoid production. Planta 2018, 249, 155–180. [Google Scholar] [CrossRef] [PubMed]
- Cox-Georgian, D.; Ramadoss, N.; Dona, C.; Basu, C. Therapeutic and Medicinal Uses of Terpenes. In Medicinal Plants: From Farm to Pharmacy; Springer: Cham, Switzerland, 2019; pp. 333–359. [Google Scholar] [CrossRef]
- Zedler, J.A.Z.; Gangl, D.; Hamberger, B.; Purton, S.; Robinson, C.; Hamberger, B. Stable expression of a bifunctional diterpene synthase in the chloroplast of Chlamydomonas reinhardtii. J. Appl. Phycol. 2015, 27, 2271–2277. [Google Scholar] [CrossRef]
- Gangl, D.; Zedler, J.A.Z.; Wlodarczyk, A.; Jensen, P.E.; Purton, S.; Robinson, C. Expression and membrane-targeting of an active plant cytochrome P450 in the chloroplast of the green alga Chlamydomonas reinhardtii. Phytochemistry 2014, 110, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Zedler, J.A.Z.; Gangl, D.; Guerra, T.; Santos, E.; Verdelho, V.V.; Robinson, C. Pilot-scale cultivation of wall-deficient transgenic Chlamydomonas reinhardtii strains expressing recombinant proteins in the chloroplast. Appl. Microbiol. Biotechnol. 2016, 100, 7061–7070. [Google Scholar] [CrossRef]
- Georgianna, D.R.; Hannon, M.J.; Marcuschi, M.; Wu, S.; Botsch, K.; Lewis, A.J.; Hyun, J.; Mendez, M.; Mayfield, S.P. Production of recombinant enzymes in the marine alga Dunaliella tertiolecta. Algal Res. 2013, 2, 2–9. [Google Scholar] [CrossRef]
- Faè, M.; Accossato, S.; Cella, R.; Fontana, F.; Goldschmidt-Clermont, M.; Leelavathi, S.; Reddy, V.S.; Longoni, P. Comparison of transplastomic Chlamydomonas reinhardtii and Nicotiana tabacum expression system for the production of a bacterial endoglucanase. Appl. Microbiol. Biotechnol. 2017, 101, 4085–4092. [Google Scholar] [CrossRef]
- Richter, L.V.; Yang, H.; Yazdani, M.; Hanson, M.R.; Ahner, B.A. A downstream box fusion allows stable accumulation of a bacterial cellulase in Chlamydomonas reinhardtii chloroplasts. Biotechnol. Biofuels 2018, 11, 133. [Google Scholar] [CrossRef] [Green Version]
- Benedetti, M.; Barera, S.; Longoni, P.; Guardini, Z.; Garcia, N.H.; Bolzonella, D.; Arredondo, D.L.; Herrera-Estrella, L.; Goldschmidt-Clermont, M.; Bassi, R.; et al. A microalgal-based preparation with synergistic cellulolytic and detoxifying action towards chemical-treated lignocellulose. Plant Biotechnol. J. 2020, 19, 124–137. [Google Scholar] [CrossRef]
- Ipsen, J.; Hallas-Møller, M.; Brander, S.; Leggio, L.L.; Johansen, K.S. Lytic polysaccharide monooxygenases and other histidine-brace copper proteins: Structure, oxygen activation and biotechnological applications. Biochem. Soc. Trans. 2021, 49, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Chandra, R. Ligninolytic enzymes and its mechanisms for degradation of lignocellulosic waste in environment. Heliyon 2020, 6, e03170. [Google Scholar] [CrossRef] [PubMed]
- Merchant, S.S.; Schmollinger, S.; Strenkert, D.; Moseley, J.L.; Blaby-Haas, C.E. From economy to luxury: Copper homeostasis in Chlamydomonas and other algae. Biochim. Biophys. Acta 2020, 1867, 118822. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Odom, O.W.; Thangamani, S.; Herrin, D.L. Toward mosquito control with a green alga: Expression of Cry toxins of Bacillus thuringiensis subsp. israelensis (Bti) in the chloroplast of Chlamydomonas. J. Appl. Phycol. 2017, 29, 1377–1389. [Google Scholar] [CrossRef]
- Kang, S.; Odom, O.W.; Malone, C.L.; Thangamani, S.; Herrin, D.L. Expression of a Synthetic Gene for the Major Cytotoxin (Cyt1Aa) of Bacillus thuringiensis subsp. israelensis in the Chloroplast of Wild-Type Chlamydomonas. Biology 2018, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Fuentes, P.; Zhou, F.; Erban, A.; Karcher, D.; Kopka, J.; Bock, R. A new synthetic biology approach allows transfer of an entire metabolic pathway from a medicinal plant to a biomass crop. eLife 2016, 5, e13664. [Google Scholar] [CrossRef]
- Lu, Y.; Rijzaani, H.; Karcher, D.; Ruf, S.; Bock, R. Efficient metabolic pathway engineering in transgenic tobacco and tomato plastids with synthetic multigene operons. Proc. Natl. Acad. Sci. USA 2013, 110, E623–E632. [Google Scholar] [CrossRef] [Green Version]
- Harada, H.; Maoka, T.; Osawa, A.; Hattan, J.-I.; Kanamoto, H.; Shindo, K.; Otomatsu, T.; Misawa, N. Construction of transplastomic lettuce (Lactuca sativa) dominantly producing astaxanthin fatty acid esters and detailed chemical analysis of generated carotenoids. Transgenic Res. 2014, 23, 303–315. [Google Scholar] [CrossRef]
- Dahlin, L.R.; Gerritsen, A.T.; Henard, C.A.; Van Wychen, S.; Linger, J.G.; Kunde, Y.; Hovde, B.; Starkenburg, S.R.; Posewitz, M.C.; Guarnieri, M.T. Development of a high-productivity, halophilic, thermotolerant microalga Picochlorum renovo. Commun. Biol. 2019, 2, 388–389. [Google Scholar] [CrossRef]
- Zhou, F.; Karcher, D.; Bock, R. Identification of a plastid intercistronic expression element (IEE) facilitating the expression of stable translatable monocistronic mRNAs from operons. Plant J. 2007, 52, 961–972. [Google Scholar] [CrossRef] [Green Version]
- Legen, J.; Ruf, S.; Kroop, X.; Wang, G.; Barkan, A.; Bock, R.; Schmitz-Linneweber, C. Stabilization and translation of synthetic operon-derived mRNA s in chloroplasts by sequences representing PPR protein-binding sites. Plant J. 2018, 94, 8–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimpel, J.A.; Nour-Eldin, H.H.; Scranton, M.A.; Li, D.; Mayfield, S.P. Refactoring the Six-Gene Photosystem II Core in the Chloroplast of the Green Algae Chlamydomonas reinhardtii. ACS Synth. Biol. 2016, 5, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Larrea-Alvarez, M.; Purton, S. Multigenic engineering of the chloroplast genome in the green alga Chlamydomonas reinhardtii. Microbiology 2020, 166, 510–515. [Google Scholar] [CrossRef]
- Ramundo, S.; Casero, D.; Mühlhaus, T.; Hemme, D.; Sommer, F.; Crèvecoeur, M.; Rahire, M.; Schroda, M.; Rusch, J.; Goodenough, U.; et al. Conditional Depletion of the Chlamydomonas Chloroplast ClpP Protease Activates Nuclear Genes Involved in Autophagy and Plastid Protein Quality Control. Plant Cell 2014, 26, 2201–2222. [Google Scholar] [CrossRef] [Green Version]
- Perlaza, K.; Toutkoushian, H.; Boone, M.; Lam, M.; Iwai, M.; Jonikas, M.C.; Walter, P.; Ramundo, S. The Mars1 kinase confers photoprotection through signaling in the chloroplast unfolded protein response. eLife 2019, 8, e49577. [Google Scholar] [CrossRef]
- Kuchka, M.; Goldschmidt-Clermont, M.; van Dillewijn, J.; Rochaix, J.-D. Mutation at the chlamydomonas nuclear NAC2 locus specifically affects stability of the chloroplast psbD transcript encoding polypeptide D2 of PS II. Cell 1989, 58, 869–876. [Google Scholar] [CrossRef]
- Boudreau, E.; Nickelsen, J.; Lemaire, S.D.; Ossenbühl, F.; Rochaix, J. The Nac2 gene of Chlamydomonas encodes a chloroplast TPR-like protein involved in psbD mRNA stability. EMBO J. 2000, 19, 3366–3376. [Google Scholar] [CrossRef] [Green Version]
- Surzycki, R.; Cournac, L.; Peltier, G.; Rochaix, J.D. Potential for hydrogen production with inducible chloroplast gene expression in Chlamydomonas. Proc. Natl. Acad. Sci. USA 2007, 104, 17548–17553. [Google Scholar] [CrossRef] [Green Version]
- Croft, M.T.; Lawrence, A.D.; Raux-Deery, E.; Warren, M.J.; Smith, A.G. Algae acquire vitamin B12 through a symbiotic relationship with bacteria. Nature 2005, 438, 90–93. [Google Scholar] [CrossRef]
- Helliwell, K.E.; Scaife, M.A.; Sasso, S.; Araujo, A.P.U.; Purton, S.; Smith, A. Unraveling Vitamin B12-Responsive Gene Regulation in Algae. Plant Physiol. 2014, 165, 388–397. [Google Scholar] [CrossRef] [Green Version]
- Ramundo, S.; Rochaix, J.-D. Controlling Expression of Genes in the Unicellular Alga Chlamydomonas reinhardtii with a Vitamin-Repressible Riboswitch. Methods Enzymol. 2015, 550, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Croft, M.T.; Moulin, M.; Webb, M.E.; Smith, A.G. Thiamine biosynthesis in algae is regulated by riboswitches. Proc. Natl. Acad. Sci. USA 2007, 104, 20770–20775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrera-Pacheco, S.E.; Hankamer, B.; Oey, M. Light and heat-shock mediated TDA1 overexpression as a tool for controlled high-yield recombinant protein production in Chlamydomonas reinhardtii chloroplasts. Algal Res. 2020, 48, 101921. [Google Scholar] [CrossRef]
- Raynaud, C.; Loiselay, C.; Wostrikoff, K.; Kuras, R.; Girard-Bascou, J.; Wollman, F.-A.; Choquet, Y. Evidence for regulatory function of nucleus-encoded factors on mRNA stabilization and translation in the chloroplast. Proc. Natl. Acad. Sci. USA 2007, 104, 9093–9098. [Google Scholar] [CrossRef] [Green Version]
- Eberhard, S.; Loiselay, C.; Drapier, D.; Bujaldon, S.; Girard-Bascou, J.; Kuras, R.; Choquet, Y.; Wollman, F.-A. Dual functions of the nucleus-encoded factor TDA1 in trapping and translation activation of atpA transcripts in Chlamydomonas reinhardtii chloroplasts. Plant J. 2011, 67, 1055–1066. [Google Scholar] [CrossRef]
- Yu, W.; Spreitzer, R.J. Chloroplast heteroplasmicity is stabilized by an amber-suppressor tryptophan tRNA(CUA). Proc. Natl. Acad. Sci. USA 1992, 89, 3904–3907. [Google Scholar] [CrossRef] [Green Version]
- Marschalek, R.; Kalpaxis, D.; Dingermann, T. Temperature sensitive synthesis of transfer RNAs in vivo in Saccharomyces cerevisiae. EMBO J. 1990, 9, 1253–1258. [Google Scholar] [CrossRef]
- Lee, H.; Bingham, S.E.; Webber, A.N. Function of 3’ non-coding sequences and stop codon usage in expression of the chloroplast psaB gene in Chlamydomonas reinhardtii. Plant Mol. Biol. 1996, 31, 337–354. [Google Scholar] [CrossRef]
- Young, R.E.B.; Purton, S. Codon reassignment to facilitate genetic engineering and biocontainment in the chloroplast of Chlamydomonas reinhardtii. Plant Biotechnol. J. 2016, 14, 1251–1260. [Google Scholar] [CrossRef] [Green Version]
- Gressel, J.; van der Vlugt, C.J.B.; Bergmans, H.E.N. Environmental risks of large scale cultivation of microalgae: Mitigation of spills. Algal Res. 2013, 2, 286–298. [Google Scholar] [CrossRef]
- Guiry, M.D. How many species of algae are there? J. Phycol. 2012, 48, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Mosey, M.; Douchi, D.; Knoshaug, E.P.; Laurens, L.M. Methodological review of genetic engineering approaches for non-model algae. Algal Res. 2021, 54, 102221. [Google Scholar] [CrossRef]
- Liu, T.; Cui, Y.; Jia, X.; Zhang, J.; Li, R.; Yu, Y.; Jia, S.; Qu, J.; Wang, X. OGDA: A comprehensive organelle genome database for algae. Database 2020, 2020, baaa097. [Google Scholar] [CrossRef] [PubMed]
- Sablok, G.; Chen, T.-W.; Lee, C.-C.; Yang, C.; Gan, R.-C.; Wegrzyn, J.L.; Porta, N.L.; Nayak, K.C.; Huang, P.-J.; Varotto, C.; et al. ChloroMitoCU: Codon patterns across organelle genomes for functional genomics and evolutionary applications. DNA Res. 2017, 24, 327–332. [Google Scholar] [CrossRef] [Green Version]
- Barclay, W.; Apt, K. Strategies for Bioprospecting Microalgae for Potential Commercial Applications. In Handbook of Microalgal Culture; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 69–79. [Google Scholar] [CrossRef]
- Cui, Y.; Qin, S.; Jiang, P. Chloroplast Transformation of Platymonas (Tetraselmis) subcordiformis with the bar Gene as Selectable Marker. PLoS ONE 2014, 9, e98607. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Zhao, J.; Hou, S.; Qin, S. Enhanced green fluorescent protein (egfp) gene expression in Tetraselmis subcordiformis chloroplast with endogenous regulators. World J. Microbiol. Biotechnol. 2016, 32, 83. [Google Scholar] [CrossRef]
- Li, D.; Han, X.; Zuo, J.; Xie, L.; He, R.; Gao, J.; Chang, L.; Yuan, L.; Cao, M. Construction of rice site-specific chloroplast transformation vector and transient expression of EGFP gene in Dunaliella salina. J. Biomed. Nanotechnol. 2011, 7, 801–806. [Google Scholar] [CrossRef]
- Douchi, D.; Mosey, M.; Astling, D.P.; Knoshaug, E.P.; Nag, A.; McGowen, J.; Laurens, L.M. Nuclear and chloroplast genome engineering of a productive non-model alga Desmodesmus armatus: Insights into unusual and selective acquisition mechanisms for foreign DNA. Algal Res. 2021, 53, 102152. [Google Scholar] [CrossRef]
- Gutiérrez, C.L.; Gimpel, J.; Escobar, C.; Marshall, S.H.; Henríquez, V. Chloroplast genetic tool for the green microalgae haematococcus pluvialis (chlorophyceae, volvocales)(1). J. Phycol. 2012, 48, 976–983. [Google Scholar] [CrossRef]
- Galarza, J.I.; Gimpel, J.A.; Rojas, V.; Arredondo-Vega, B.O.; Henríquez, V. Over-accumulation of astaxanthin in Haematococcus pluvialis through chloroplast genetic engineering. Algal Res. 2018, 31, 291–297. [Google Scholar] [CrossRef]
- Wang, K.; Cui, Y.; Wang, Y.; Gao, Z.; Liu, T.; Meng, C.; Qin, S. Chloroplast Genetic Engineering of a Unicellular Green Alga Haematococcus pluvialis with Expression of an Antimicrobial Peptide. Mar. Biotechnol. 2020, 22, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Chuai, W.; Li, K.; Hou, G.; Wu, M.; Chen, J.; Wang, H.; Jia, J.; Han, D.; Hu, Q. Ultrahigh-cell-density heterotrophic cultivation of the unicellular green alga Chlorella sorokiniana for biomass production. Biotechnol. Bioeng. 2021, 118, 4138–4151. [Google Scholar] [CrossRef] [PubMed]
- Levin, G.; Kulikovsky, S.; Liveanu, V.; Eichenbaum, B.; Meir, A.; Isaacson, T.; Tadmor, Y.; Adir, N.; Schuster, G. The desert green algae Chlorella ohadii thrives at excessively high light intensities by exceptionally enhancing the mechanisms that protect photosynthesis from photoinhibition. Plant J. 2021, 106, 1260–1277. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Wan, D. The complete chloroplast genome sequence of Chlorella vulgaris and phylogenetic analysis. Mitochondrial DNA Part B 2020, 5, 2713–2715. [Google Scholar] [CrossRef] [PubMed]
- Weber, S.; Grande, P.M.; Blank, L.M.; Klose, H. Insights into cell wall disintegration of Chlorella vulgaris. PLoS ONE 2022, 17, e0262500. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Gao, Z.; Wang, Y.; Meng, C.; Li, J.; Qin, S.; Cui, Y. The chloroplast genetic engineering of a unicellular green alga Chlorella vulgaris with two foreign peptides co-expression. Algal Res. 2021, 54, 102214. [Google Scholar] [CrossRef]
- Wakasugi, T.; Nagai, T.; Kapoor, M.; Sugita, M.; Ito, M.; Ito, S.; Tsudzuki, J.; Nakashima, K.; Tsudzuki, T.; Suzuki, Y.; et al. Complete nucleotide sequence of the chloroplast genome from the green alga Chlorella vulgaris: The existence of genes possibly involved in chloroplast division. Proc. Natl. Acad. Sci. USA 1997, 94, 5967–5972. [Google Scholar] [CrossRef] [Green Version]
- Nawkarkar, P.; Chugh, S.; Sharma, S.; Jain, M.; Kajla, S.; Kumar, S. Characterization of the Chloroplast Genome Facilitated the Transformation of Parachlorella kessleri-I, A Potential Marine Alga for Biofuel Production. Curr. Genom. 2020, 21, 610–623. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, K.; Xu, W.; Wang, Y.; Gao, Z.; Cui, H.; Meng, C.; Qin, S. Plastid Engineering of a Marine Alga, Nannochloropsis gaditana, for Co-Expression of Two Recombinant Peptides. J. Phycol. 2021, 57, 569–576. [Google Scholar] [CrossRef]
- Xie, W.-H.; Zhu, C.-C.; Zhang, N.-S.; Li, D.-W.; Yang, W.-D.; Liu, J.-S.; Sathishkumar, R.; Li, H.-Y. Construction of Novel Chloroplast Expression Vector and Development of an Efficient Transformation System for the Diatom Phaeodactylum tricornutum. Mar. Biotechnol. 2014, 16, 538–546. [Google Scholar] [CrossRef] [Green Version]
- de Grahl, I.; Reumann, S. Stramenopile microalgae as “green biofactories” for recombinant protein production. World J. Microbiol. Biotechnol. 2021, 37, 163. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Nesamma, A.A.; Lali, A.M.; Jutur, P.P.; Prakash, G. The chloroplast genome of a resilient chlorophycean microalga Asterarcys sp. Algal Res. 2020, 49, 101952. [Google Scholar] [CrossRef]
- Benedetti, M.; Vecchi, V.; Barera, S.; Dall’Osto, L. Biomass from microalgae: The potential of domestication towards sustainable biofactories. Microb. Cell Fact. 2018, 17, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esland, L.; Larrea-Alvarez, M.; Purton, S. Selectable Markers and Reporter Genes for Engineering the Chloroplast of Chlamydomonas reinhardtii. Biology 2018, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, B.M.; Mikkelson, K.L.; Gutierrez, N.M.; Cunningham, J.L.; Wolff, K.L.; Szyjka, S.J.; Yohn, C.B.; Redding, K.E.; Mendez, M.J. An exogenous chloroplast genome for complex sequence manipulation in algae. Nucleic Acids Res. 2011, 40, 2782–2792. [Google Scholar] [CrossRef] [Green Version]
- Jakubiec, A.; Sarokina, A.; Choinard, S.; Vlad, F.; Malcuit, I.; Sorokin, A.P. Replicating minichromosomes as a new tool for plastid genome engineering. Nat. Plants 2021, 7, 932–941. [Google Scholar] [CrossRef]
- Occhialini, A.; Pfotenhauer, A.C.; Li, L.; Harbison, S.A.; Lail, A.J.; Burris, J.N.; Piasecki, C.; Piatek, A.A.; Daniell, H.; Stewart, C.N.; et al. Mini-synplastomes for plastid genetic engineering. Plant Biotechnol. J. 2021, 20, 360–373. [Google Scholar] [CrossRef]
- Rumin, J.; Junior, R.G.D.O.; Bérard, J.-B.; Picot, L. Improving Microalgae Research and Marketing in the European Atlantic Area: Analysis of Major Gaps and Barriers Limiting Sector Development. Mar. Drugs 2021, 19, 319. [Google Scholar] [CrossRef]
- Naduthodi, M.I.S.; Claassens, N.J.; D’Adamo, S.; van der Oost, J.; Barbosa, M.J. Synthetic Biology Approaches To Enhance Microalgal Productivity. Trends Biotechnol. 2021, 39, 1019–1036. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cutolo, E.A.; Mandalà, G.; Dall’Osto, L.; Bassi, R. Harnessing the Algal Chloroplast for Heterologous Protein Production. Microorganisms 2022, 10, 743. https://doi.org/10.3390/microorganisms10040743
Cutolo EA, Mandalà G, Dall’Osto L, Bassi R. Harnessing the Algal Chloroplast for Heterologous Protein Production. Microorganisms. 2022; 10(4):743. https://doi.org/10.3390/microorganisms10040743
Chicago/Turabian StyleCutolo, Edoardo Andrea, Giulia Mandalà, Luca Dall’Osto, and Roberto Bassi. 2022. "Harnessing the Algal Chloroplast for Heterologous Protein Production" Microorganisms 10, no. 4: 743. https://doi.org/10.3390/microorganisms10040743
APA StyleCutolo, E. A., Mandalà, G., Dall’Osto, L., & Bassi, R. (2022). Harnessing the Algal Chloroplast for Heterologous Protein Production. Microorganisms, 10(4), 743. https://doi.org/10.3390/microorganisms10040743