
Could Pontimonas Harbour Halophilic Members Able to Withstand Very Broad Salinity Variations?

 ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Characterisation
2.2. DNA Extraction, 16S rDNA Amplicon Library Preparation and Sequencing
2.3. Sequence Processing and Data Analysis
2.4. Statistical Methods
3. Results
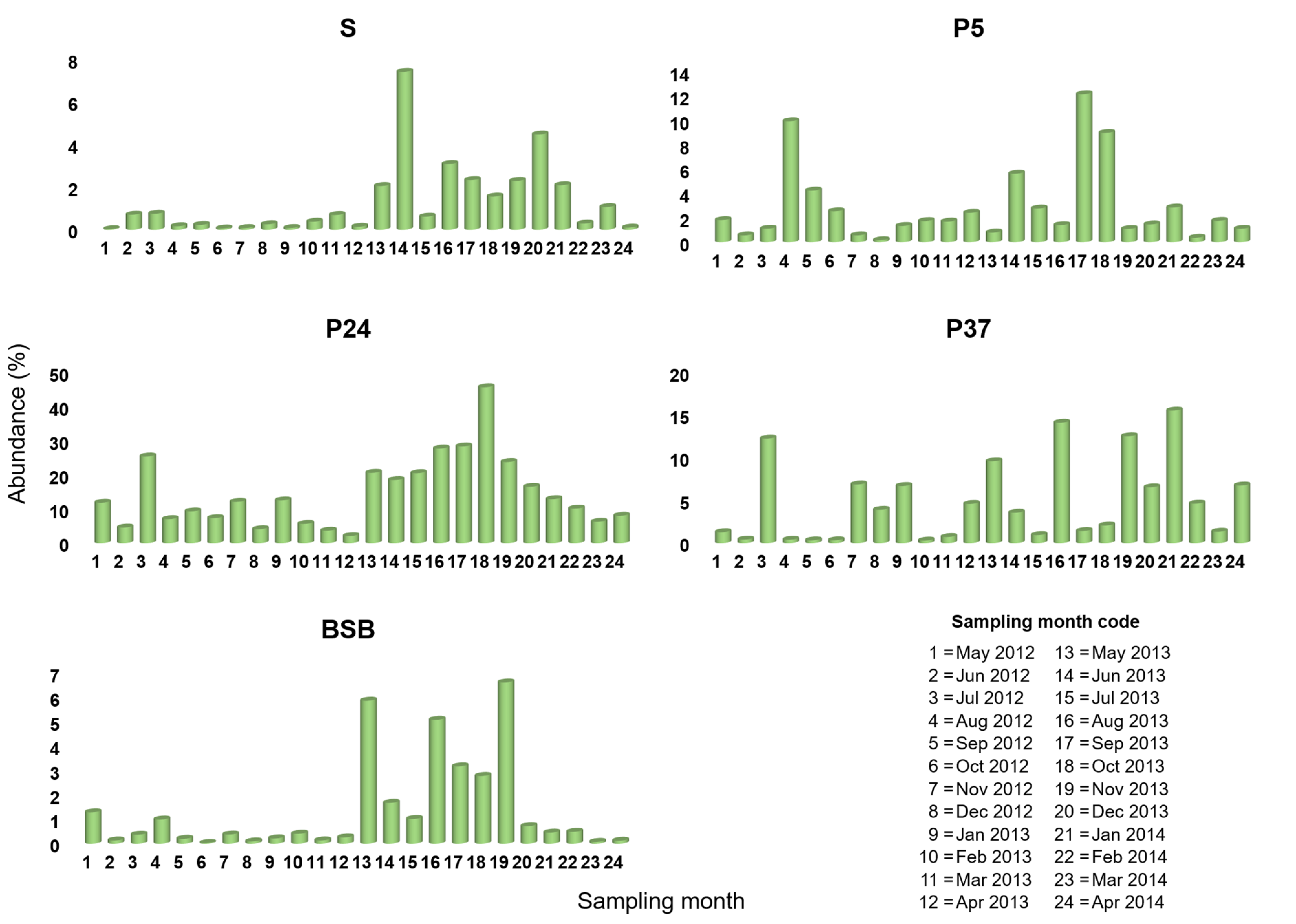
3.1. Spatio-Temporal Variation in Pontimonas
3.2. Environmental Parameters Affecting Pontimonas Occurrence
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- LPSN-List of Prokaryotic Names with Standing in Nomenclature, Family Enterobacteriaceae. Available online: https://lpsn.dsmz.de/genus/pontimonas (accessed on 1 February 2022).
- Jang, G.I.; Cho, Y.; Cho, B.C. Pontimonas salivibrio gen. nov., sp. nov., a new member of the family Microbacteriaceae isolated from a seawater reservoir of a solar saltern. Int. J. Syst. Evol. Microbiol. 2013, 63, 2124–2131. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.C.; Jang, G.I.; Hwang, C.Y. Pontimonas. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.E.B., Ed.; John Wiley & Sons: New York, NY, USA, 2015; pp. 1–5. [Google Scholar]
- Cho, B.C.; Hardies, S.C.; Jang, G.I.; Hwang, C.Y. Complete genome of streamlined marine actinobacterium Pontimonas salivibrio strain CL-TW6T adapted to coastal planktonic lifestyle. BMC Genom. 2018, 19, 625. [Google Scholar] [CrossRef] [PubMed]
- Leoni, C.; Volpicella, M.; Fosso, B.; Manzari, C.; Piancone, E.; Dileo, M.C.G.; Arcadi, E.; Yakimov, M.; Pesole, G.; Ceci, L.R. A differential metabarcoding approach to describe taxonomy profiles of bacteria and archaea in the saltern of Margherita di Savoia (Italy). Microorganisms 2020, 8, 936. [Google Scholar] [CrossRef] [PubMed]
- Al-Mailem, D.M.; Kansour, M.K.; Radwan, S.S. Capabilities and limitations of DGGE for the analysis of hydrocarbonoclastic prokaryotic communities directly in environmental samples. Microbiol. Open 2017, 6, e00495. [Google Scholar] [CrossRef]
- Selivanova, E.A.; Poshvina, D.V.; Khlopko, Y.A.; Gogoleva, N.E.; Plotnikov, A.O. Diversity of prokaryotes in planktonic communities of Saline Sol-Iletsk lakes (Orenburg Oblast, Russia). Microbiology 2018, 87, 569–582. [Google Scholar] [CrossRef]
- Shurigin, V.; Hakobyan, A.; Panosyan, H.; Egamberdieva, D.; Davranov, K.; Birkeland, N.K. A glimpse of the prokaryotic diversity of the Large Aral Sea reveals novel extremophilic bacterial and archaeal groups. Microbiol. Open 2019, 8, e00850. [Google Scholar] [CrossRef]
- Skorupa, D.J.; Akyel, A.; Fields, M.W.; Gerlach, R. Facultative and anaerobic consortia of haloalkaliphilic ureolytic micro-organisms capable of precipitating calcium carbonate. J. Appl. Microbiol. 2019, 127, 1479–1489. [Google Scholar] [CrossRef]
- Çelik, P.A.; Mutlu, M.B.; Korkmaz, F.; Yaman, B.N.; Gedikli, S.; Çabuk, D.D.A. Boron mine ponds: Metagenomic insight to bacterial diversity. Biol. Divers. Conserv. 2021, 14, 229–235. [Google Scholar] [CrossRef]
- Quiroz, M.; Triadó-Margarit, X.; Casamayor, E.O.; Gajardo, G. Comparison of Artemia–bacteria associations in brines, laboratory cultures and the gut environment: A study based on Chilean hypersaline environments. Extremophiles 2015, 19, 135–147. [Google Scholar] [CrossRef]
- Xue, M.; Liang, H.; He, Y.; Wen, C. Characterization and in-vivo evaluation of potential probiotics of the bacterial flora within the water column of a healthy shrimp larviculture system. Chin. J. Oceanol. Limnol. 2016, 34, 484–491. [Google Scholar] [CrossRef]
- Hozumi, A.; Hong, P.Y.; Kaartvedt, S.; Røstad, A.; Jones, B.H. Water quality, seasonality, and trajectory of an aquaculture-wastewater plume in the Red Sea. Aquac. Environ. Interac. 2018, 10, 61–77. [Google Scholar] [CrossRef]
- Gorrasi, S.; Pasqualetti, M.; Franzetti, A.; Gonzalez-Martinez, A.; Gonzalez-Lopez, J.; Muñoz-Palazon, B.; Fenice, M. Persistence of Enterobacteriaceae drawn into a marine saltern (Saline di Tarquinia, Italy) from the adjacent coastal zone. Water 2021, 13, 1443. [Google Scholar] [CrossRef]
- Gorrasi, S.; Franzetti, A.; Ambrosini, R.; Pittino, F.; Pasqualetti, M.; Fenice, M. Spatio-temporal variation of the bacterial communities along a salinity gradient within a thalassohaline environment (Saline di Tarquinia salterns, Italy). Molecules 2021, 26, 1338. [Google Scholar] [CrossRef]
- Barghini, P.; Pasqualetti, M.; Gorrasi, S.; Fenice, M. Bacteria from the “Saline di Tarquinia” marine salterns reveal very atypical growth profiles with regards to salinity and temperature. Mediterr. Mar. Sci. 2018, 19, 513–525. [Google Scholar] [CrossRef]
- Gorrasi, S.; Pasqualetti, M.; Franzetti, A.; Pittino, F.; Fenice, M. Vibrio communities along a salinity gradient within a marine saltern hypersaline environment (Saline di Tarquinia, Italy). Environ. Microbiol. 2020, 22, 4356–4366. [Google Scholar] [CrossRef]
- Alfinito, S.; Iberite, M.; Fumanti, B. The algal microflora of the salt works of Tarquinia (Italy). Hydrobiology 1990, 203, 137–146. [Google Scholar] [CrossRef]
- Pasqualetti, M.; Bernini, R.; Carletti, L.; Crisante, F.; Tempesta, S. Salinity and nitrate concentration on the growth and carotenoids accumulation in a strain of Dunaliella salina (Chlorophyta) cultivated under laboratory conditions. Transit. Water. Bull. 2010, 4, 94–104. [Google Scholar] [CrossRef]
- Tempesta, S.; Paoletti, M.; Pasqualetti, M. Morphological and molecular identification of a strain of the unicellular green alga Dunaliella sp. isolated from Tarquinia Salterns. Transit. Water. Bull. 2010, 4, 60–70. [Google Scholar] [CrossRef]
- Barghini, P.; Giovannini, V.; Fenice, M.; Gorrasi, S.; Pasqualetti, M. High lutein production by a halo-tolerant strain of Dunaliella sp. (chlorophyceae) isolated from solar salterns in central Italy. J. Environ. Prot. Ecol. 2018, 19, 704–712. [Google Scholar]
- Barghini, P.; Silvi, S.; Aquilanti, A.; Marcelli, M.; Fenice, M. Bacteria from marine salterns as a model of microorganisms adapted to high environmental variations. J. Environ. Prot. Ecol. 2014, 15, 897–906. [Google Scholar]
- Barghini, P.; Pasqualetti, M.; Gorrasi, S.; Fenice, M. Study of bacterial diversity of a saltern crystallisation pond (“Saline di Tarquinia”, Italy) and its correlation with salinity variations. J. Environ. Prot. Ecol. 2018, 19, 139–145. [Google Scholar]
- Gorrasi, S.; Pesciaroli, C.; Barghini, P.; Pasqualetti, M.; Fenice, M. Structure and diversity of the bacterial community of an Arctic estuarine system (Kandalaksha Bay) subject to intense tidal currents. J. Mar. Syst. 2019, 196, 77–85. [Google Scholar] [CrossRef]
- Gorrasi, S.; Pesciaroli, C.; Barghini, P.; Pasqualetti, M.; Giovannini, V.; Fenice, M. Metagenetic profiling of the bacterial communities of an intertidal pool in Kandalaksha Bay (White Sea, Russia). J. Environ. Prot. Ecol. 2019, 20, 1317–1324. [Google Scholar]
- Rice, E.W.; Baird, R.B.; Eaton, A.D.; Clesceri, L.S. Standard Methods for the Examination of Water and Wastewater, 22nd ed.; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- Centro Funzionale Regionale, Regione Lazio, Annali. Available online: http://www.idrografico.regione.lazio.it/annali/index.htm (accessed on 1 March 2021).
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Claesson, M.J.; O’Sullivan, O.; Wang, Q.; Nikkilä, J.; Marchesi, J.R.; Smidt, H.; de Vos, W.M.; Ross, R.P.; O’Toole, P.W. Comparative analysis of pyrosequencing and a phylogenetic microarray for exploring microbial community structures in the human distal intestine. PLoS ONE 2009, 4, e6669. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Verhoeven, K.J.F.; Simonsen, K.L.; McIntyre, L. Implementing false discovery rate control: Increasing your power. Oikos 2005, 108, 643–647. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Explains % | Contribution % | F | P | P (adj) 1 |
|---|---|---|---|---|---|
| Sampling Site.S | 29.4 | 50.0 | 39.2 | 0.00094 | 0.00189 |
| Sampling Site.P5 | 15.7 | 26.7 | 26.7 | 0.00094 | 0.00113 |
| Sampling Site.P37 | 7.8 | 13.2 | 15.2 | 0.00094 | 0.00113 |
| Sampling Site.P24 | 7.8 | 13.2 | 15.2 | 0.00094 | 0.00113 |
| Salinity | 2.8 | 4.8 | 5.8 | 0.00754 | 0.00905 |
| BOD5 | 1.6 | 2.7 | 3.3 | 0.03110 | 0.03110 |
| sin(Month) | 1.5 | 2.6 | 3.3 | 0.03770 | 0.03770 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorrasi, S.; Pasqualetti, M.; Braconcini, M.; Muñoz-Palazon, B.; Fenice, M. Could Pontimonas Harbour Halophilic Members Able to Withstand Very Broad Salinity Variations? Microorganisms 2022, 10, 790. https://doi.org/10.3390/microorganisms10040790
Gorrasi S, Pasqualetti M, Braconcini M, Muñoz-Palazon B, Fenice M. Could Pontimonas Harbour Halophilic Members Able to Withstand Very Broad Salinity Variations? Microorganisms. 2022; 10(4):790. https://doi.org/10.3390/microorganisms10040790
Chicago/Turabian StyleGorrasi, Susanna, Marcella Pasqualetti, Martina Braconcini, Barbara Muñoz-Palazon, and Massimiliano Fenice. 2022. "Could Pontimonas Harbour Halophilic Members Able to Withstand Very Broad Salinity Variations?" Microorganisms 10, no. 4: 790. https://doi.org/10.3390/microorganisms10040790
APA StyleGorrasi, S., Pasqualetti, M., Braconcini, M., Muñoz-Palazon, B., & Fenice, M. (2022). Could Pontimonas Harbour Halophilic Members Able to Withstand Very Broad Salinity Variations? Microorganisms, 10(4), 790. https://doi.org/10.3390/microorganisms10040790