
Diffusional Interactions among Marine Phytoplankton and Bacterioplankton: Modelling H2O2 as a Case Study


Abstract
:1. Introduction
1.1. Marine Phytoplankton
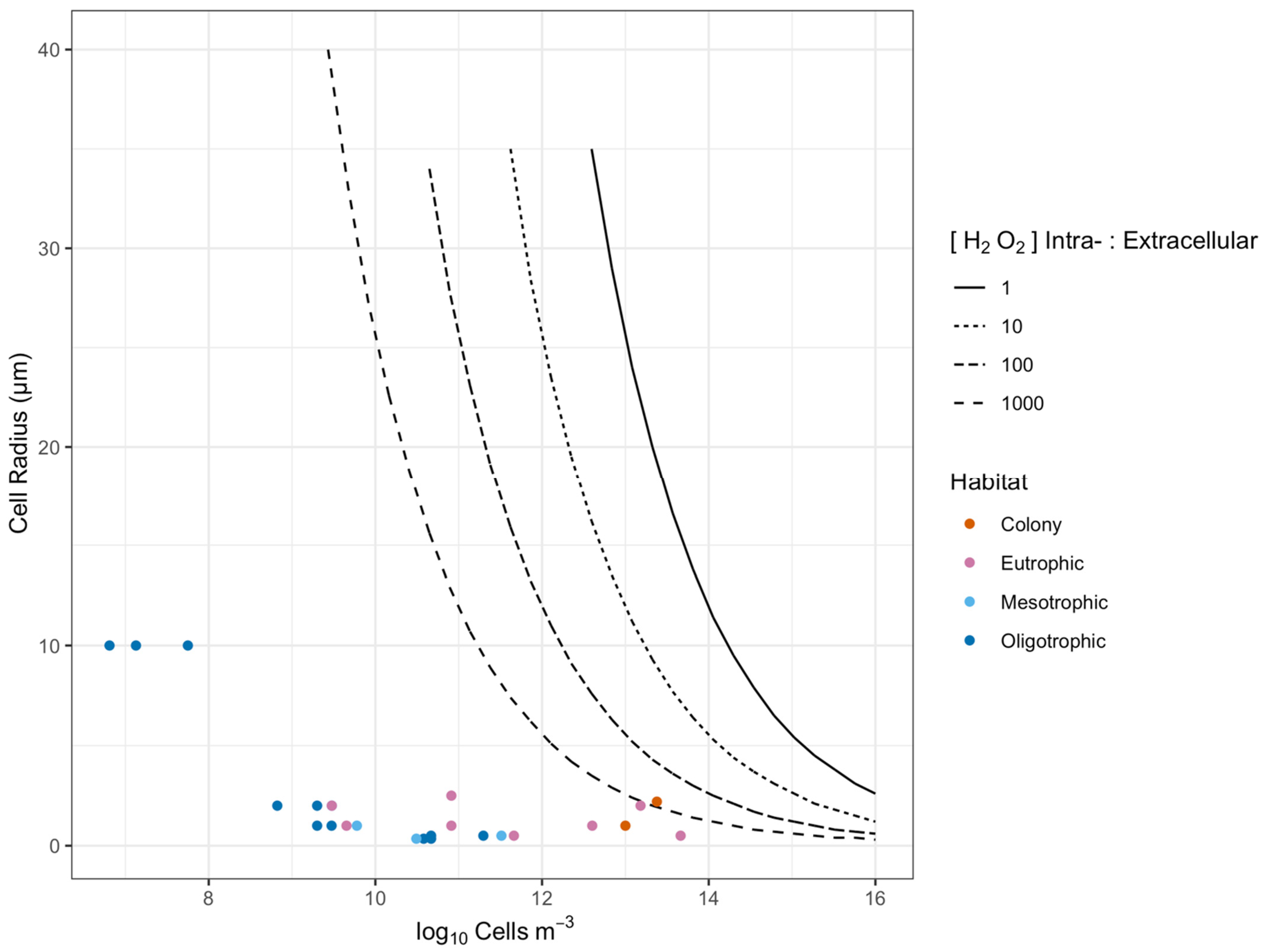
1.2. Phytoplankton Cell Spacing in Oceans

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitat | Taxa | Metabolism | Cell m−3 | Cell Radius µm | Citation |
|---|---|---|---|---|---|
| Oligotrophic | Bacterioplankton | Hetero | 4.7 × 1010 | 0.5 | [9] |
| Oligotrophic | Bacterioplankton | Hetero | 2.0 × 1011 | 0.5 | [10] |
| Oligotrophic | Diatoms | Phyto | 5.6 × 107 | 10.0 | [11] |
| Oligotrophic | Dinoflagellates | Phyto | 6.5 × 106 | 10.0 | [11] |
| Oligotrophic | PicoEukaryotes | Phyto | 6.7 × 108 | 2.0 | [12] |
| Oligotrophic | PicoEukaryotes | Phyto | 2.0 × 109 | 2.0 | [10] |
| Oligotrophic | Prochlorococcus | Phyto | 4.7 × 1010 | 0.3 | [12] |
| Oligotrophic | Prochlorococcus | Phyto | 3.8 × 1010 | 0.3 | [10] |
| Oligotrophic | Silicoflagellates | Phyto | 1.3 × 107 | 10.0 | [11] |
| Oligotrophic | Synechococcus | Phyto | 3.0 × 109 | 1.0 | [12] |
| Oligotrophic | Synechococcus | Phyto | 2.0 × 109 | 1.0 | [10] |
| Mesotrophic | Bacterioplankton | Hetero | 3.3 × 1011 | 0.5 | [10] |
| Mesotrophic | PicoEukaryotes | Phyto | 3.0 × 109 | 2.0 | [10] |
| Mesotrophic | Prochlorococcus | Phyto | 3.1 × 1010 | 0.3 | [10] |
| Mesotrophic | Synechococcus | Phyto | 6.0 × 109 | 1.0 | [10] |
| Eutrophic | Bacterioplankton | Hetero | 4.6 × 1011 | 0.5 | [10] |
| Eutrophic | Bacterioplankton | Hetero | 4.6 × 1013 | 0.5 | [13] |
| Eutrophic | Chlorophytes | Phyto | 8.2 × 1010 | 2.5 | [14] |
| Eutrophic | Chroococcales | Phyto | 8.2 × 1010 | 1.0 | [14] |
| Eutrophic | Cyanobium | Phyto | 4.0 × 1012 | 1.0 | [14] |
| Eutrophic | PicoEukaryotes | Phyto | 3.0 × 109 | 2.0 | [10] |
| Eutrophic | PicoEukaryotes | Phyto | 1.5 × 1013 | 2.0 | [13] |
| Eutrophic | Synechococcus | Phyto | 4.5 × 109 | 1.0 | [10] |
| Colony | Cyanobium | Phyto | 1.0 × 1013 | 1.0 | [14] |
| Colony | Phaeocystis | Phyto | 2.4 × 1013 | 2.2 | [8] |
1.3. Reactive Oxygen Species
1.4. Concentrations and Properties of H2O2
1.5. Goals
2. Results and Discussion
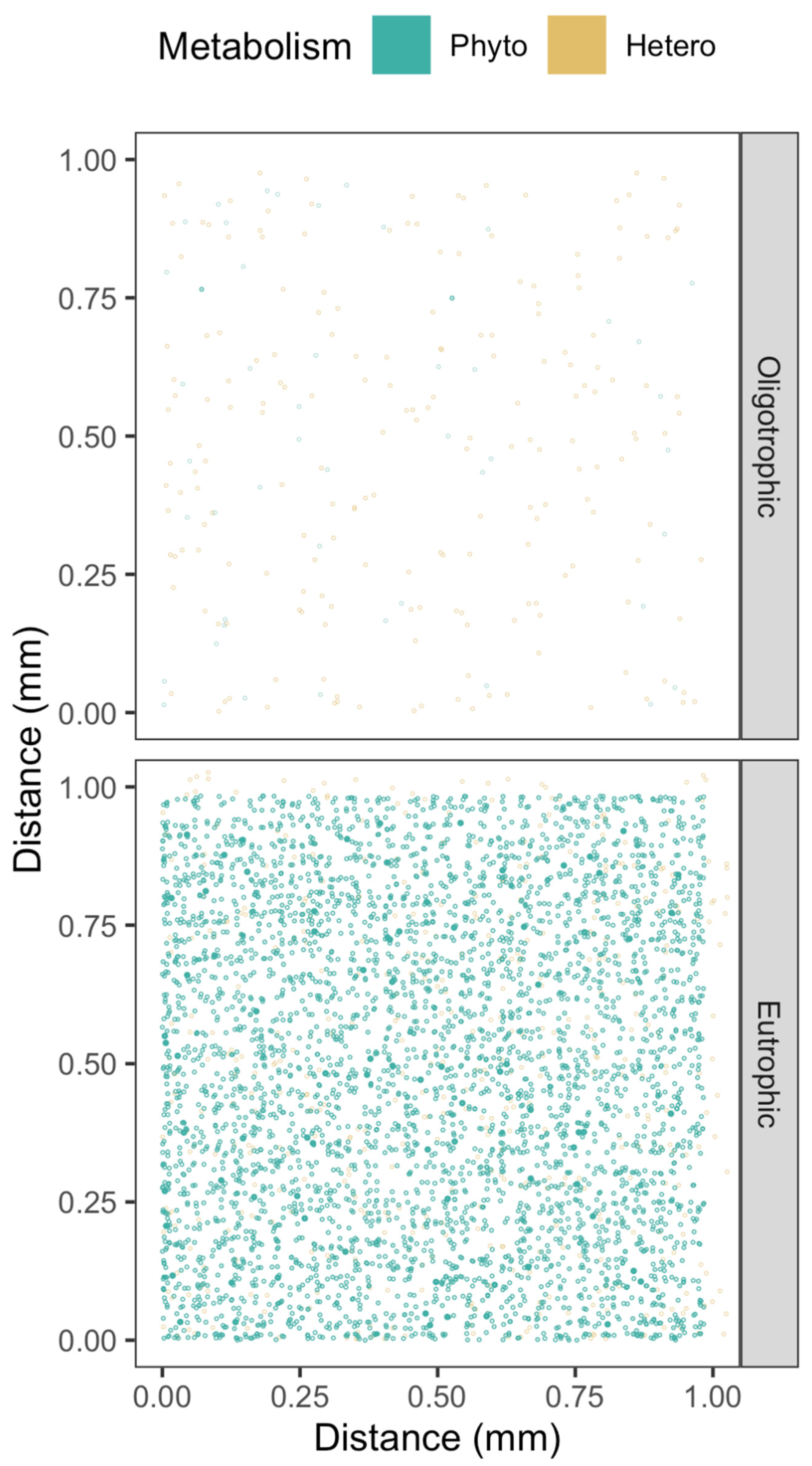
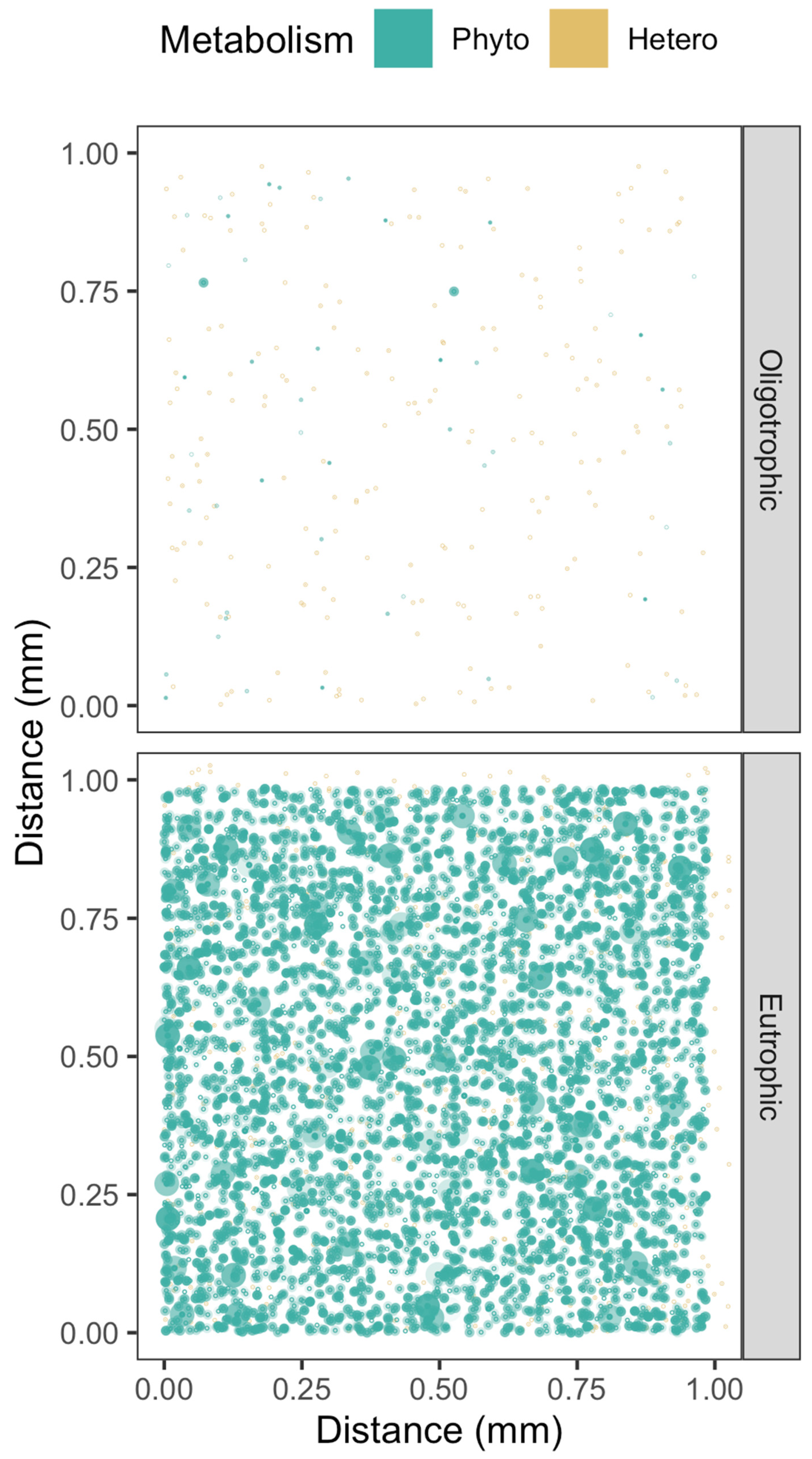
2.1. Cell to Cell Spacing across Habitats and Taxa
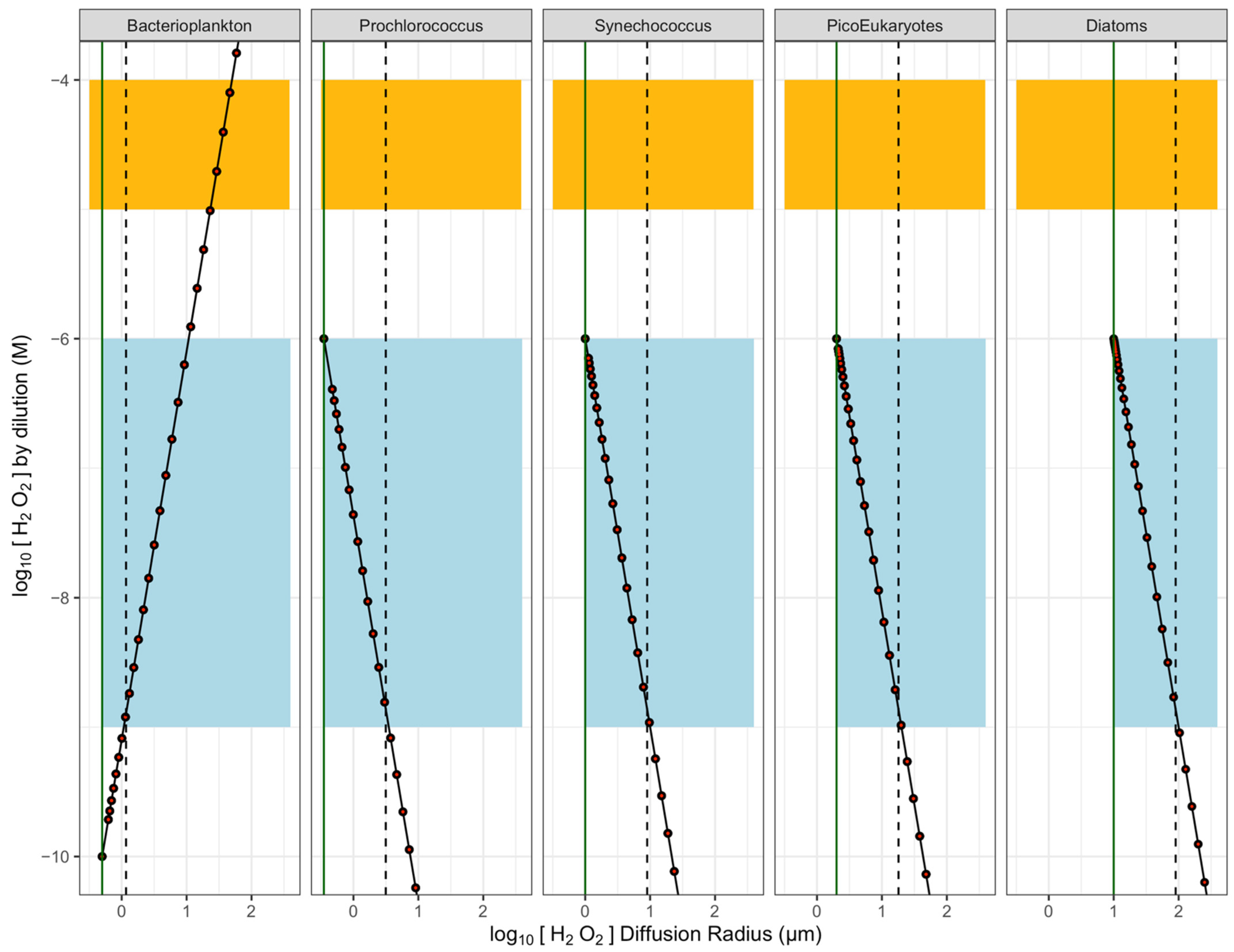
2.2. H2O2 Concentration Gradients around Cells
2.3. Visualizing Cell to Cell Exchange of [H2O2]
2.4. Threshold Distances for Cell to Cell Exchange of [H2O2]
3. Conclusions
4. Methods, Simplifying Assumptions, and Limitations
4.1. Estimation of Cell to Cell Spacing
4.2. Simulating Concentration Gradients
4.3. Simulating Threshold Radii for [H2O2] above or below Seawater [H2O2]
4.4. Simulations and Plotting
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reynolds, C.S. The Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006; ISBN 978-1-139-45489-6. [Google Scholar]
- Falkowski, P.G.; Katz, M.E.; Knoll, A.H.; Quigg, A.; Raven, J.A.; Schofield, O.; Taylor, F.J.R. The Evolution of Modern Eukaryotic Phytoplankton. Sci. Wash. 2004, 305, 354–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, E.M. Life at Low Reynolds Number. Am. J. Phys. 1976, 45, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Finkel, Z.V.; Beardall, J.; Flynn, K.J.; Quigg, A.; Rees, T.A.V.; Raven, J.A. Phytoplankton in a Changing World: Cell Size and Elemental Stoichiometry. J. Plankton Res. 2010, 32, 119–137. [Google Scholar] [CrossRef] [Green Version]
- Stolte, W.; Riegman, R. Effect of Phytoplankton Cell Size on Transient-State Nitrate and Ammonium Uptake Kinetics. Microbiology 1995, 141, 1221–1229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gemmell, B.J.; Oh, G.; Buskey, E.J.; Villareal, T.A. Dynamic Sinking Behaviour in Marine Phytoplankton: Rapid Changes in Buoyancy May Aid in Nutrient Uptake. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161126. [Google Scholar] [CrossRef]
- Bertrand, E.M.; McCrow, J.P.; Moustafa, A.; Zheng, H.; McQuaid, J.B.; Delmont, T.O.; Post, A.F.; Sipler, R.E.; Spackeen, J.L.; Xu, K.; et al. Phytoplanktonbacterial Interactions Mediate Micronutrient Colimitation at the Coastal Antarctic Sea Ice Edge. Proc. Natl. Acad. Sci. USA 2015, 112, 9938–9943. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, H.; Tang, K. Effects of Protozoan Grazing on Colony Formation in Phaeocystis Globosa (Prymnesiophyceae) and the Potential Costs and Benefits. Aquat. Microb. Ecol. 2002, 27, 261–273. [Google Scholar] [CrossRef] [Green Version]
- Robarts, R.; Zohary, T.; Waiser, M.; Yacobi, Y. Bacterial Abundance, Biomass, and Production in Relation to Phytoplankton Biomass in the Levantine Basin of the Southeastern Mediterranean Sea. Mar. Ecol. Prog. Ser. 1996, 137, 273–281. [Google Scholar] [CrossRef]
- Grob, C.; Ulloa, O.; Li, W.; Alarcón, G.; Fukasawa, M.; Watanabe, S. Picoplankton Abundance and Biomass Across the Eastern South Pacific Ocean Along Latitude 32.5. Mar. Ecol. Prog. Ser. 2007, 332, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Olguín, H.F.; Boltovskoy, D.; Lange, C.B.; Brandini, F.; Flynn, K.J. Distribution of Spring Phytoplankton (Mainly Diatoms) in the Upper 50 m of the Southwestern Atlantic Ocean (30). J. Plankton Res. 2006, 28, 1107–1128. [Google Scholar] [CrossRef] [Green Version]
- Agustí, S.; Llabrés, M. Solar Radiation-induced Mortality of Marine Pico-phytoplankton in the Oligotrophic Ocean. Photochem. Photobiol. 2007, 83, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Klinkenberg, G.; Schumann, R. Abundance Changes of Autotrophic and Heterotrophic Picoplankton in the Zingster Strom, a Shallow, Tideless Estuary South of the Darß-Zingst Peninsula (Southern Baltic Sea). Arch. Hydrobiol. 1995, 134, 359–377. [Google Scholar] [CrossRef]
- Albrecht, M.; Pröschold, T.; Schumann, R. Identification of Cyanobacteria in a Eutrophic Coastal Lagoon on the Southern Baltic Coast. Front. Microbiol. 2017, 8, 923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pintoa, E.; Van Nieuwerburgha, L.; De Barros, M.P.; Pedersén, M.; Colepicolo, P.; Snoeijs, P. Density-Dependent Patterns of Thiamine and Pigment Production in the Diatom Nitzschia Microcephala. Phytochemistry 2003, 63, 155–163. [Google Scholar] [CrossRef]
- Fogel, M.L.; Cifuentes, L.A.; Velinsky, D.J.; Sharp, J.H. Relationship of Carbon Availability in Estuarine Phytoplankton to Isotopic Composition. Mar. Ecol. Prog. Ser. 1992, 82, 291–301. [Google Scholar] [CrossRef]
- Hansen, P.J.; Lundholm, N.; Rost, B. Growth Limitation in Marine Red-Tide Dinoflagellates: Efects of pH Versus Inorganic Carbon Availability. Mar. Ecol. Prog. Ser. 2007, 334, 10. [Google Scholar] [CrossRef] [Green Version]
- Avery, G.B.; Cooper, W.J.; Kieber, R.J.; Willey, J.D. Hydrogen Peroxide at the Bermuda Atlantic Time Series Station: Temporal Variability of Seawater Hydrogen Peroxide. Mar. Chem. 2005, 97, 236–244. [Google Scholar] [CrossRef]
- Kearns, K.D.; Hunter, M.D. Algal Extracellular Products Suppress Anabaena Flos-Aquae Heterocyst Spacing. Microb. Ecol. 2002, 43, 174–180. [Google Scholar] [CrossRef]
- Biller, S.J.; Berube, P.M.; Lindell, D.; Chisholm, S.W. Prochlorococcus: The Structure and Function of Collective Diversity. Nat. Rev. Microbiol. 2015, 13, 13–27. [Google Scholar] [CrossRef]
- Kozuleva, M.A.; Ivanov, B.N.; Vetoshkina, D.V.; Borisova-Mubarakshina, M.M. Minimizing an Electron Flow to Molecular Oxygen in Photosynthetic Electron Transfer Chain: An Evolutionary View. Front. Plant Sci. 2020, 11, 211. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants Under Stressful Conditions. J. Bot. 2012, 2012, e217037. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Jia, Z.; Trush, M.A. Defining ROS in Biology and Medicine. React. Oxyg. Species (Apex) 2016, 1, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radi, R. Oxygen Radicals, Nitric Oxide, and Peroxynitrite: Redox Pathways in Molecular Medicine. Proc. Natl. Acad. Sci. USA 2018, 115, 5839–5848. [Google Scholar] [CrossRef] [Green Version]
- Fridovich, I. Oxygen Toxicity: A Radical Explanation. J. Exp. Biol. 1998, 201, 1203–1209. [Google Scholar] [CrossRef]
- Zinser, E.R. The Microbial Contribution to Reactive Oxygen Species Dynamics in Marine Ecosystems. Environ. Microbiol. Rep. 2018, 10, 412–427. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Hansel, C.M.; Voelker, B.M.; Lamborg, C.H. Extensive Dark Biological Production of Reactive Oxygen Species in Brackish and Freshwater Ponds. Environ. Sci. Technol. 2016, 50, 2983–2993. [Google Scholar] [CrossRef] [PubMed]
- Diaz, J.M.; Hansel, C.M.; Voelker, B.M.; Mendes, C.M.; Andeer, P.F.; Zhang, T. Widespread Production of Extracellular Superoxide by Heterotrophic Bacteria. Science 2013, 340, 1223–1226. [Google Scholar] [CrossRef] [PubMed]
- Diaz, J.M.; Plummer, S. Production of Extracellular Reactive Oxygen Species by Phytoplankton: Past and Future Directions. J. Plankton Res. 2018, 40, 655–666. [Google Scholar] [CrossRef] [Green Version]
- Diaz, J.M.; Plummer, S.; Tomas, C.; Alves-de-Souza, C. Production of Extracellular Superoxide and Hydrogen Peroxide by Five Marine Species of Harmful Bloom-Forming Algae. J. Plankton Res. 2018, 40, 667–677. [Google Scholar] [CrossRef] [Green Version]
- Rose, A.L.; Webb, E.A.; Waite, T.D.; Moffett, J.W. Measurement and Implications of Nonphotochemically Generated Superoxide in the Equatorial Pacific Ocean. Environ. Sci. Technol. 2008, 42, 2387–2393. [Google Scholar] [CrossRef]
- Schneider, R.J.; Roe, K.L.; Hansel, C.M.; Voelker, B.M. Species-Level Variability in Extracellular Production Rates of Reactive Oxygen Species by Diatoms. Front. Chem. 2016, 4, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, K.M.; Coe, A.; Gast, R.J.; Plummer, S.; Suffridge, C.P.; Diaz, J.M.; Bowman, J.S.; Wankel, S.D.; Hansel, C.M. Extracellular Superoxide Production by Key Microbes in the Global Ocean. Limnol. Oceanogr. 2019, 64, 2679–2693. [Google Scholar] [CrossRef] [Green Version]
- Kustka, A.B.; Shaked, Y.; Milligan, A.J.; King, D.W.; Morel, F.M.M. Extracellular Production of Superoxide by Marine Diatoms: Contrasting Effects on Iron Redox Chemistry and Bioavailability. Limnol. Oceanogr. 2005, 50, 1172–1180. [Google Scholar] [CrossRef]
- Hansel, C.M.; Buchwald, C.; Diaz, J.M.; Ossolinski, J.E.; Dyhrman, S.T.; Mooy, B.A.S.V.; Polyviou, D. Dynamics of Extracellular Superoxide Production by Trichodesmium Colonies from the Sargasso Sea. Limnol. Oceanogr. 2016, 61, 1188–1200. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; Breusegem, F.V. ROS Signaling: The New Wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef]
- Rhee, S.G. H2O2, a Necessary Evil for Cell Signaling. Science 2006, 312, 1882–1883. [Google Scholar] [CrossRef]
- del Río, L.A.; López-Huertas, E. ROS Generation in Peroxisomes and Its Role in Cell Signaling. Plant Cell Physiol. 2016, 57, 1364–1376. [Google Scholar] [CrossRef]
- del Río, L.A.; Puppo, A. (Eds.) Reactive Oxygen Species in Plant Signaling, Signaling and Communication in Plants; Springer: Dordrecht, The Netherlands; New York, NY, USA, 2009; ISBN 978-3-642-00389-9/978-3-642-00390-5. [Google Scholar]
- Foyer, C.H.; Noctor, G. Redox Homeostasis and Antioxidant Signaling: A Metabolic Interface Between Stress Perception and Physiological Responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, F.-J.; Renger, G.; Friedrich, T.; Kreslavski, V.D.; Zharmukhamedov, S.K.; Los, D.A.; Kuznetsov, V.V.; Allakhverdiev, S.I. Reactive Oxygen Species: Re-evaluation of Generation, Monitoring and Role in Stress-Signaling in Phototrophic Organisms. Biochim. Biophys. Acta (BBA) Bioenerg. 2014, 1837, 835–848. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive Oxygen Species Homeostasis and Signalling During Drought and Salinity Stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Shi, K.; Gao, Z.; Shi, T.-Q.; Song, P.; Ren, L.-J.; Huang, H.; Ji, X.-J. Reactive Oxygen Species-Mediated Cellular Stress Response and Lipid Accumulation in Oleaginous Microorganisms: The State of the Art and Future Perspectives. Front. Microbiol. 2017, 8, 793. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, R. Kinetics of Biological Hydrogen Peroxide Production. Ph.D. Thesis, Colorado School of Mines, Golden, CO, USA, 2015. [Google Scholar]
- Millero, F.J.; Sharma, V.K.; Karn, B. The Rate of Reduction of Copper(II) with Hydrogen Peroxide in Seawater. Mar. Chem. 1991, 36, 71–83. [Google Scholar] [CrossRef]
- Moffett, J.W.; Zika, R.G. Reaction Kinetics of Hydrogen Peroxide with Copper and Iron in Seawater. Environ. Sci. Technol. 1987, 21, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Kelly, T.J.; Daum, P.H.; Schwartz, S.E. Measurements of Peroxides in Cloudwater and Rain. J. Geophys. Res. Atmos. 1985, 90, 7861–7871. [Google Scholar] [CrossRef]
- Morris, J.J.; Johnson, Z.I.; Wilhelm, S.W.; Zinser, E.R. Diel Regulation of Hydrogen Peroxide Defenses by Open Ocean Microbial Communities. J. Plankton Res. 2016, 38, 1103–1114. [Google Scholar] [CrossRef]
- Yuan, J.; Shiller, A.M. The Distribution of Hydrogen Peroxide in the Southern and Central Atlantic Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2001, 48, 2947–2970. [Google Scholar] [CrossRef]
- Tjell, A.Ø.; Almdal, K. Diffusion Rate of Hydrogen Peroxide Through Water-Swelled Polyurethane Membranes. Sens. Bio-Sens. Res. 2018, 21, 35–39. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative Stress in Marine Environments: Biochemistry and Physiological Ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 3rd ed.; Oxford Science Publications: Oxford, UK; Clarendon Press, Oxford University Press: Oxford, UK, 1999; ISBN 978-0-19-850045-2/978-0-19-850044-5. [Google Scholar]
- González-Flecha, B.; Demple, B. Homeostatic Regulation of Intracellular Hydrogen Peroxide Concentration in Aerobically Growing Escherichia Coli. J. Bacteriol. 1997, 179, 382–388. [Google Scholar] [CrossRef] [Green Version]
- Lyublinskaya, O.; Antunes, F. Measuring Intracellular Concentration of Hydrogen Peroxide with the Use of Genetically Encoded H2O2 Biosensor HyPer. Redox Biol. 2019, 24, 101200. [Google Scholar] [CrossRef] [PubMed]
- Stone, J.R.; Yang, S. Hydrogen Peroxide: A Signaling Messenger. Antioxid. Redox Signal. 2006, 8, 243–270. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Hydrogen Peroxide as a Central Redox Signaling Molecule in Physiological Oxidative Stress: Oxidative Eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef]
- Seaver, L.C.; Imlay, J.A. Hydrogen Peroxide Fluxes and Compartmentalization Inside Growing Escherichia Coli. J. Bacteriol. 2001, 183, 7182–7189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, B.K.; Sikes, H.D. Quantifying Intracellular Hydrogen Peroxide Perturbations in Terms of Concentration. Redox Biol. 2014, 2, 955–962. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Imlay, J.A. Improved Measurements of Scant Hydrogen Peroxide Enable Experiments That Define Its Threshold of Toxicity for Escherichia Coli. Free Radic. Biol. Med. 2018, 120, 217–227. [Google Scholar] [CrossRef]
- Forman, H.J.; Bernardo, A.; Davies, K.J.A. What Is the Concentration of Hydrogen Peroxide in Blood and Plasma? Arch. Biochem. Biophys. 2016, 603, 48–53. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Hampton, M.B. Thiol Chemistry and Specificity in Redox Signaling. Free Radic. Biol. Med. 2008, 45, 549–561. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [Green Version]
- Fedurayev, P.V.; Mironov, K.S.; Gabrielyan, D.A.; Bedbenov, V.S.; Zorina, A.A.; Shumskaya, M.; Los, D.A. Hydrogen Peroxide Participates in Perception and Transduction of Cold Stress Signal in Synechocystis. Plant Cell Physiol. 2018, 59, 1255–1264. [Google Scholar] [CrossRef]
- Khademian, M.; Imlay, J.A. Escherichia Coli Cytochrome c Peroxidase Is a Respiratory Oxidase That Enables the Use of Hydrogen Peroxide as a Terminal Electron Acceptor. Proc. Natl. Acad. Sci. USA 2017, 114, E6922–E6931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bienert, G.P.; Chaumont, F. Aquaporin-Facilitated Transmembrane Diffusion of Hydrogen Peroxide. Biochim. Biophys. Acta 2014, 1840, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Almasalmeh, A.; Krenc, D.; Wu, B.; Beitz, E. Structural Determinants of the Hydrogen Peroxide Permeability of Aquaporins. FEBS J. 2014, 281, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Schoebel, S.; Schmitz, F.; Dong, H.; Hedfalk, K. Characterization of Aquaporin-Driven Hydrogen Peroxide Transport. Biochim. Biophys. Acta (BBA) Biomembr. 2020, 1862, 183065. [Google Scholar] [CrossRef]
- Miller, E.W.; Dickinson, B.C.; Chang, C.J. Aquaporin-3 Mediates Hydrogen Peroxide Uptake to Regulate Downstream Intracellular Signaling. Proc. Natl. Acad. Sci. USA 2010, 107, 15681–15686. [Google Scholar] [CrossRef] [Green Version]
- Bond, R.J.; Hansel, C.M.; Voelker, B.M. Heterotrophic Bacteria Exhibit a Wide Range of Rates of Extracellular Production and Decay of Hydrogen Peroxide. Front. Mar. Sci. 2020, 7, 72. [Google Scholar] [CrossRef] [Green Version]
- Coe, A.; Ghizzoni, J.; LeGault, K.; Biller, S.; Roggensack, S.E.; Chisholm, S.W. Survival of Prochlorococcus in Extended Darkness. Limnol. Oceanogr. 2016, 61, 1375–1388. [Google Scholar] [CrossRef]
- Zinser, E.R. Cross-Protection from Hydrogen Peroxide by Helper Microbes: The Impacts on the Cyanobacterium Prochlorococcus and Other Beneficiaries in Marine Communities. Environ. Microbiol. Rep. 2018, 10, 199–411. [Google Scholar] [CrossRef]
- Zhu, Z.; Meng, R.; Smith, W.O., Jr.; Doan-Nhu, H.; Nguyen-Ngoc, L.; Jiang, X. Bacterial Composition Associated With Giant Colonies of the Harmful Algal Species Phaeocystis Globosa. Front. Microbiol. 2021, 12, 737484. [Google Scholar] [CrossRef]
- Oda, T.; Moritomi, J.; Kawano, I.; Hamaguchi, S.; Ishimatsu, A.; Muramatsu, T. Catalase- and Superoxide Dismutase-induced Morphological Changes and Growth Inhibition in the Red Tide Phytoplankton Chattonella Marina. Biosci. Biotechnol. Biochem. 1995, 59, 2044–2048. [Google Scholar] [CrossRef]
- Morris, J.J.; Rose, A.L.; Lu, Z. Reactive Oxygen Species in the World Ocean and Their Impacts on Marine Ecosystems. Redox Biol. 2022, 52, 102285. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Nakamura, A.; Shikayama, M.; Kawano, I.; Ishimatsu, A.; Muramatsu, T. Generation of Reactive Oxygen Species by Raphidophycean Phytoplankton. Biosci. Biotechnol. Biochem. 1997, 61, 1658–1662. [Google Scholar] [CrossRef] [PubMed]
- Average Separation of Objects within a Volume? 2009. Available online: http://www.forum.cosmoquest.org/showthread.php?93443-Average-separation-of-objects-within-a-volume (accessed on 21 October 2021).
- Lambert W Function. 2022. Available online: https://en.wikipedia.org/wiki/Lambert_W_function (accessed on 19 January 2022).
- RC Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- RStudio Team. RStudio: Integrated Development Environment for R.; RStudio, Inc.: Boston, MA, USA, 2015. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Borchers, H.W. Pracma: Practical Numerical Math Functions. 2021. Available online: https://cran.r-project.org/web/packages/pracma/index.html (accessed on 22 October 2021).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Wilke, C.O. Cowplot: Streamlined Plot Theme and Plot Annotations for ‘Ggplot2’. 2019. Available online: https://cran.r-project.org/web/packages/ggplot2/index.html (accessed on 12 December 2021).
- Kassambara, A. Ggpubr: ‘Ggplot2’ Based Publication Ready Plots. 2018. Available online: https://cran.r-project.org/web/packages/ggpubr/index.html (accessed on 17 July 2019).
- Hester, J. Glue: Interpreted String Literals. 2018. Available online: https://cran.r-project.org/web/packages/glue/index.html (accessed on 17 July 2019).
- Zhu, H. kableExtra: Construct Complex Table with ‘Kable’ and Pipe Syntax. 2019. Available online: https://cran.r-project.org/web/packages/kableExtra/index.html (accessed on 30 July 2019).
- Goerg, G.M. LambertW: Probabilistic Models to Analyze and Gaussianize Heavy-Tailed, Skewed Data. 2020. Available online: https://cran.r-project.org/web/packages/LambertW/index.html (accessed on 16 December 2020).
- Pedersen, T.L. RStudio Ggforce: Accelerating ‘Ggplot2’. 2021. Available online: https://cran.r-project.org/web/packages/ggforce/index.html (accessed on 5 March 2021).
- D’Agostino McGowan, L.; Bryan, J. Googledrive: An Interface to Google Drive. 2020. Available online: https://cran.r-project.org/web/packages/googledrive/index.html (accessed on 21 October 2021).
- Bryan, J. Googlesheets4: Access Google Sheets Using the Sheets API V4. 2021. Available online: https://cran.r-project.org/web/packages/googlesheets4/index.html (accessed on 21 October 2021).
- Zotero|Your Personal Research Assistant. Available online: https://www.zotero.org/ (accessed on 7 December 2021).
- Aust, F. Citr: ’RStudio’ Add-in to Insert Markdown Citations. 2018. Available online: https://github.com/crsh/citr (accessed on 17 July 2019).
- Xie, Y. Knitr: A Comprehensive Tool for Reproducible Research in R. In Implementing Reproducible Computational Research; Stodden, V., Leisch, F., Peng, R.D., Eds.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2014; Available online: https://www.taylorfrancis.com/chapters/edit/10.1201/9781315373461-1/knitr-comprehensive-tool-reproducible-research-yihui-xie (accessed on 30 July 2019).
- Xie, Y. Dynamic Documents with R and Knitr; Chapman and Hall/CRC: Boca Raton, FL, USA, 2015; Available online: https://duhi23.github.io/Analisis-de-datos/Yihue.pdf (accessed on 30 July 2019).
- Xie, Y. Knitr: A General-Purpose Package for Dynamic Report Generation in R; 2018. Available online: https://cran.r-project.org/web/packages/knitr/index.html (accessed on 30 July 2019).
- Xie, Y. Bookdown: Authoring Books and Technical Documents with R Markdown; 2019. Available online: https://cran.r-project.org/web/packages/bookdown/index.html (accessed on 22 November 2021).






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omar, N.M.; Prášil, O.; McCain, J.S.P.; Campbell, D.A. Diffusional Interactions among Marine Phytoplankton and Bacterioplankton: Modelling H2O2 as a Case Study. Microorganisms 2022, 10, 821. https://doi.org/10.3390/microorganisms10040821
Omar NM, Prášil O, McCain JSP, Campbell DA. Diffusional Interactions among Marine Phytoplankton and Bacterioplankton: Modelling H2O2 as a Case Study. Microorganisms. 2022; 10(4):821. https://doi.org/10.3390/microorganisms10040821
Chicago/Turabian StyleOmar, Naaman M., Ondřej Prášil, J. Scott P. McCain, and Douglas A. Campbell. 2022. "Diffusional Interactions among Marine Phytoplankton and Bacterioplankton: Modelling H2O2 as a Case Study" Microorganisms 10, no. 4: 821. https://doi.org/10.3390/microorganisms10040821
APA StyleOmar, N. M., Prášil, O., McCain, J. S. P., & Campbell, D. A. (2022). Diffusional Interactions among Marine Phytoplankton and Bacterioplankton: Modelling H2O2 as a Case Study. Microorganisms, 10(4), 821. https://doi.org/10.3390/microorganisms10040821