
Isolation and Properties of the Bacterial Strain Janthinobacterium sp. SLB01


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of the Janthinobacterium sp. SLB01 Strain
2.2. Pigment Extraction
2.3. Violacein Production
2.4. Biofilm Detection
2.5. Biochemical Characterization of the Strain
2.6. Antibiotics Sensitivity Tests of the Strain
2.7. Statistical Analysis
3. Results
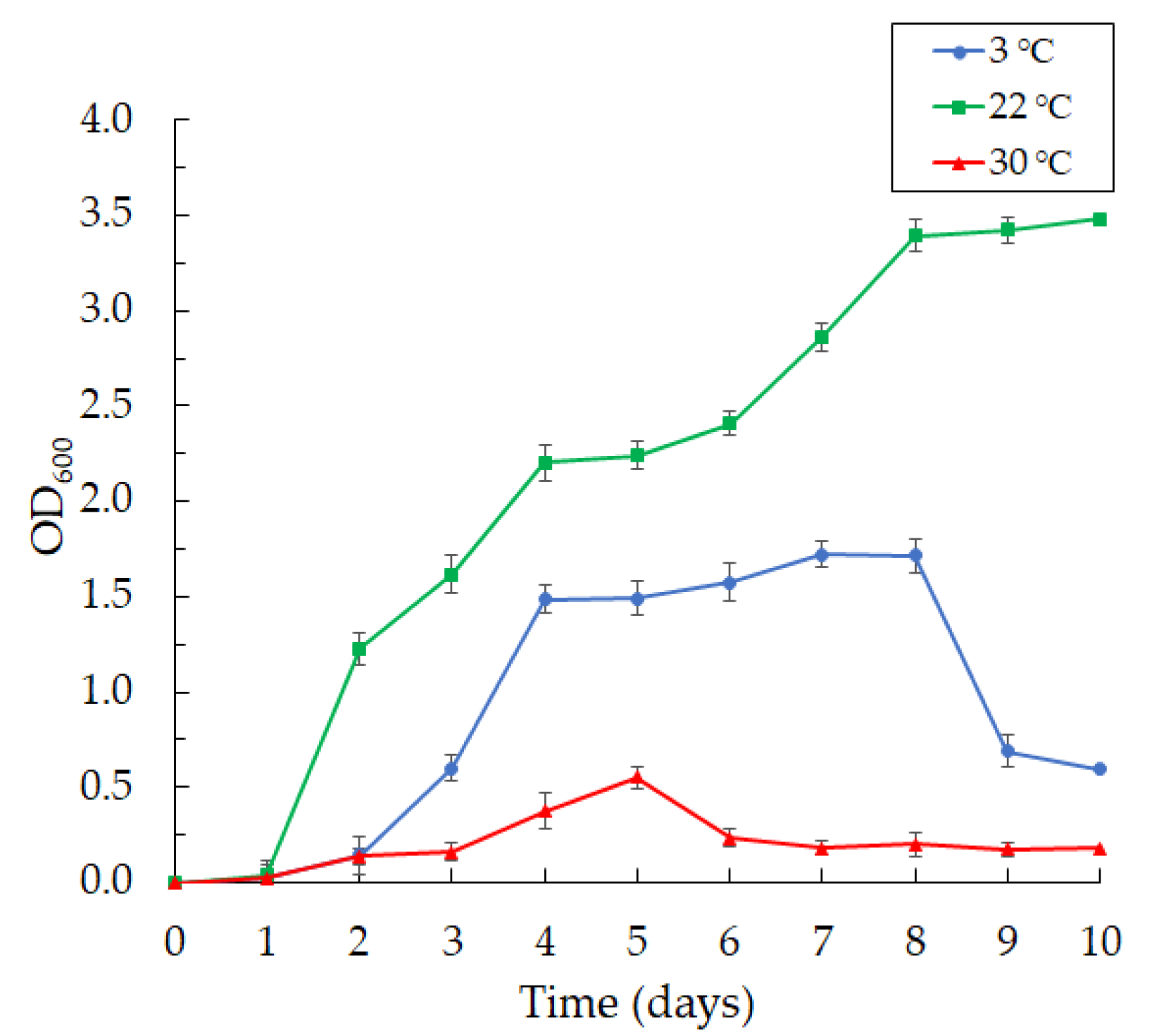
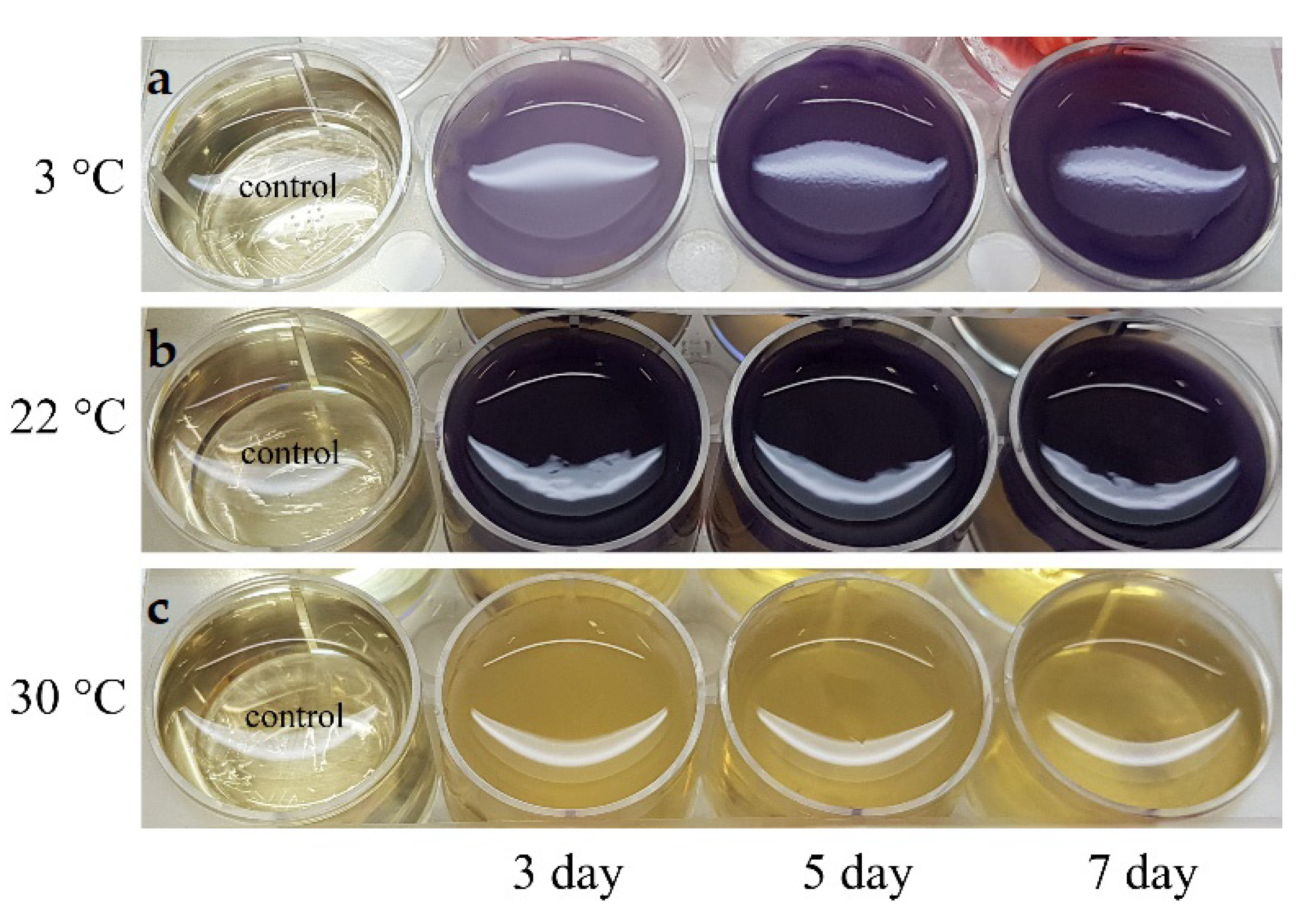
3.1. Isolation of the Bacterial Strains and Culture Conditions
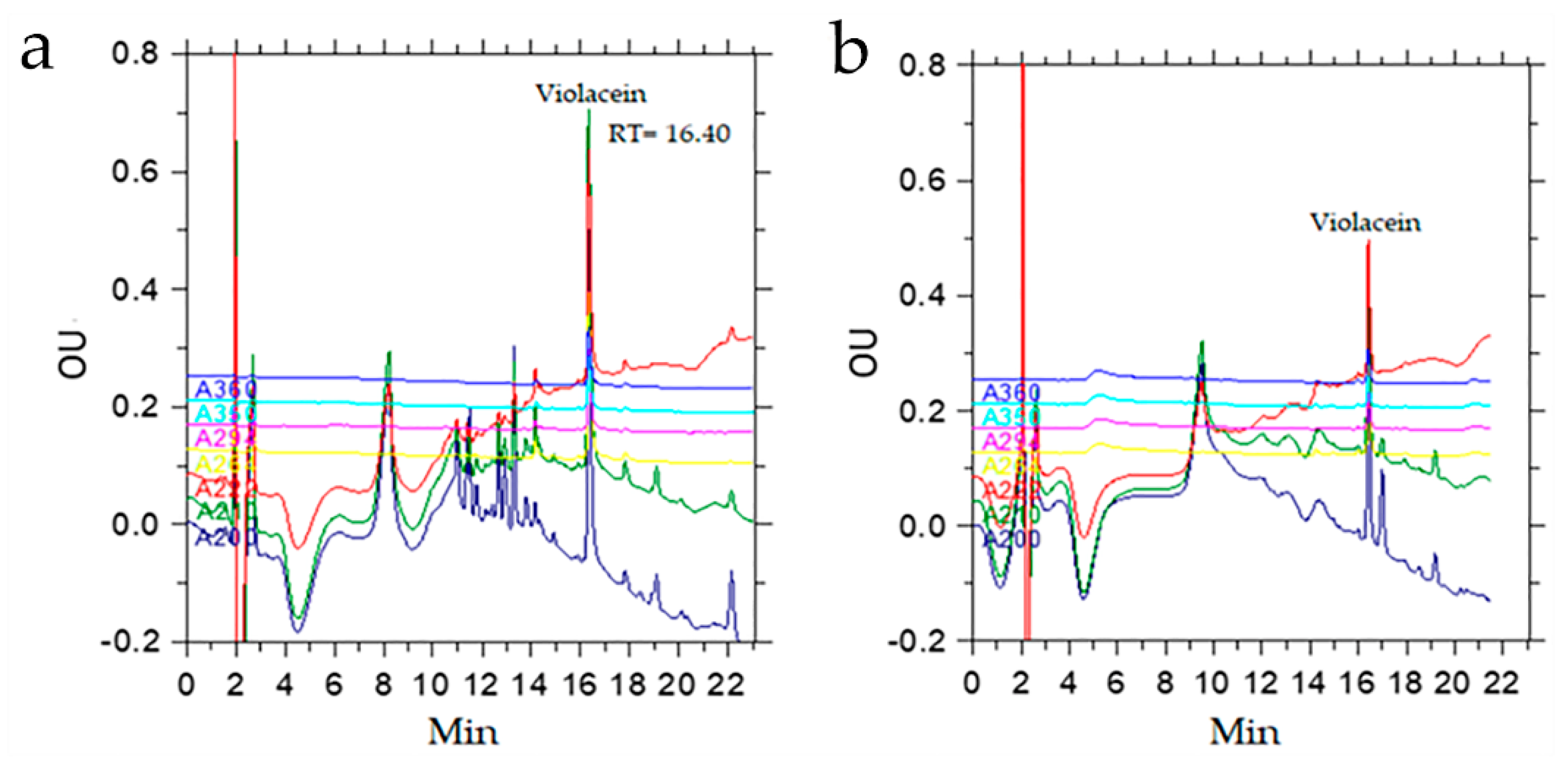
3.2. Characterization of Violacein
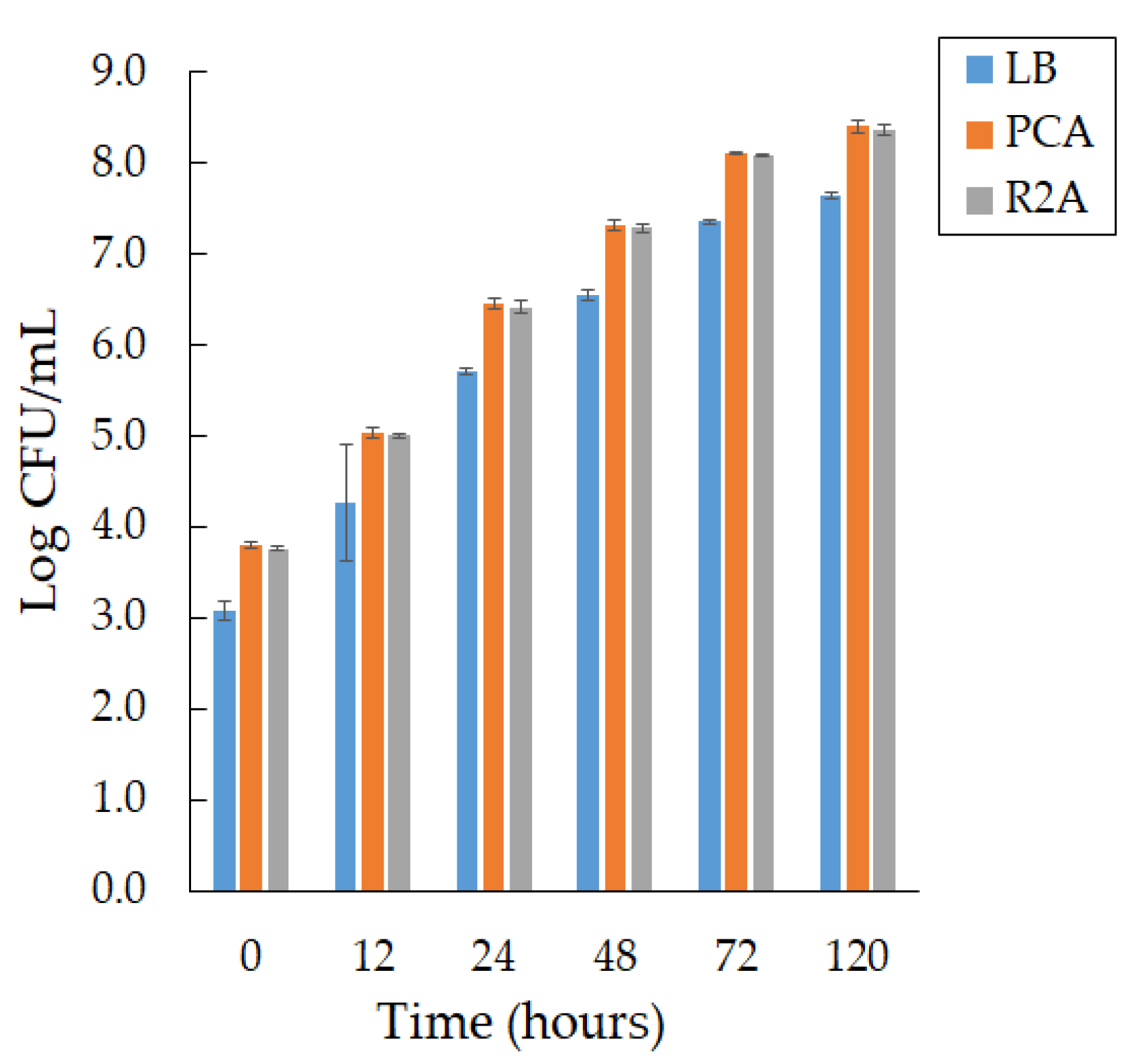
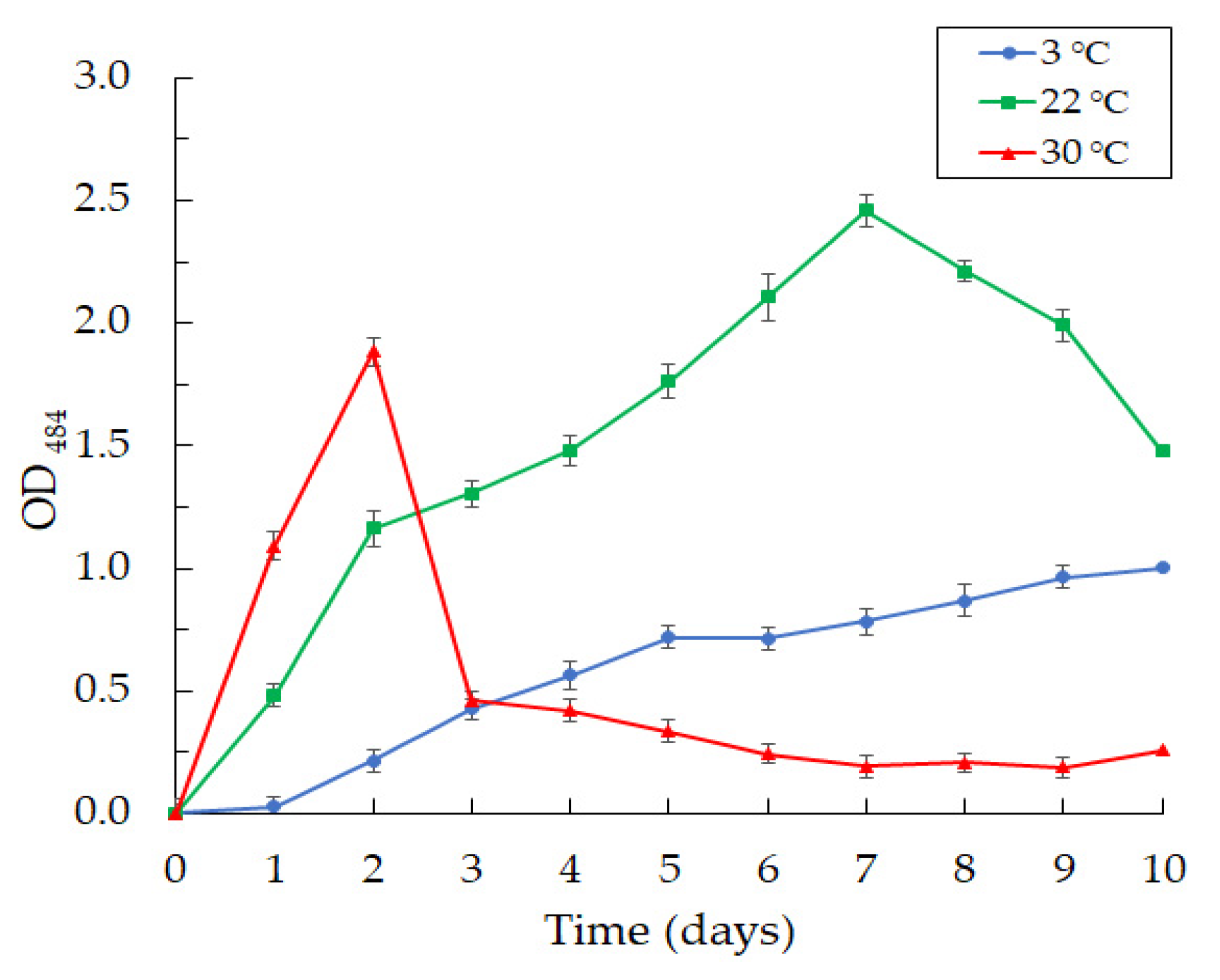
3.3. Production of Violacein
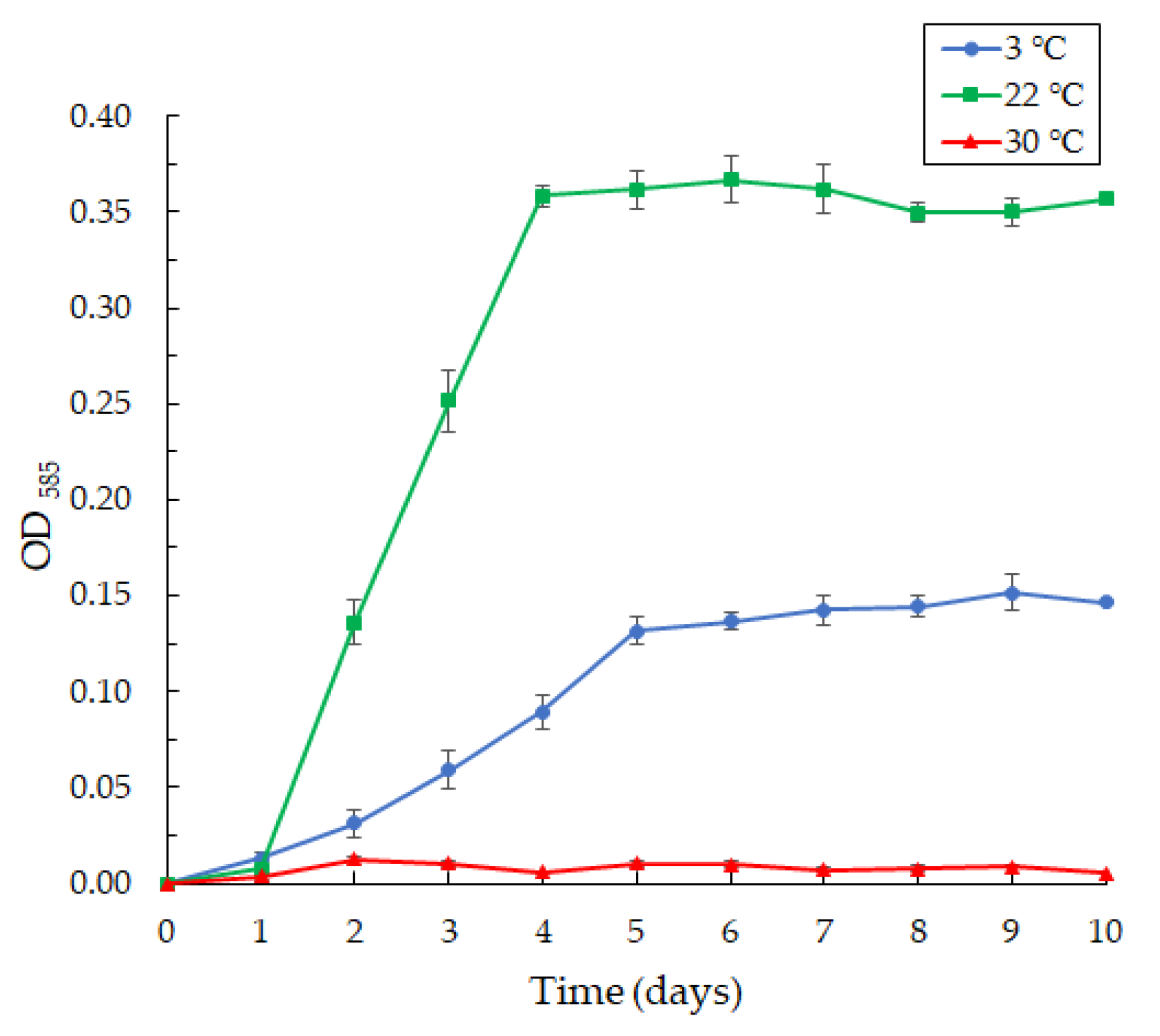
3.4. Biofilm Formation
3.5. Biochemical Analysis
3.6. Antibiotic Sensitivity Tests of the Strain
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garrity, G.M.; Bell, J.A.; Lilburn, T.E. Family II Oxalobacteraceae fam. nov. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Garrity, G.M., Brenner, D.J., Krieg, N.R., Staley, J., Eds.; Springer: New York, NY, USA, 2005; Volume 2, p. 623. [Google Scholar] [CrossRef]
- Coenye, T. The family Burkholderiaceae. In The Prokaryotes: Alphaproteobacteria and Betaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: New York, NY, USA, 2014; pp. 759–776. [Google Scholar] [CrossRef]
- Gillis, M.; Logan, N.A. Janthinobacterium. In Bergey’s Manual of Systematics of Archaea and Bacteria; Trujillo, M.E., Dedysh, S., DeVos, P., Hedlund, B., Kämpfer, P., Rainey, F.A., Whitman, W.B., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA; Bergey’s Manual Trust: Athen, GA, USA, 2015; pp. 1–12. [Google Scholar] [CrossRef]
- De Ley, J.; Segers, P.; Gillis, M. Intra- and intergeneric similarities of Chromobacterium and Janthinobacterium ribosomal ribonucleic acid cistrons. Int. J. Syst. Evol. Microbiol. 1978, 28, 154–168. [Google Scholar] [CrossRef] [Green Version]
- Sneath, P.H.A. Genus Janthinobacterium. In Bergey’s Manual of Systematic Bacteriology; Krieg, N.R., Holt, J.C., Eds.; The Williams & Wilkins Co.: Baltimore, MD, USA, 1984; pp. 376–377. [Google Scholar]
- Schloss, P.D.; Allen, H.K.; Klimowicz, A.K.; Mlot, C.; Gross, J.A.; Savengsuksa, S.; McEllin, J.; Clardy, J.; Ruess, R.W.; Handelsman, J. Psychrotrophic strain of Janthinobacterium lividum from a cold Alaskan soil produces prodigiosin. DNA Cell Biol. 2010, 29, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Skrivergaard, S.; Korsgaard, B.S.; Schreiber, L.; Marshall, I.P.; Finster, K.; Schramm, A. High quality draft genome sequence of Janthinobacterium psychrotolerans sp. nov., isolated from a frozen freshwater pond. Stand. Genom. Sci. 2017, 12, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillis, M.; De Ley, J. The Genera Chromobacterium and Janthinobacterium. In The Prokaryotes: Handbook on the Biology of Bacteria, 2nd ed.; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 7, pp. 737–746. [Google Scholar] [CrossRef]
- Leon, L.L.; Miranda, C.C.; De Souza, A.O.; Durán, N. Antileishmanial activity of the violacein extracted from Chromobacterium violaceum. J. Antimicrob. Chemother. 2001, 48, 449–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrighetti-Fröhner, C.R.; Antonio, R.V.; Creczynski-Pasa, T.B.; Barardi, C.R.; Simões, C.M. Cytotoxicity and potential antiviral evaluation of violacein produced by Chromobacterium violaceum. Mem. Inst. Oswaldo Cruz 2003, 98, 843–848. [Google Scholar] [CrossRef]
- Pantanella, F.; Berlutti, F.; Passariello, C.; Sarli, S.; Morea, C.; Schippa, S. Violacein and biofilm production in Janthinobacterium lividum. J. Appl. Microbiol. 2006, 102, 992–999. [Google Scholar] [CrossRef]
- Konzen, M.; De Marco, D.; Cordova, C.A.; Vieira, T.O.; Antônio, R.V.; Creczynski-Pasa, T.B. Antioxidant properties of violacein: Possible relation on its biological function. Bioorg. Med. Chem. 2006, 14, 8307–8313. [Google Scholar] [CrossRef]
- Durán, N.; Justo, G.Z.; Ferreira, C.V.; Melo, P.S.; Cordi, L.; Martins, D. Violacein: Properties and biological activities. Biotechnol. Appl. Biochem. 2007, 48, 127–133. [Google Scholar] [CrossRef]
- Lopes, S.C.; Blanco, Y.C.; Justo, G.Z.; Nogueira, P.A.; Rodrigues, F.L.; Goelnitz, U.; Wunderlich, G.; Facchini, G.; Brocchi, M.; Duran, N.; et al. Violacein extracted from Chromobacterium violaceum inhibits Plasmodium growth in vitro and in vivo. Antimicrob. Agents Chemother. 2009, 53, 2149–2152. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.L.; Gocke, Y.; Bolten, C.; Brock, N.L.; Dickschat, J.S.; Wittmann, C. Microbial production of the drugs violacein and deoxyviolacein: Analytical development and strain comparison. Biotechnol. Lett. 2012, 34, 717–720. [Google Scholar] [CrossRef]
- Asencio, G.; Lavin, P.; Alegría, K.; Domínguez, M.; Bello, H.; González-Rocha, G.; González-Aravena, M. Antibacterial activity of the Antarctic bacterium Janthinobacterium sp. SMN 33.6 against multi-resistant gram-negative bacteria. Electron. J. Biotechnol. 2014, 17, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Ballestriero, F.; Daim, M.; Penesyan, A.; Nappi, J.; Schleheck, D.; Bazzicalupo, P.; Di Schiavi, E.; Egan, S. Antinematode activity of Violacein and the role of the insulin/IGF-1 pathway in controlling violacein sensitivity in Caenorhabditis elegans. PLoS ONE 2014, 9, e109201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimi, S.M.; Xu, T.; Wei, M.Q. Violacein anticancer activity is enhanced under hypoxia. Oncol. Rep. 2015, 33, 1731–1736. [Google Scholar] [CrossRef] [PubMed]
- Sasidharan, A.; Sasidharan, N.K.; Amma, D.B.; Vasu, R.K.; Nataraja, A.V.; Bhaskaran, K. Antifungal activity of violacein purified from a novel strain of Chromobacterium sp. NIIST (MTCC 5522). J. Microbiol. 2015, 53, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Valdes, N.; Soto, P.; Cottet, L.; Alarcon, P.; Gonzalez, A.; Castillo, A.; Corsini, G.; Tello, M. Draft genome sequence of Janthinobacterium lividum strain MTR reveals its mechanism of capnophilic behavior. Stand. Genom. Sci. 2015, 10, 110. [Google Scholar] [CrossRef] [Green Version]
- Baricz, A.; Teban, A.; Chiriac, C.M.; Szekeres, E.; Farkas, A.; Nica, M.; Dascălu, A.; Oprișan, C.; Lavin, P.; Coman, C. Investigating the potential use of an Antarctic variant of Janthinobacterium lividum for tackling antimicrobial resistance in a One Health approach. Sci. Rep. 2018, 8, 15272. [Google Scholar] [CrossRef] [Green Version]
- Dike-Ndudim, J.N.; Ugenyi, L.C.; Ndubueze, C.W. Assessment of antifungal potentials of violacein extract from Chromobacterium violaceum isolated from domestic and recreational water sources in Owerri, Imo State, Nigeria. World J. Adv. Res. Rev. 2021, 10, 168–172. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yuk, N.; Shin, H.J.; Jung, H.J. The Natural Pigment Violacein Potentially Suppresses the Proliferation and Stemness of Hepatocellular Carcinoma Cells In Vitro. Int. J. Mol. Sci. 2021, 22, 10731. [Google Scholar] [CrossRef]
- Matz, C.; Deines, P.; Boenigk, J.; Arndt, H.; Eberl, L.; Kjelleberg, S.; Jürgens, K. Impact of violacein-producing bacteria on survival and feeding of bacterivorous nanoflagellates. Appl. Environ. Microbiol. 2004, 70, 1593–1599. [Google Scholar] [CrossRef] [Green Version]
- Harris, R.N.; Brucker, R.M.; Walke, J.B.; Becker, M.H.; Schwantes, C.R.; Flaherty, D.C.; Lam, B.A.; Woodhams, D.C.; Briggs, C.J.; Vredenburg, V.T.; et al. Skin microbes on frogs prevent morbidity and mortality caused by a lethal skin fungus. ISME J. 2009, 3, 818–824. [Google Scholar] [CrossRef]
- Jensen, S.; Ovreas, L.; Bergh, O.; Torsvik, V. Phylogenetic analysis of bacterial communities associated with larvae of the Atlantic halibut proposesuccession from a uniform normal flora. Syst. Appl. Microbiol. 2004, 27, 728–736. [Google Scholar] [CrossRef] [PubMed]
- Oh, W.T.; Giri, S.S.; Yun, S.; Kim, H.J.; Kim, S.G.; Kim, S.W.; Kang, J.W.; Han, S.J.; Kwon, J.; Jun, J.W.; et al. Janthinobacterium lividum as An Emerging Pathogenic Bacterium Affecting Rainbow Trout (Oncorhynchus mykiss) Fisheries in Korea. Pathogens 2019, 8, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, B.; Gonzalez, C.J.; Stobie, M.; Curry, J.I.; McLoughlin, M.F. Recovery of Janthinobacterium lividum from diseased rainbow trout, Oncorhynchus mykiss (Walbaum), in Northern Ireland and Scotland. J. Fish Dis. 1992, 15, 357–359. [Google Scholar] [CrossRef]
- Lincoln, S.P.; Fermor, T.R.; Tindall, B.J. Janthinobacterium agaricidamnosum sp. nov., a soft rot pathogen of Agaricusbisporus. Int. J. Syst. Evol. Microbiol. 1999, 49, 1577–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haack, F.S.; Poehlein, A.; Kröger, C.; Voigt, C.A.; Piepenbring, M.; Bode, H.B.; Daniel, R.; Schäfer, W.; Streit, W.R. Molecular keys to the Janthinobacterium and Duganella spp. interaction with the plant pathogen Fusarium graminearum. Front. Microbiol. 2016, 7, 1668. [Google Scholar] [CrossRef] [Green Version]
- Jung, W.J.; Kim, S.W.; Giri, S.S.; Kim, H.J.; Kim, S.G.; Kang, J.W.; Kwon, J.; Lee, S.B.; Oh, W.T.; Jun, J.W.; et al. Janthinobacterium tructae sp. nov., Isolated from Kidney of Rainbow Trout (Oncorhynchus mykiss). Pathogens 2021, 10, 229. [Google Scholar] [CrossRef]
- Petrushin, I.S.; Belikov, S.I.; Chernogor, L.I. Draft Genome Sequence of Janthinobacterium sp. Strain SLB01, Isolated from the Diseased Sponge Lubomirskia baicalensis. Microbiol. Resour. Announc. 2019, 8, e01108–e01119. [Google Scholar] [CrossRef] [Green Version]
- Khanaev, I.V.; Kravtsova, L.S.; Maikova, O.O.; Bukshuk, N.A.; Sakirko, M.V.; Kulakova, N.V.; Butina, T.V.; Nebesnykh, I.A.; Belikov, S.I. Current state of the sponge fauna (Porifera: Lubomirskiidae) of Lake Baikal: Sponge disease and the problem of conservation of diversity. J. Great Lakes Res. 2018, 44, 77–85. [Google Scholar] [CrossRef]
- Belikov, S.; Belkova, N.; Butina, T.; Chernogor, L.; Van Kley, A.M.; Nalian, A.; Rorex, C.; Khanaev, I.; Maikova, O.; Feranchuk, S. Diversity and shifts of the bacterial community associated with Baikal sponge mass mortalities. PLoS ONE 2019, 14, e0213926. [Google Scholar] [CrossRef]
- Chernogor, L.; Klimenko, E.; Khanaev, I.; Belikov, S. Microbiome analysis of healthy and diseased sponges Lubomirskia baicalensis by using cell cultures of primmorphs. PeerJ 2020, 8, e9080. [Google Scholar] [CrossRef]
- Belikov, S.I.; Petrushin, I.S.; Chernogor, L.I. Genome Analysis of the Janthinobacterium sp. Strain SLB01 from the Diseased Sponge of the Lubomirskia baicalensis. Curr. Issues Mol. Biol. 2021, 43, 2220–2237. [Google Scholar] [CrossRef] [PubMed]
- Dantas, C.; Tauler, R.; Ferreira, M.M.C. Exploring in vivo violacein biosynthesis by application of multivariate curve resolution on fused UV–VIS absorption, fluorescence, and liquid chromatography–mass spectrometry data. Anal. Bioanal. Chem. 2012, 405, 1293–1302. [Google Scholar] [CrossRef] [PubMed]
- Shimaraev, M.N.; Troitskaya, E.S.; Gnatovskii, R.Y. Modern Climate Changes and Deep Water Temperature of Lake Baikal. Dokl. Earth Sci. 2009, 427, 804–808. [Google Scholar] [CrossRef]
- Vacelet, J.; Vacelet, E.; Gaino, E.; Gallissian, M.-F. Bacterial attack of spongin skeleton during the 1986–1990 Mediterranean sponge disease. In Sponges in Time and Space; van Kempen, T.M.G., Braekman, J.C., van Soest, R.W.M., Eds.; A.A. Balkema: Rotterdam, The Netherlands, 1994; pp. 355–362. [Google Scholar]
- Webster, N.S. Sponge disease: A global threat? Environ. Microbiol. 2007, 9, 1363–1375. [Google Scholar] [CrossRef]
- Pita, L.; Rix, L.; Slaby, B.M.; Franke, A.; Hentschel, U. The sponge holobiont in a changing ocean: From microbes to ecosystems. Microbiome 2018, 6, 46. [Google Scholar] [CrossRef]
- Erwin, P.M.; Pita, L.; López-Legentil, S.; Turon, X. Stability of sponge-associated bacteria over large seasonal shifts in temperature and irradiance. Appl. Environ. Microbiol. 2012, 78, 7358–7368. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Liu, M.; Simister, R.; Webster, N.S.; Thomas, T. Marine microbial symbiosis heats up: The phylogenetic and functional response of a sponge holobiont to thermal stress. ISME J. 2013, 7, 991–1002. [Google Scholar] [CrossRef]
- Vizza, C.; Pechal, J.L.; Benbow, M.E.; Lang, J.M.; Chaloner, D.T.; Jones, S.E.; Lamberti, G.A. Nitrate amendment reduces biofilm biomass and shifts microbial communities in remote, oligotrophic ponds. Freshw. Sci. 2018, 37, 251–263. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Acharya, V.; Singh, D.; Kumar, S. Strategies for high-altitude adaptation revealed from high-quality draft genome of non-violacein producing Janthinobacterium lividum ERGS5:01. Stand. Genom. Sci. 2018, 13, 11. [Google Scholar] [CrossRef]
- Lu, H.; Deng, T.; Cai, Z.; Liu, F.; Yang, X.; Wang, Y.; Xu, M. Janthinobacterium violaceinigrum sp. nov., Janthinobacterium aquaticum sp. nov. and Janthinobacterium rivuli sp. nov., isolated from a subtropical stream in China. Int. J. Syst. Evol. Microbiol. 2020, 70, 2719–2725. [Google Scholar] [CrossRef]
- Aslamov, I.A.; Kozlov, V.V.; Kirillin, G.B.; Mizandrontsev, I.B.; Kucher, K.M.; Makarov, M.M.; Gornov, A.Y.; Granin, N.G. Ice–water heat exchange during ice growth in Lake Baikal. J. Great Lakes Res. 2014, 40, 599–607. [Google Scholar] [CrossRef]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Camara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stauff, D.L.; Bassler, B.L. Quorum sensing in Chromobacterium violaceum: DNA recognition and gene regulation by the CviR receptor. J. Bacteriol. 2011, 193, 3871–3878. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Janthinobacterium sp. Strain SLB01 |
|---|---|
| Gram’s reaction | − |
| Motility | + |
| Nitrate reduction | + |
| H2S production | + |
| Urease production | − |
| Phenylalanine deamination | − |
| Lipolytic activity | + |
| Hydrolysis of gelatine | + |
| Starch | + |
| Casein | + |
| Arabinose | − |
| Glucose | + |
| Lactose | + |
| Mannose | + |
| Sucrose | + |
| Citrate | + |
| Adonitol | − |
| Inositol | + |
| D-Mannitol | − |
| D-Sorbitol | − |
| L-Arginine | + |
| L-Lysine | + |
| L-Ornithine | + |
| Antibiotic Tested | Symbol | Dose (µg) | Inhibition Zone Diameter (mm) | Interpretation |
|---|---|---|---|---|
| Ampicillin | AMP | 10 | 5 | R |
| Ceftriaxone | CTR | 30 | 19 | I |
| Imipenem | IPM | 10 | 6 | R |
| Nalidixic acid | NA | 30 | 13 | R |
| Levofloxacin | LE | 5 | 15 | R |
| Lomefloxacin | LOM | 10 | 21 | I |
| Norfloxacin | NX | 10 | 5 | R |
| Ofloxacin | OF | 5 | 14 | I |
| Ciprofloxacin | CIP | 5 | 22 | I |
| Amikacin | AK | 30 | 22 | S |
| Gentalyn | GEN | 10 | 8 | R |
| Gentamicin | HLG | 120 | 16 | S |
| Kanamycin | K | 30 | 18 | S |
| Netilmycin | NET | 30 | 9 | R |
| Tobramycin | TOB | 10 | 15 | S |
| Doxycycline | DO | 30 | 30 | S |
| Tetracycline | TE | 30 | 25 | S |
| Chloramphenicol | C | 30 | 5 | R |
| Erythromycin | E | 15 | 5 | R |
| Rifampicin | RIF | 5 | 20 | S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chernogor, L.; Bakhvalova, K.; Belikova, A.; Belikov, S. Isolation and Properties of the Bacterial Strain Janthinobacterium sp. SLB01. Microorganisms 2022, 10, 1071. https://doi.org/10.3390/microorganisms10051071
Chernogor L, Bakhvalova K, Belikova A, Belikov S. Isolation and Properties of the Bacterial Strain Janthinobacterium sp. SLB01. Microorganisms. 2022; 10(5):1071. https://doi.org/10.3390/microorganisms10051071
Chicago/Turabian StyleChernogor, Lubov, Kseniya Bakhvalova, Alina Belikova, and Sergei Belikov. 2022. "Isolation and Properties of the Bacterial Strain Janthinobacterium sp. SLB01" Microorganisms 10, no. 5: 1071. https://doi.org/10.3390/microorganisms10051071
APA StyleChernogor, L., Bakhvalova, K., Belikova, A., & Belikov, S. (2022). Isolation and Properties of the Bacterial Strain Janthinobacterium sp. SLB01. Microorganisms, 10(5), 1071. https://doi.org/10.3390/microorganisms10051071