
Modulation of Signal Regulatory Protein α (SIRPα) by Plasmodium Antigenic Extract: A Preliminary In Vitro Study on Peripheral Blood Mononuclear Cells

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. P. falciparum and P. vivax Antigens
2.2. PBMC Isolation and Antigenic Stimulation
2.3. Flow Cytometry Assay
2.4. Statistical Analysis
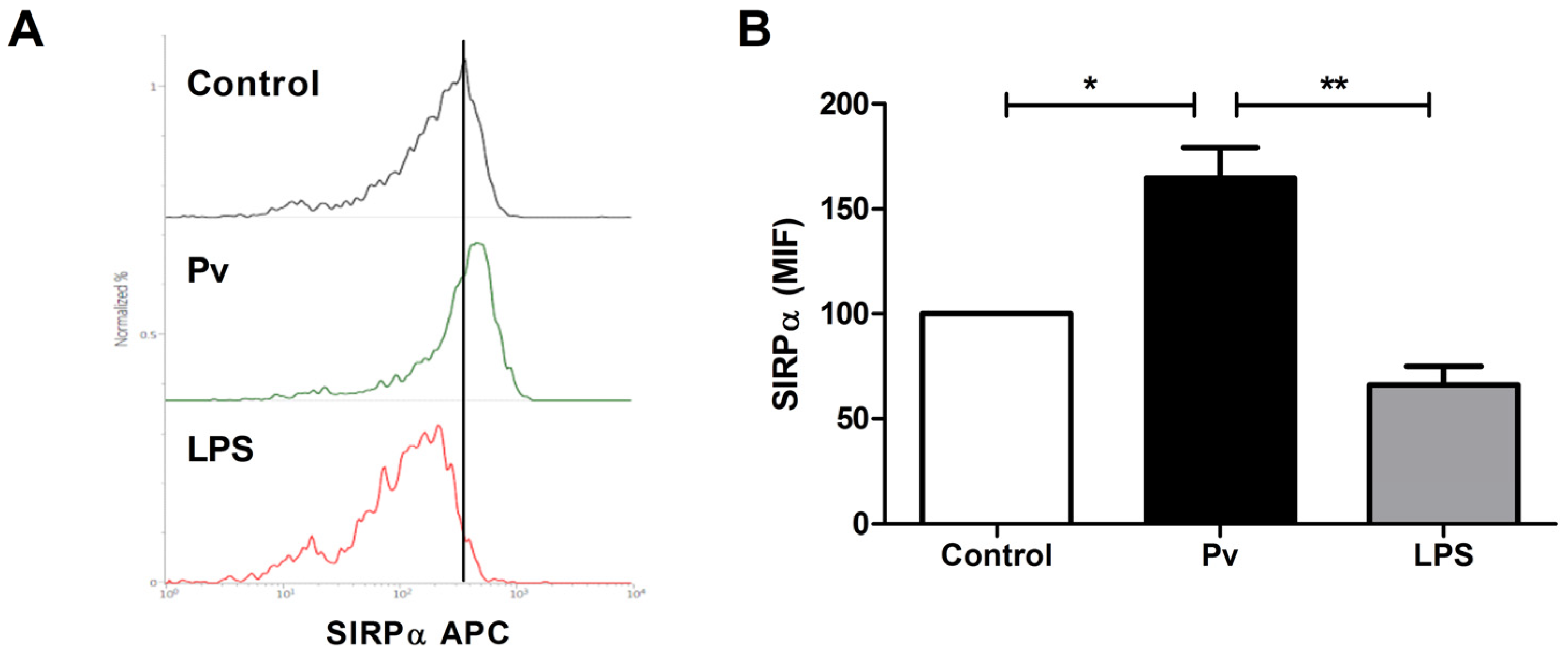
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olive, C. Pattern recognition receptors: Sentinels in innate immunity and targets of new vaccine adjuvants. Expert Rev. Vaccines 2012, 11, 237–256. [Google Scholar] [CrossRef] [PubMed]
- Beutler, B. Innate immunity: An overview. Mol. Immunol. 2004, 40, 845–859. [Google Scholar] [CrossRef]
- Lang, C.; Gross, U.; Lüder, C.G.K. Subversion of innate and adaptive immune responses by Toxoplasma gondii. Parasitol. Res. 2007, 100, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Rénia, L.; Goh, Y.S. Malaria parasites: The great escape. Front. Immunol. 2016, 7, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rikihisa, Y. Ehrlichia subversion of host innate responses. Curr. Opin. Microbiol. 2006, 9, 95–101. [Google Scholar] [CrossRef]
- Elliott, S.R.; Spurck, T.P.; Dodin, J.M.; Maier, A.G.; Voss, T.S.; Yosaatmadja, F.; Payne, P.D.; McFadden, G.I.; Cowman, A.F.; Rogerson, S.J.; et al. Inhibition of dendritic cell maturation by malaria is dose dependent and does not require Plasmodium falciparum erythrocyte membrane protein 1. Infect. Immun. 2007, 75, 3621–3632. [Google Scholar] [CrossRef] [Green Version]
- Millington, O.R.; Di Lorenzo, C.; Phillips, R.S.; Garside, P.; Brewer, J.M. Suppression of adaptive immunity to heterologous antigens during Plasmodium infection through hemozoin-induced failure of dendritic cell function. J. Biol. 2006, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Urban, B.; Ferguson, D.; Pain, A.; Willcox, N.; Plebanski, M.; Austyn, J.M.; Roberts, D.J. Plasmodium falciparum-infected erythrocytes modulate the maturation of dendritic cells. Nature 1999, 400, 73–77. [Google Scholar] [CrossRef]
- Leitner, W.W.; Krzych, U. Plasmodium falciparum malaria blood stage parasites preferentially inhibit macrophages with high phagocytic activity. Parasite Immunol. 1997, 19, 103–110. [Google Scholar] [CrossRef]
- Sampaio, N.G.; Eriksson, E.M.; Schofield, L. Plasmodium falciparum PfEMP1 modulates monocyte/macrophage transcription factor activation and cytokine and chemokine responses. Infect. Immun. 2017, 86, e00447-17. [Google Scholar] [CrossRef] [Green Version]
- Schwarzer, E.; Alessio, M.; Ulliers, D.; Arese, P. Phagocytosis of the malarial pigment, hemozoin, impairs expression of major histocompatibility complex class II antigen, CD54, and CD11c in human monocytes. Infect. Immun. 1998, 66, 1601–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarzer, E.; Arese, P. Phagocytosis of malarial pigment hemozoin inhibits NADPH-oxidase activity in human monocyte-derived macrophages. Biochim. Biophys. Acta 1996, 1316, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Urban, B.C.; Hien, T.T.; Day, N.P.; Phu, N.H.; Roberts, R.; Pongponratn, E.; Jones, M.; Mai, N.T.H.; Bethell, D.; Turner, G.D.H.; et al. Fatal Plasmodium falciparum malaria causes specific patterns of splenic architectural disorganization. Infect. Immun. 2005, 73, 1986–1994. [Google Scholar] [CrossRef] [Green Version]
- Van den Berg, T.K.; van Beek, E.M.; Bühring, H.J.; Colonna, M.; Hamaguchi, M.; Howard, C.J.; Kasuga, M.; Liu, Y.; Matozaki, T.; Neel, B.G.; et al. A nomenclature for signal regulatory protein family members. J. Immunol. 2005, 175, 7788–7789. [Google Scholar] [CrossRef] [Green Version]
- Daëron, M.; Jaeger, S.; Du Pasquier, L.; Vivier, E. Immunoreceptor tyrosine-based inhibition motifs: A quest in the past and future. Immunol. Rev. 2008, 224, 11–43. [Google Scholar] [CrossRef] [PubMed]
- Gardai, S.J.; Xiao, Y.-Q.; Dickinson, M.; Nick, J.A.; Voelker, D.R.; Greene, K.E.; Henson, P.M. By binding SIRPalpha or calreticulin/CD91, lung collectins act as dual function surveillance molecules to suppress or enhance inflammation. Cell 2003, 115, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Janssen, W.J.; McPhillips, K.A.; Dickinson, M.G.; Linderman, D.J.; Morimoto, K.; Xiao, Y.Q.; Oldham, K.M.; Vandivier, R.W.; Henson, P.M.; Gardai, S.J. Surfactant proteins A and D suppress alveolar macrophage phagocytosis via interaction with SIRP alpha. Am. J. Respir. Crit. Care Med. 2008, 178, 158–167. [Google Scholar]
- Latour, S.; Tanaka, H.; Demeure, C.; Mateo, V.; Rubio, M.; Brown, E.J.; Maliszewski, C.; Lindberg, F.P.; Oldenborg, A.; Ullrich, A.; et al. Bidirectional negative regulation of human T and dendritic cells by CD47 and its cognate receptor signal-regulator protein-alpha: Down-regulation of IL-12 responsiveness and inhibition of dendritic cell activation. J. Immunol. 2001, 167, 2547–2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.E.; Patel, V.; Seatter, S.D.; Deehan, M.R.; Brown, M.H.; Brooke, G.P.; Goodridge, H.S.; Howard, C.J.; Rigley, K.P.; Harnett, W.; et al. A novel MyD-1 (SIRP-1alpha) signaling pathway that inhibits LPS-induced TNFalpha production by monocytes. Blood 2003, 102, 2532–2540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trager, W.; Jensen, J.B. Human malaria parasites in continuous culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef]
- Moll, K.; Kaneko, A.; Scherf, A.; Wahlgren, M. Methods in Malaria Research; MR4/ATCC: Manassas, VA, USA, 2013. [Google Scholar]
- Carvalho, B.O.; Lopes, S.; Nogueira, P.A.; Orlandi, P.P.; Bargieri, D.; Blanco, Y.C.; Mamoni, R.; Leite, J.A.; Rodrigues, M.M.; Soares, I.D.S.; et al. On the cytoadhesion of Plasmodium vivax-infected erythrocytes. J. Infect. Dis. 2010, 202, 638–647. [Google Scholar]
- Goonewardene, R.; Carter, R.; Gamage, C.P.; Del Giudice, G.; David, P.H.; Howie, S.; Mendis, K.N. Human T cell proliferative responses to Plasmodium vivax antigens: Evidence of immunosuppression following prolonged exposure to endemic malaria. Eur. J. Immunol. 1990, 20, 1387–1391. [Google Scholar] [CrossRef] [PubMed]
- Riley, E.M.; Andersson, G.; Otoo, L.N.; Jepsen, S.; Greenwood, B.M. Cellular immune responses to Plasmodium falciparum antigens in Gambian children during and after an acute attack of falciparum malaria. Clin. Exp. Immunol. 1988, 73, 17–22. [Google Scholar]
- Wangoo, A.; Ganguly, N.K.; Mahajan, R.C. Immunosuppression in murine malaria: Suppressor role of macrophages and their products during acute and chronic Plasmodium berghei infection. APMIS 1990, 98, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Bobade, D.; Khandare, A.V.; Deval, M.; Shastry, P.; Deshpande, P. Hemozoin-induced activation of human monocytes toward M2-like phenotype is partially reversed by antimalarial drugs-chloroquine and artemisinin. MicrobiologyOpen 2019, 8, e00651. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, A.F.; Read, R.C.; Arevalo-Herrera, M.; Herrera, S.; Elliott, T.; Polak, M.E. Malaria systems immunology: Plasmodium vivax induces tolerance during primary infection through dysregulation of neutrophils and dendritic cells. J. Infect. 2018, 77, 440–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, H.A.D.S.D.; Costa-Correa, E.H.; Bianco-Junior, C.; Andrade, M.C.R.; Lima-Junior, J.D.C.; Pratt-Riccio, L.R.; Daniel-Ribeiro, C.T.; Totino, P.R.R. Detection of signal regulatory protein α in Saimiri sciureus (Squirrel Monkey) by anti-human monoclonal antibody. Front. Immunol. 2017, 8, 1814. [Google Scholar] [CrossRef]
- Kong, X.-N.; Yan, H.-X.; Chen, L.; Dong, L.-W.; Yang, W.; Liu, Q.; Yu, L.-X.; Huang, D.-D.; Liu, S.-Q.; Liu, H.; et al. LPS-induced down-regulation of signal regulatory protein α contributes to innate immune activation in macrophages. J. Exp. Med. 2007, 204, 2719–2731. [Google Scholar] [CrossRef]
- Liu, Q.; Wen, W.; Tang, L.; Qin, C.-J.; Lin, Y.; Zhang, H.-L.; Wu, H.; Ashton, C.; Wu, H.-P.; Ding, J.; et al. Inhibition of SIRPα in dendritic cells potentiates potent antitumor immunity. Oncoimmunology 2016, 5, e1183850. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.-W.; Kong, X.-N.; Yan, H.-X.; Yu, L.-X.; Chen, L.; Yang, W.; Liu, Q.; Huang, D.-D.; Wu, M.-C.; Wang, H.-Y. Signal regulatory protein alpha negatively regulates both TLR3 and cytoplasmic pathways in type I interferon induction. Mol. Immunol. 2008, 45, 3025–3035. [Google Scholar] [CrossRef]
- Baral, P.; Utaisincharoen, P. Involvement of signal regulatory protein α, a negative regulator of Toll-like receptor signaling, in impairing the MyD88-independent pathway and intracellular killing of Burkholderia pseudomallei-infected mouse macrophages. Infect. Immun. 2012, 80, 4223–4231. [Google Scholar] [CrossRef] [Green Version]
- Londino, J.D.; Gulick, D.; Isenberg, J.S.; Mallampalli, R.K. Cleavage of signal regulatory protein α (SIRPα) enhances inflammatory signaling. J. Biol. Chem. 2015, 290, 31113–31125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burel, J.G.; Apte, S.H.; McCarthy, J.S.; Doolan, D.L. Plasmodium vivax but not Plasmodiumfalciparum blood-stage infection in humans is associated with the expansion of a CD8+ T cell population with cytotoxic potential. PLoS Negl. Trop. Dis. 2016, 10, e0005031. [Google Scholar]
- Goncalves, R.M.; Scopel, K.K.G.; Bastos, M.S.; Ferreira, M.U. Cytokine balance in human malaria: Does Plasmodium vivax elicit more inflammatory responses than Plasmodium falciparum? PLoS ONE 2012, 7, e44394. [Google Scholar] [CrossRef] [PubMed]
- Veillette, A.; Chen, J. SIRPα-CD47 immune checkpoint blockade in anticancer therapy. Trends Immunol. 2018, 39, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Ayi, K.; Lu, Z.; Serghides, L.; Ho, J.M.; Finney, C.; Wang, J.; Liles, W.C.; Kain, K.C. CD47-SIRPα interactions regulate macrophage uptake of Plasmodium falciparum-infected erythrocytes and clearance of malaria in vivo. Infect. Immun. 2016, 84, 2002–2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roquilly, A.; Jacqueline, C.; Davieau, M.; Mollé, A.; Sadek, A.; Fourgeux, C.; Rooze, P.; Broquet, A.; Misme-Aucouturier, B.; Chaumette, T.; et al. Alveolar macrophages are epigenetically altered after inflammation, leading to long-term lung immunoparalysis. Nat. Immunol. 2020, 21, 636–648. [Google Scholar] [CrossRef]
- Haniffa, M.; Bigley, V.; Collin, M. Human mononuclear phagocyte system reunited. Semin. Cell Dev. Biol. 2015, 41, 59–69. [Google Scholar] [CrossRef]
- Galgani, M.; Busiello, I.; Censini, S.; Zappacosta, S.; Racioppi, L.; Zarrilli, R. Helicobacter pylori induces apoptosis of human monocytes but not monocyte-derived dendritic cells: Role of the cag pathogenicity island. Infect. Immun. 2004, 72, 4480–4485. [Google Scholar] [CrossRef] [Green Version]
- Khan, N.; Vidyarthi, A.; Pahari, S.; Agrewala, J.N. Distinct strategies employed by dendritic cells and macrophages in restricting Mycobacterium tuberculosis infection: Different philosophies but same desire. Int. Rev. Immunol. 2016, 35, 386–398. [Google Scholar] [CrossRef]
- Manuzak, J.; Dillon, S.; Wilson, C. Differential interleukin-10 (IL-10) and IL-23 production by human blood monocytes and dendritic cells in response to commensal enteric bacteria. Clin. Vaccine Immunol. 2012, 19, 1207–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norimatsu, M.; Harris, J.; Chance, V.; Dougan, G.; Howard, C.J.; Villarreal-Ramos, B. Differential response of bovine monocyte-derived macrophages and dendritic cells to infection with Salmonella typhimurium in a low-dose model in vitro. Immunology 2003, 108, 55–61. [Google Scholar] [CrossRef]
- Ahvazi, B.C.; Jacobs, P.; Stevenson, M.M. Role of macrophage-derived nitric oxide in suppression of lymphocyte proliferation during blood-stage malaria. J. Leukoc. Biol. 1995, 58, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Ding, Y.; Zhou, T.; Fu, X.; Xu, W. Plasmodium yoelii blood-stage primes macrophage-mediated innate immune response through modulation of toll-like receptor signaling. Malar. J. 2012, 11, 104. [Google Scholar] [CrossRef] [Green Version]
- Ocaña-Morgner, C.; A Wong, K.; Rodriguez, A. Interactions between dendritic cells and CD4+ T cells during Plasmodium infection. Malar. J. 2008, 7, 88. [Google Scholar] [CrossRef] [Green Version]
- Sponaas, A.-M.; Belyaev, N.; Falck-Hansen, M.; Potocnik, A.; Langhorne, J. Transient deficiency of dendritic cells results in lack of a merozoite surface protein 1-specific CD4 T cell response during peak Plasmodium chabaudi blood-stage infection. Infect. Immun. 2012, 80, 4248–4256. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, P.d.C.; de Souza, H.A.d.S.; Blanco, C.M.; Santos-de-Oliveira, L.; Pratt-Riccio, L.R.; Daniel-Ribeiro, C.T.; Totino, P.R.R. Modulation of Signal Regulatory Protein α (SIRPα) by Plasmodium Antigenic Extract: A Preliminary In Vitro Study on Peripheral Blood Mononuclear Cells. Microorganisms 2022, 10, 903. https://doi.org/10.3390/microorganisms10050903
Martins PdC, de Souza HAdS, Blanco CM, Santos-de-Oliveira L, Pratt-Riccio LR, Daniel-Ribeiro CT, Totino PRR. Modulation of Signal Regulatory Protein α (SIRPα) by Plasmodium Antigenic Extract: A Preliminary In Vitro Study on Peripheral Blood Mononuclear Cells. Microorganisms. 2022; 10(5):903. https://doi.org/10.3390/microorganisms10050903
Chicago/Turabian StyleMartins, Priscilla da Costa, Hugo Amorim dos Santos de Souza, Carolina Moreira Blanco, Luana Santos-de-Oliveira, Lilian Rose Pratt-Riccio, Cláudio Tadeu Daniel-Ribeiro, and Paulo Renato Rivas Totino. 2022. "Modulation of Signal Regulatory Protein α (SIRPα) by Plasmodium Antigenic Extract: A Preliminary In Vitro Study on Peripheral Blood Mononuclear Cells" Microorganisms 10, no. 5: 903. https://doi.org/10.3390/microorganisms10050903
APA StyleMartins, P. d. C., de Souza, H. A. d. S., Blanco, C. M., Santos-de-Oliveira, L., Pratt-Riccio, L. R., Daniel-Ribeiro, C. T., & Totino, P. R. R. (2022). Modulation of Signal Regulatory Protein α (SIRPα) by Plasmodium Antigenic Extract: A Preliminary In Vitro Study on Peripheral Blood Mononuclear Cells. Microorganisms, 10(5), 903. https://doi.org/10.3390/microorganisms10050903