
Facing the Human and Animal Brucellosis Conundrums: The Forgotten Lessons


Abstract
:1. Introduction
2. The Conundrums of Human Brucellosis
2.1. The Many Names of the Human Disease
2.2. The Clinical Course of Brucellosis: A Journey from Chaos to Order to Chaos
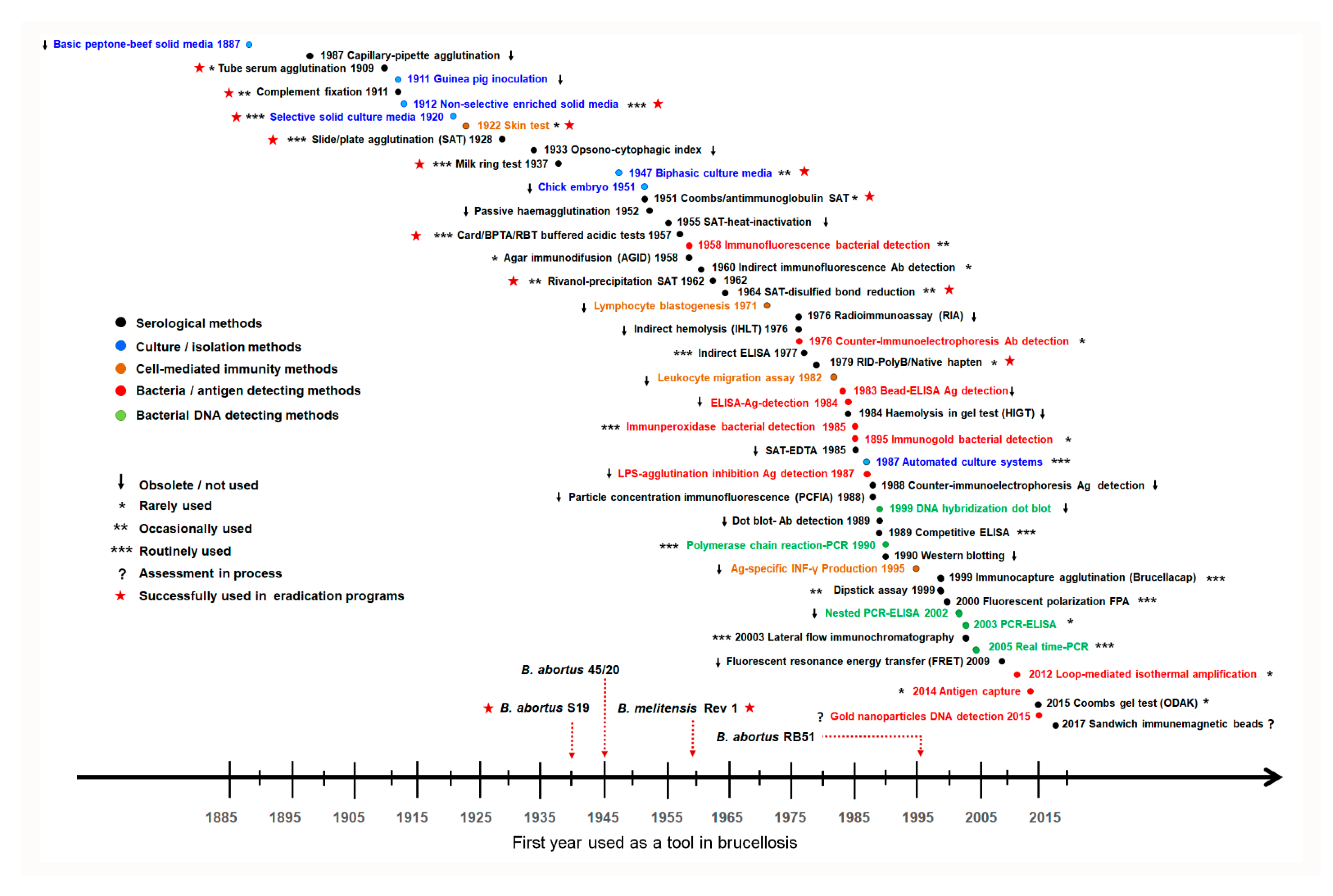
2.3. The Repertoire of Diagnostic Tools and Their Misuse
2.4. Prevalence and the Reality of Numbers
3. The Conundrums of Animal Brucellosis
3.1. True Gaps of Knowledge
3.2. Misestimating Prevalence
3.3. From Wrong Prevalence and Official Eradication Programs to Reality
3.4. Sophisticated and Expensive Is Not Better
3.5. The “Confirmatory Test” Confusion and the Pitfalls of Primary Binding Assays
3.6. Jumping to Conclusions: PCR as a Direct Diagnostic Test
3.7. Ariadne’s Thread in the Cattle Vaccine Labyrinth
4. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maurois, A. The Life of Sir Alexander Fleming; Jonatan Cape: London, UK, 1959. [Google Scholar]
- Hughes, M.L. Mediterranean, Malta, or Undulant Fever; MacMillan & Co., Ed.: London, UK; New York, NY, USA, 1897. [Google Scholar]
- Bruce, D. Note on the discovery of a microorganism in Malta Fever. Practitioner 1887, 40, 161–170. [Google Scholar]
- Meyer, K.F.; Shaw, E.B. A comparison of the morphologic, cultural and biochemical characteristics of B. abortus and B. melitensis: Studies on the genus Brucella Nov. Gen. I. J. Infect. Dis. 1920, 27, 173–184. [Google Scholar] [CrossRef]
- Spink, W.W. The evolution of the concept that brucellosis is a disease of animals and man. In The Nature of Brucellosis; The University of Minnesota Press: Minneapolis, MN, USA, 1956; pp. 3–27. [Google Scholar]
- Bosilkovski, M.; Krteva, L.; Dimzova, M.; Vidinic, I.; Sopova, Z.; Spasovska, K. Human brucellosis in Macedonia—10 years of clinical experience in endemic region. Croat. Med. J. 2010, 51, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalrymple-Champneys, W. Clinical features. In Brucella Infection and Undulant Fever in Man; Oxford University Press: London, UK, 1960; pp. 61–94. [Google Scholar]
- Parlak, M.; Akbayram, S.; Doğan, M.; Tuncer, O.; Bayram, Y.; Ceylan, N.; Özlük, S.; Akbayram, H.T.; Öner, A. Clinical manifestations and laboratory findings of 496 children with brucellosis in Van, Turkey. Pediatr. Int. 2015, 57, 586–589. [Google Scholar] [CrossRef]
- Pedro-Pons, A.; Farreras, P.; Foz, A.; Sur s, J.; Surinyach, R.; Frouchtman, R. Enfermedades infecciosas. II.A. Enfermedades producidas por bacterias. Brucelosis. In Patología y Clínica Médicas; Pedro-Pons, A., Salvat, Eds.; S.A.: Barcelona, Spain, 1968; Volume VI, pp. 338–374. [Google Scholar]
- Moreno, E.; Moriyón, I. Brucella melitensis: A nasty bug with hidden credentials for virulence. Proc. Natl. Acad. Sci. USA 2002, 99, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Roop, R.M.; Barton, I.S.; Hopersberger, D.; Martin, D.W. Uncovering the hidden credentials of Brucella virulence. Microbiol. Mol. Biol. Rev. 2021, 85, e00021-19. [Google Scholar] [CrossRef]
- Barquero-Calvo, E.; Chaves-Olarte, E.; Weiss, D.S.; Guzmán-Verri, C.; Chacón-Díaz, C.; Rucavado, A.; Moriyón, I.; Moreno, E. Brucella abortus uses a stealthy strategy to avoid activation of the innate immune system during the onset of infection. PLoS ONE 2007, 2, e631. [Google Scholar] [CrossRef]
- Zheludkov, M.M.; Tsirelson, L.E. Reservoirs of Brucella infection in nature. Biol. Bull. 2010, 37, 709–715. [Google Scholar] [CrossRef]
- Ducrotoy, M.J.; Bertu, W.J.; Matope, G.; Cadmus, S.; Conde-Álvarez, R.; Gusi, A.M.; Welburn, S.; Ocholi, R.; Blasco, J.M.; Moriyón, I. Brucellosis in Sub-Saharan Africa: Current challenges for management, diagnosis and control. Acta Trop. 2017, 165, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Mattar, S.; Tique, V.; Miranda, J.; Montes, E.; Garzon, D. Undifferentiated tropical febrile illness in Cordoba, Colombia: Not everything is dengue. J. Infect. Public Health 2017, 10, 507–512. [Google Scholar] [CrossRef]
- Lai, S.; Zhou, H.; Xiong, W.; Gilbert, M.; Huang, Z.; Yu, J.; Yin, W.; Wang, L.; Chen, Q.; Li, Y.; et al. Changing epidemiology of human brucellosis, China, 1955–2014. Emerg. Infect. Dis. 2017, 23, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Musallam, I.; Ndour, A.P.; Yempabou, D.; Ngong, C.A.C.; Dzousse, M.F.; Mouiche-Mouliom, M.M.; Feussom, J.M.K.; Ntirandekura, J.B.; Ntakirutimana, D.; Fane, A.; et al. Brucellosis in dairy herds: A public health concern in the milk supply chains of West and Central Africa. Acta Trop. 2019, 197, 105042. [Google Scholar] [CrossRef] [PubMed]
- Norman, F.F.; Monge-Maillo, B.; Chamorro-Tojeiro, S.; Pérez-Molina, J.-A.; López-Vélez, R. Imported brucellosis: A case series and literature review. Travel Med. Infect. Dis. 2016, 14, 182–199. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.L. Undulant (Malta) fever. Lancet 1896, 148, 238–239. [Google Scholar] [CrossRef] [Green Version]
- Evans, A.C. Further studies on Bacterium abortus and related bacteria: I. The pathogenicity of Bacterium lipolyticus for guinea-pigs. J. Infect. Dis. 1918, 22, 576–579. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Castañeda, M. Brucelosis, 3rd ed.; La Prensa Médica Mexicana S.A.: Mexico City, Mexico, 1986; ISBN 968-435-103-8. [Google Scholar]
- Spink, W.W. The pathogenesis of brucellosis. In The Nature of Brucellosis; The University of Minnesota Press: Minneapolis, MN, USA, 1956; pp. 113–144. [Google Scholar]
- Spink, W.W. The natural course of brucellosis. In The Nature of Brucellosis; The University of Minnesota Press: Minneapolis, MN, USA, 1956; pp. 145–170. [Google Scholar]
- Evans, A.C. Chronic brucellosis. J. Am. Med. Assoc. 1934, 103, 665–667. [Google Scholar] [CrossRef]
- Angle, F.E. Treatment of acute and chronic brucellosis (undulant fever): Personal observation of one hundred cases over a period of seven years. J. Am. Med. Assoc. 1935, 105, 939–942. [Google Scholar] [CrossRef]
- Janbon, M. Considerations on the General Development of Human Brucellosis; WHO/FAO Expert Panel Brucell. WHO/Bruc/28; WHO: Geneva, Switzerland, 1950. [Google Scholar]
- De Villafañe-Lastra, T. Clinical features of human brucellosis. In Proceedings of the Third Inter-American Congress on Brucellosis, Washington, DC, USA, 6–10 November 1950; pp. 191–209. [Google Scholar]
- Zafari, P.; Zarifian, A.; Alizadeh-Navaei, R.; Taghadosi, M.; Rafiei, A. Association between polymorphisms of cytokine genes and brucellosis: A comprehensive systematic review and meta-analysis. Cytokine 2020, 127, 154949. [Google Scholar] [CrossRef]
- Dean, A.S.; Crump, L.; Greter, H.; Hattendorf, J.; Schelling, E.; Zinsstag, J. Clinical manifestations of human brucellosis: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2012, 6, e1929. [Google Scholar] [CrossRef] [Green Version]
- Adetunji, S.A.; Ramirez, G.; Foster, M.J.; Arenas-Gamboa, A.M. A systematic review and meta-analysis of the prevalence of osteoarticular brucellosis. PLoS Negl. Trop. Dis. 2019, 13, e0007112. [Google Scholar] [CrossRef]
- Zheng, R.; Xie, S.; Niyazi, S.; Lu, X.; Sun, L.; Zhou, Y.; Zhang, Y.; Wang, K. Meta-analysis of the changes of peripheral blood T cell subsets in patients with brucellosis. J. Immunol. Res. 2018, 2018, 8439813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappas, G.; Akritidis, N.; Bosilkovski, M.; Tsianos, E. Brucellosis. N. Engl. J. Med. 2005, 353, 2325–2336. [Google Scholar] [CrossRef] [PubMed]
- Yagupsky, P.; Morata, P.; Colmenero, J.D. Laboratory diagnosis of human brucellosis. Clin. Microbiol. Rev. 2019, 33, e00073-19. [Google Scholar] [CrossRef] [PubMed]
- Guerra, M.A.; Nicoletti, P. Comparison of the susceptibility of Brucella abortus isolates obtained before and after cows were treated with oxytetracycline and streptomycin. Am. J. Vet. Res. 1986, 47, 2612–2613. [Google Scholar] [PubMed]
- Blasco, J.M.; Moreno, E.; Moriyón, I. Brucellosis vaccines and vaccine candidates. In Veterinary Vaccines. Principles and Applications, 1st ed.; Metwally, S., Viljoen, G.J., El Idrissi, A., Eds.; FAO: Rome, Italy; Wiley & Sons: Hoboken, NJ, USA, 2021; pp. 295–316. [Google Scholar]
- Blasco, J.M.; Díaz, R. Brucella melitensis Rev-1 vaccine as a cause of human brucellosis. Lancet 1993, 342, 805. [Google Scholar] [CrossRef]
- Negrón, M.E.; Kharod, G.A.; Bower, W.A.; Walke, H. Human Brucella abortus RB51 infections caused by consumption of unpasteurized domestic dairy products—United States, 2017–2019. Morb. Mortal. Wkly. Rep. Notes 2019, 68, 185. [Google Scholar] [CrossRef] [Green Version]
- Vrioni, G.; Pappas, G.; Priavali, E.; Gartzonika, C.; Levidiotou, S. An eternal microbe: Brucella DNA load persists for years after clinical cure. Clin. Infect. Dis. 2008, 46, e131-6. [Google Scholar] [CrossRef] [Green Version]
- Wright, A.E.; Smith, F. On the application of the serum test to the differential diagnosis of typhoid and Malta fever. Lancet 1897, 1, 656–659. [Google Scholar]
- Spink, W.W. The diagnosis of human brucellosis. In The Nature of Brucellosis; The University of Minnesota Press: Minneapolis, MN, USA, 1956; pp. 191–215. [Google Scholar]
- Díaz, R.; Moriyón, I. Laboratory techniques in the diagnosis of human brucellosis. In Brucellosis; Young, E.J., Corbel, M.J., Eds.; CRC Press Inc.: Boca Raton, FL, USA, 1989; pp. 73–83. [Google Scholar]
- Díaz, R.; Casanova, A.; Ariza, J.; Moriyón, I. The rose bengal test in human brucellosis: A neglected test for the diagnosis of a neglected disease. PLoS Negl. Trop. Dis. 2011, 5, e950. [Google Scholar] [CrossRef] [Green Version]
- Bodenham, R.F.; Lukambagire, A.H.S.; Ashford, R.T.; Buza, J.J.; Cash-Goldwasser, S.; Crump, J.A.; Kazwala, R.R.; Maro, V.P.; McGiven, J.; Mkenda, N.; et al. Prevalence and speciation of brucellosis in febrile patients from a pastoralist community of Tanzania. Sci. Rep. 2020, 10, 7081. [Google Scholar] [CrossRef]
- Chipwaza, B.; Mhamphi, G.G.; Ngatunga, S.D.; Selemani, M.; Amuri, M.; Mugasa, J.P.; Gwakisa, P.S. Prevalence of bacterial febrile illnesses in children in Kilosa district, Tanzania. PLoS Negl. Trop. Dis. 2015, 9, e0003750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, G.S.; Miles, A.A. The serological differentiation of smooth strains of the Brucella group. Br. J. Exp. Pathol. 1932, 13, 1–13. [Google Scholar]
- Huddleson, I.F. Laboratory diagnosis of brucellosis. In Brucellosis in Man and Animals; The Commonwealth Fund.: New York, NY, USA, 1943; pp. 214–284. [Google Scholar]
- Ross, G.R. The agglutination test in undulant fever due to Brucella abortus. A preliminary note on the value of the “abortoscope”. Trans. R. Soc. Trop. Med. Hyg. 1927, 21, 57–62. [Google Scholar] [CrossRef]
- Ducrotoy, M.J.; Conde-Álvarez, R.; Blasco, J.M.; Moriyón, I. A review of the basis of the immunological diagnosis of ruminant brucellosis. Vet. Immunol. Immunopathol. 2016, 171, 81–102. [Google Scholar] [CrossRef] [PubMed]
- Mancilla, M.; López-Goñi, I.; Moriyón, I.; Zárraga, A.M. Genomic island 2 is an unstable genetic element contributing to brucella lipopolysaccharide spontaneous smooth-to-rough dissociation. J. Bacteriol. 2010, 192, 6346–6351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancilla, M.; Marín, C.M.; Blasco, J.M.; Zárraga, A.M.; López-Goñi, I.; Moriyón, I. Spontaneous excision of the O-polysaccharide wbka glycosyltranferase gene is a cause of dissociation of smooth to rough Brucella colonies. J. Bacteriol. 2012, 194, 1860–1867. [Google Scholar] [CrossRef] [Green Version]
- de Glanville, W.A.; Conde-Álvarez, R.; Moriyón, I.; Njeru, J.; Díaz, R.; Cook, E.A.J.J.; Morin, M.; Bronsvoort, B.M.d.C.; Thomas, L.F.; Kariuki, S.; et al. Poor performance of the rapid test for human brucellosis in health facilities in Kenya. PLoS Negl. Trop. Dis. 2017, 11, e0005508. [Google Scholar] [CrossRef]
- Alumasa, L.; Thomasid, L.F.; Amanya, F.; Njorogeid, S.M.; Moriyón, I.; Makhandiaid, J.; Rushtonid, J.; Fèvre, E.M.; Falzon, L.C. Hospital-based evidence on cost-effectiveness of brucellosis diagnostic tests and treatment in Kenyan hospitals. PLoS Negl. Trop. Dis. 2021, 15, e0008977. [Google Scholar] [CrossRef]
- Alton, G.G.; Jones, L.M.; Angus, R.D.; Verger, J.M. Techniques for the Brucellosis Laboratory; INRA: Paris, France, 1988. [Google Scholar]
- Nicoletti, P. Utilization of the card test in brucellosis eradication. J. Am. Vet. Med. Assoc. 1967, 151, 1778–1783. [Google Scholar]
- Casanova, A.; Ariza, J.; Rubio, M.; Masuet, C.; Díaz, R. Brucella capt vs. classical tests in the serological diagnosis and management of human brucellosis. Clin. Vaccine Immunol. 2009, 16, 844–851. [Google Scholar] [CrossRef] [Green Version]
- Lukambagire, A.S.; Mendes, Â.J.; Bodenham, R.F.; McGiven, J.A.; Mkenda, N.A.; Mathew, C.; Rubach, M.P.; Sakasaka, P.; Shayo, D.D.; Maro, V.P.; et al. Performance characteristics and costs of serological tests for brucellosis in a pastoralist community of northern Tanzania. Sci. Rep. 2021, 11, 5480. [Google Scholar] [CrossRef] [PubMed]
- Ekiri, A.B.; Kilonzo, C.; Bird, B.H.; Vanwormer, E.; Wolking, D.J.; Smith, W.A.; Masanja, H.; Kazwala, R.R.; Mazet, J.A.K. Utility of the rose bengal test as a point-of-care test for human brucellosis in endemic African settings: A systematic review. J. Trop. Med. 2020, 2020, 6586182. [Google Scholar] [CrossRef] [PubMed]
- Oomen, L.J.; Waghela, S. The rose bengal plate test in human brucellosis. Trop. Geogr. Med. 1974, 26, 300–302. [Google Scholar] [PubMed]
- Ruiz-Mesa, J.D.; Sánchez-González, J.; Reguera, J.M.; Martín, L.; López-Palmero, S.; Colmenero, J.D. Rose Bengal test: Diagnostic yield and use for the rapid diagnosis of human brucellosis in emergency departments in endemic areas. Clin. Microbiol. Infect. 2005, 11, 221–225. [Google Scholar] [CrossRef] [Green Version]
- Steele, J.H.; Otis-Emik, L. Brucellosis incidence in the United States. In Proceedings of the Third International Brucellosis Congress, Washington DC, USA, 6–10 November 1950; pp. 28–36. [Google Scholar]
- Bruce, D. Observations on Malta fever. Br. Med. J. 1889, 1, 1101. [Google Scholar] [CrossRef] [Green Version]
- Bosilkovski, M.; Stojanov, A.; Stevanovic, M.; Karadzovski, Z.; Krstevski, K. Impact of measures to control brucellosis on disease characteristics in humans: Experience from an endemic region in the Balkans. Infect. Dis. 2017, 50, 340–345. [Google Scholar] [CrossRef]
- Dadar, M.; Shahali, Y.; Whatmore, A.M. Human brucellosis caused by raw dairy products: A review on the occurrence, major risk factors and prevention. Int. J. Food Microbiol. 2019, 292, 39–47. [Google Scholar] [CrossRef]
- Damon, S.R.; Donham, C.R.; Hutchings, L.M.; Simms, B.T.; Steele, J.H. Serology of brucellosis in rural Indiana. Public Health Rep. 1953, 68, 563. [Google Scholar] [CrossRef] [Green Version]
- Jelastopulu, E.; Bikas, C.; Petropoulos, C.; Leotsinidis, M. Incidence of human brucellosis in a rural area in Western Greece after the implementation of a vaccination programme against animal brucellosis. BMC Public Health 2008, 8, 241. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Serrano, L.P.; Ordóñez-Banegas, P.; Díaz-García, M.O.; Torres-Frías, A. Vigilancia de la brucelosis. Bol. Epidemiol. Sem. 2004, 12, 209–220. [Google Scholar]
- Spink, W.W. The dissemination of Brucella organisms from the animal reservoir. In The Nature of Brucellosis; The University of Minnesota Press: Minneapolis, MN, USA, 1956; pp. 80–89. [Google Scholar]
- Evans, A.C. Brucellosis in the United States. Am. J. Public Health Nations Health 1947, 37, 139–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosales-Galeano, C.; Puentes-Mojica, C.; Arias-Rojas, O.; Romero-Zúñiga, J.R. Epidemiological aspects of brucellosis in humans in the Health Areas of Aguas Zarcas and Los Chiles, Costa Rica, 2015–2017le. Rev. Cien. Vet. 2020, 38, 1–16. [Google Scholar] [CrossRef]
- Ávila-Granados, L.M.; García-González, D.G.; Zambrano-Varón, J.L.; Arenas-Gamboa, A.M. Brucellosis in Colombia: Current status and challenges in the control of an endemic disease. Front. Vet. Sci. 2019, 6, 321. [Google Scholar] [CrossRef] [PubMed]
- Luna-Martínez, E.; Mejía-Terán, C. Brucellosis in México: Current status and trends. Vet. Microbiol. 2002, 90, 19–30. [Google Scholar] [CrossRef]
- Godfroid, J.; Al-Dahouk, S.; Pappas, G.; Roth, F.; Matope, G.; Muma, J.; Marcotty, T.; Pfeiffer, D.; Skjerve, E. A “One Health” surveillance and control of brucellosis in developing countries: Moving away from improvisation. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappas, G.; Papadimitriou, P.; Akritidis, N.; Christou, L.; Tsianos, E.V. The new global map of human brucellosis. Lancet Infect. Dis. 2006, 6, 91–99. [Google Scholar] [CrossRef]
- Corbel, M.J. Brucellosis: An overview. Emerg. Infect. Dis. 1997, 3, 213–221. [Google Scholar] [CrossRef]
- Joint FAO/WHO Expert Committee on Brucellosis; World Health Organization. Joint FAO/WHO Expert Committee on Brucellosis: Sixth Report; WHO: Geneva, Switzerland, 1986; Volume 740. [Google Scholar]
- Pereira, C.R.; de Almeida, J.V.F.C.; de Oliveira, I.R.C.; de Oliveira, L.F.; Pereira, L.J.; Zangerônimo, M.G.; Lage, A.P.; Dorneles, E.M.S. Occupational exposure to Brucella spp.: A systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2020, 14, e0008164. [Google Scholar] [CrossRef]
- Mirnejad, R.; Jazi, F.M.; Mostafaei, S.; Sedighi, M. Epidemiology of brucellosis in Iran: A comprehensive systematic review and meta-analysis study. Microb. Pathog. 2017, 109, 239–247. [Google Scholar] [CrossRef]
- Dadar, M.; Fakhri, Y.; Shahali, Y.; Mousavi Khaneghah, A. Contamination of milk and dairy products by Brucella species: A global systematic review and meta-analysis. Food Res. Int. 2020, 128, 108775. [Google Scholar] [CrossRef]
- Dadar, M.; Shahali, Y.; Fakhri, Y.; Godfroid, J. The global epidemiology of Brucella infections in terrestrial wildlife: A meta-analysis. Transbound. Emerg. Dis. 2021, 68, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Döpfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T.; et al. World Health Organization estimates of the global and regional disease burden of 22 foodborne bacterial, protozoal, and viral diseases, 2010: A Data synthesis. PLoS Med. 2015, 12, e1001921. [Google Scholar] [CrossRef] [Green Version]
- Dean, A.S.; Crump, L.; Greter, H.; Schelling, E.; Zinsstag, J. Global burden of human brucellosis: A systematic review of disease frequency. PLoS Negl. Trop. Dis. 2012, 6, e1865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boorstin, D.J. The greatest obstacle to discovery is not ignorance—It is the illusion of knowledge. The 6 O’Clock Scholar: Librarian of Congress Daniel Boorstin And His Love Affair With Books by Carol Krucoff, Start Page K, Quote Page K, Column 2, Washington, D.C. The Washington Post, 29 January 1984. [Google Scholar]
- Corbel, M.J.; Alton, G.G.; Banai, M.; Díaz, R.; Dranovskaia, B.A.; Elberg, S.S.; Garin-Bastuji, B.; Kolar, J.; Mantovani, A.; Mousa, A.M.; et al. Brucellosis in Humans and Animals; WHO Press: Geneva, Switzerland, 2006; Volume 52, ISBN 9241547138. [Google Scholar]
- Nicoletti, P. Relationship between animal and human disease. In Brucellosis: Clinical and Laboratory Aspects; Young, E.D., Corbel, M.J., Eds.; Routledge: Boca Raton, FL, USA, 1989; pp. 41–52. ISBN 0-8493-6661-5. [Google Scholar]
- Zhang, W.-Y.; Guo, W.-D.; Sun, S.-H.; Jiang, J.-F.; Sun, H.-L.; Li, S.-L.; Liu, W.; Cao, W.-C. Human brucellosis, Inner Mongolia, China. Emerg. Infect. Dis. 2010, 16, 2001–2003. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, M.A.; Mingala, C.N.; Tubalinal, G.A.S.; Gaban, P.B.V.; Nakajima, C.; Suzuki, Y. Emerging Infectious Diseases in Water Buffalo: An Economic and Public Health Concern; IntechOpen: London, UK, 2018; Available online: https://www.intechopen.com/chapters/59095 (accessed on 1 December 2021).
- Wernery, U. Camelid brucellosis: A review. Rev. Sci. Tech. 2014, 33, 839–857. [Google Scholar] [CrossRef]
- Mick, V.; Le Carrou, G.; Corde, Y.; Game, Y.; Jay, M.; Garin-Bastuji, B. Brucella melitensis in France: Persistence in wildlife and probable spillover from alpine ibex to domestic animals. PLoS ONE 2014, 9, e94168. [Google Scholar] [CrossRef] [Green Version]
- National Academies of Sciences, Engineering, and Medicine; Division on Earth and Life Studies; Board on Agriculture and Natural Resources; Committee on Revisiting Brucellosis in the Greater Yellowstone Area. Ecology and Epidemiology of Brucella abortus in the Greater Yellowstone Ecosystem. In Revisiting Brucellosis in the Greater Yellowstone Area; National Academies Press: Washington, DC, USA, 2017. Available online: https://www.ncbi.nlm.nih.gov/books/NBK566534/#sec_ch3_3 (accessed on 1 December 2021).
- Godfroid, J.; Nielsen, K.H.; Saegerman, C. Diagnosis of brucellosis in livestock and wildlife. Croat. Med. J. 2010, 51, 296–305. [Google Scholar] [CrossRef] [Green Version]
- Simpson, G.; Thompson, P.N.; Saegerman, C.; Marcotty, T.; Letesson, J.J.; de Bolle, X.; Godfroid, J. Brucellosis in wildlife in Africa: A systematic review and meta-analysis. Sci. Rep. 2021, 5960. [Google Scholar] [CrossRef]
- Ducrotoy, M.J.; Muñoz, P.M.; Conde-Álvarez, R.; Blasco, J.M.; Moriyón, I. A systematic review of current immunological tests for the diagnosis of cattle brucellosis. Prev. Vet. Med. 2018, 151, 57–72. [Google Scholar] [CrossRef]
- Davies, G.O. Gaiger and Davies’ Veterinary Pathology and Bacteriology, 3rd ed.; Bailliere, Tindall and Cox: London, UK, 1946; p. 249. [Google Scholar]
- Alonso, S.; Dohoo, I.; Lindahl, J.; Verdugo, C.; Akuku, I.; Grace, D. Prevalence of tuberculosis, brucellosis and trypanosomiasis in cattle in Tanzania: A systematic review and meta-analysis. Anim. Health Res. Rev. 2016, 17, 16–27. [Google Scholar] [CrossRef]
- Lopes, L.B.; Nicolino, R.; Haddad, J.P. Brucellosis-risk factors and prevalence: A review. Open Vet. Sci. J. 2010, 4, 72–84. [Google Scholar] [CrossRef]
- Hegazy, Y.M.; Schley, D.; Ridler, A.; Beauvais, W.; Musallam, I.; Guitian, J. Control of Brucella melitensis in endemic settings: A simulation study in the Nile Delta, Egypt. Transbound. Emerg. Dis. 2021, 68, 2364–2375. [Google Scholar] [CrossRef] [PubMed]
- Taleski, V.; Zerva, L.; Kantardjiev, T.; Cvetnic, Z.; Erski-Biljic, M.; Nikolovski, B.; Bosnjakovski, J.; Katalinic-Jankovic, V.; Panteliadou, A.; Stojkoski, S.; et al. An overview of the epidemiology and epizootiology of brucellosis in selected countries of Central and Southeast Europe. Vet. Microbiol. 2002, 90, 147–155. [Google Scholar] [CrossRef]
- Zhang, N.; Huang, D.; Wu, W.; Liu, J.; Liang, F.; Zhou, B.; Guan, P. Animal brucellosis control or eradication programs worldwide: A systematic review of experiences and lessons learned. Prev. Vet. Med. 2018, 160, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Benkirane, A. Ovine and caprine brucellosis: World distribution and control/eradication strategies in West Asia/North Africa region. Small Rumin. Res. 2006, 62, 19–25. [Google Scholar] [CrossRef]
- Francisco, J.; Vargas, O. Brucellosis in Venezuela. Vet. Microbiol. 2002, 90, 39–44. [Google Scholar]
- Hernández-Mora, G.; Ruiz-Villalobos, N.; Bonilla-Montoya, R.; Romero-Zuniga, J.-J.; Jimenez-Arias, J.; González-Barrientos, R.; Barquero-Calvo, E.; Chacón-Díaz, C.; Rojas, N.; Chaves-Olarte, E.; et al. Epidemiology of bovine brucellosis in Costa Rica: Lessons learned from failures in the control of the disease. PLoS ONE 2017, 12, e0182380. [Google Scholar] [CrossRef] [Green Version]
- Oseguera Montiel, D.; Frankena, K.; Udo, H.; van der Zijpp, A. Opportunities for brucellosis control in Mexico: Views based on the sustainable livelihoods perspective. Front. Vet. Sci. 2019, 6, 216. [Google Scholar] [CrossRef]
- Padilla Poester, F.; Picao Goncalves, V.S.; Pereira Lage, A. Brucellosis in Brazil. Vet. Microbiol. 2002, 90, 55–62. [Google Scholar] [CrossRef]
- Alton, G.G. The national brucellosis programme of Australia. In Bovine Brucellosis. An International Symposium; Crawford, R.P., Hidalgo, R.J., Eds.; Texas A&M University Press: College Station, TX, USA, 1977; pp. 335–345. ISBN 0-89096-032-1. [Google Scholar]
- Ragan, V.E. The Animal and Plant Health Inspection Service (APHIS) brucellosis eradication program in the United States. Vet. Microbiol. 2002, 90, 11–18. [Google Scholar] [CrossRef]
- Blasco, J.M.; Molina-Flores, B. Control and eradication of Brucella melitensis infection in sheep and goats. Vet. Clin. N. Am. Food Anim. Pract. 2011, 27, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Crawford, R.P.; Hidalgo, R.J. Editors’ summary of the Symposium. In Bovine Brucellosis. An International Symposium; Crawford, R.P., Hidalgo, R.J., Eds.; Texas A&M University Press: College Station, TX, USA, 1977; pp. XIII–XXIV. ISBN 0-89096-032-1. [Google Scholar]
- Hellstrom, J.S. New Zealand is free from bovine brucellosis. Surveillance 1991, 18, 14. [Google Scholar]
- Kellar, J.A.; Dore, A. Surveillance for Bovine Brucellosis (B. abortus) in Canada… A New Direction; Animal Disease Surveillance Unpublished Report; Canadian Food Inspection Agency: Ottawa, ON, Canada, 1998. [Google Scholar]
- Scharp, D.W.; Al Khalaf, S.A.; Al Muhanna, M.W.; Cheema, R.A.; Godana, W. Use of mass vaccination with a reduced dose of Rev 1 vaccine for Brucella melitensis control in a population of small ruminants. Trop. Anim. Health Prod. 1999, 31, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Berman, D.T. Brucellosis. In Diseases of Cattle in the Tropics—Current Topics in Veterinary Medicine and Animal Science, 1st ed.; Ristic, M., McIntyre, I., Eds.; Martinus Nijhoff Publishers: The Hague, The Netherlands, 1981; Volume 6, pp. 271–281. [Google Scholar]
- Anonymous. Scientific opinion on “Performance of brucellosis diagnostic methods for bovines, sheep, and goats”. EFSA J. 2006, 432, 1–44. [Google Scholar]
- Greiner, M.; Verloo, D.; De Massis, F. Meta-analytical equivalence studies on diagnostic tests for bovine brucellosis allowing assessment of a test against a group of comparative tests. Prev. Vet. Med. 2009, 92, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Gusi, A.M.; Bertu, W.J.; Jesús de Miguel, M.; Dieste-Pérez, L.; Smits, H.L.; Ocholi, R.A.; Blasco, J.M.; Moriyón, I.; Muñoz, P.M. Comparative performance of lateral flow immunochromatography, iELISA and Rose Bengal tests for the diagnosis of cattle, sheep, goat and swine brucellosis. PLoS Negl. Trop. Dis. 2019, 13, e0007509. [Google Scholar] [CrossRef] [PubMed]
- Falzon, L.C.; Traoré, S.; Kallo, V.; Assamoi, J.-B.; Bonfoh, B.; Schelling, E. Evaluation of the fluorescence polarization assay as a rapid on-spot test for ruminant brucellosis in Côte d’Ivoire. Front. Vet. Sci. 2019, 6, 287. [Google Scholar] [CrossRef] [PubMed]
- Alton, G.G. The control of bovine brucellosis. Recent developments. Rev. Mond. Zootechol. 1981, 39, 17–24. [Google Scholar]
- McGiven, J.A. New developments in the immunodiagnosis of brucellosis in livestock and wildlife. Rev. Sci. Tech. 2013, 32, 163–176. [Google Scholar] [CrossRef]
- Gall, D.; Colling, A.; Marino, O.; Moreno, E.; Nielsen, K.H.; Pérez, B.; Samartino, L.E. Enzyme immunoassays for serological diagnosis of bovine brucellosis: A trial in Latin America. Clin. Diagn. Lab. Immunol. 1998, 5, 654–661. [Google Scholar] [CrossRef] [Green Version]
- Junqueira Junior, D.G.J.; Rosinha, G.M.S.; Carvalho, C.E.G.; Oliveira, C.E.; Sanches, C.C. Detection of Brucella spp. DNA in the semen of seronegative bulls by polymerase chain reaction. Transbound. Emerg. Dis. 2013, 60, 376–377. [Google Scholar] [CrossRef] [PubMed]
- Lourencetti, M.P.S.; Souza, M.A.; Ganda, M.R.; Santos, J.P.; Ferreira Júnior, A.; Miyashiro, S.; Lima, A.M.C. High level of B19 strain detection in Brazilian cattle semen. Trop. Anim. Health Prod. 2018, 50, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, C.; Soares, C.O.; Dos Santos, L.R.; Rosinha, G.M.S. Bovine brucellosis vaccine strain s19 detected in calves before vaccination. Genet. Mol. Res. 2020, 19, gmr18659. [Google Scholar] [CrossRef]
- Rahman, S.U.; Zhu, L.; Cao, L.; Zhang, Y.; Chu, X.; Feng, S.; Li, Y.; Wu, J.; Wang, X. Prevalence of caprine brucellosis in Anhui province, China. Vet. World 2019, 12, 558–564. [Google Scholar] [CrossRef]
- Ilhan, Z.; Solmaz, H.; Aksakal, A.; Gulhan, T.; Ekin, I.H.; Boynukara, B. Comparison of PCR assay and bacteriological culture method for the detection of Brucella melitensis in stomach content samples of aborted sheep fetuses. Dtsch. Tierarztl. Wochenschr. 2007, 114, 460–464. [Google Scholar]
- Manivannan, K.; Mahmoud, S.M.; Ramasamy, M.; Shehata, A.A.E.; Ahmed, H.; Solaimuthu, C.; Dhandapani, K. Molecular detection of brucellosis in dromedary camels of Qatar by real-time PCR technique. Comp. Immunol. Microbiol. Infect. Dis. 2021, 78, 101690. [Google Scholar] [CrossRef]
- Khamesipour, F.; Doosti, A.; Rahimi, E. Molecular study of brucellosis in camels by the use of TaqMan® real-time PCR. Acta Microbiol. Immunol. Hung. 2015, 62, 409–421. [Google Scholar] [CrossRef] [Green Version]
- Nyarku, R.; Hassim, A.; Jonker, A.; Quan, M. Development of a genus-specific Brucella Real-Time PCR assay targeting the 16S-23S rDNA internal transcribed spacer from different specimen types. Vet. Sci. 2020, 7, 175. [Google Scholar] [CrossRef]
- Philippon, A.; Renoux, G.; Plommet, M. Brucellose bovine expérimentale. II. Répartition de Brucella abortus dans l’organisme six semaines après le part et trois mois a cinq mois et demi après l’épreuve infectante. Ann. Rech. Vét. 1970, 1, 203–213. [Google Scholar]
- Plommet, M.; Fensterbank, R. Vaccination against bovine brucellosis with a low dose of strain 19 administered by the conjunctival route. III. Serological response and immunity in the pregnant cow. Ann. Rech. Vét. 1976, 7, 9–23. [Google Scholar]
- Plommet, M. Studies on experimental brucellosis in cows in France. In Bovine Brucellosis. An International SYMPOSIUM; Crawford, R.P., Hidalgo, R.J., Eds.; Texas A&M University Press: College Station, TX, USA, 1977; pp. 116–134. [Google Scholar]
- Grilló, M.J.; Barberán, M.; Blasco, J.M. Transmission of Brucella melitensis from sheep to lambs. Vet. Rec. 1997, 140, 602–605. [Google Scholar] [CrossRef]
- El-Diasty, M.; Wareth, G.; Melzer, F.; Mustafa, S.; Sprague, L.D.; Neubauer, H. Isolation of Brucella abortus and Brucella melitensis from seronegative cows is a serious impediment in brucellosis control. Vet. Sci. 2018, 5, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corner, L.A.; Alton, G.G. Persistence of Brucella abortus strain 19 infection in adult cattle vaccinated with reduced doses. Res. Vet. Sci. 1981, 31, 342–344. [Google Scholar] [CrossRef]
- Fluegel-Dougherty, A.M.F.; Cornish, T.E.; O’Toole, D.; Boerger-Fields, A.M.; Henderson, O.L.; Mills, K.W. Abortion and premature birth in cattle following vaccination with Brucella abortus strain RB51. J. Vet. Diagn. Investig. 2013, 25, 630–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plommet, M.; Plommet, A.M. Vaccination against bovine brucellosis with a low dose of strain 19 administered by the conjunctival route II. Determination of the minimum dose leading to colonization of the regional lymph nodes of cattle. Ann. Rech. Vét. 1976, 7, 1–8. [Google Scholar] [PubMed]
- Zundel, E.; Verger, J.M.; Grayon, M.; Michel, R. Conjunctival vaccination of pregnant ewes and goats with Brucella melitensis Rev 1 vaccine: Safety and serological responses. Ann. Rech. Vét. 1992, 23, 177–188. [Google Scholar]
- OIE. Brucellosis (Infection with B. abortus, B. melitensis and B. suis). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; OIE: Paris, France, 2021; Volume 1, pp. 1–44. ISBN 978-92-9044-718-4. [Google Scholar]
- Moriyón, I.; Grilló, M.J.; Monreal, D.; González, D.; Marín, C.M.; López-Goñi, I.; Mainar-Jaime, R.C.; Moreno, E.; Blasco, J.M. Rough vaccines in animal brucellosis: Structural and genetic basis and present status. Vet. Res. 2004, 35, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Tabynov, K. Influenza viral vector-based Brucella abortus vaccine: A novel vaccine candidate for veterinary practice. Expert Rev. Vaccines 2016, 15, 1237–1239. [Google Scholar] [CrossRef] [Green Version]
- Tabynov, K.; Kydyrbayev, Z.; Ryskeldinova, S.; Yespembetov, B.; Zinina, N.; Assanzhanova, N.; Kozhamkulov, Y.; Inkarbekov, D.; Gotskina, T.; Sansyzbay, A. Novel influenza virus vectors expressing Brucella L7/L12 or Omp16 proteins in cattle induced a strong T-cell immune response, as well as high protectiveness against B. abortus infection. Vaccine 2014, 32, 2034–2041. [Google Scholar] [CrossRef] [Green Version]
- Tabynov, K.; Orynbayev, M.; Renukaradhya, G.J.; Sansyzbay, A. Simultaneous subcutaneous and conjunctival administration of the influenza viral vector-based Brucella abortus vaccine to pregnant heifers provides better protection against B. abortus 544 infection than the commercial B. abortus S19 vaccine. Vaccine 2016, 34, 5049–5052. [Google Scholar] [CrossRef]
- Chaudhuri, P.; Saminathan, M.; Ali, S.A.; Kaur, G.; Singh, S.V.; Lalsiamthara, J.; Goswami, T.K.; Singh, A.K.; Singh, S.K.; Malik, P.; et al. Immunization with Brucella abortus S19Δper conferred protection in water buffaloes against virulent challenge with B. abortus strain S544. Vaccines 2021, 9, 1423. [Google Scholar] [CrossRef] [PubMed]
- Castaño-Zubieta, M.R.; Rossetti, C.A.; García-González, D.G.; Maurizio, E.; Hensel, M.E.; Rice-Ficht, A.C.; Ficht, T.A.; Arenas-Gamboa, Á.M. Evaluation of the safety profile of the vaccine candidate Brucella melitensis 16MΔvjbR strain in goats. Vaccine 2021, 39, 617–625. [Google Scholar] [CrossRef]
- Hensel, M.E.; García-González, D.G.; Chaki, S.P.; Hartwig, A.; Gordy, P.W.; Bowen, R.; Ficht, T.A.; Arenas-Gamboa, A.M. Vaccine candidate Brucella melitensis 16MΔvjbR Is safe in a pregnant sheep model and confers protection. mSphere 2020, 5, e00120-20. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, T.F.; Haddad, J.P.A.; Paixão, T.A.; Santos, R.L. Meta-analysis of brucellosis vaccinology in natural hosts. Pesqui. Vet. Bras. 2020, 40, 604–613. [Google Scholar] [CrossRef]
- Fensterbank, R. Brucellose des bovins et des petits ruminants: Diagnostic, prophylaxie et vaccination. Rev. Sci. Tech. 1986, 5, 587–603. [Google Scholar] [CrossRef]
- De Oliveira, M.M.; Pereira, C.R.; de Oliveira, I.R.C.; Godfroid, J.; Lage, A.P.; Dorneles, E.M.S. Efficacy of Brucella abortus S19 and RB51 vaccine strains: A systematic review and meta-analysis. Transbound. Emerg. Dis. 2021, 1–20. [Google Scholar] [CrossRef]
- Denisov, A.A.; Sclyarov, O.D.; Salmakov, K.M.; Shumilov, K.V. The Russian experience in brucellosis veterinary public health. Rev. Sci. Tech. Off. Int. Epiz. 2013, 32, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, A.V.; Salmakov, K.M.; Olsen, S.C.; Plumb, G.E. A live vaccine from Brucella abortus strain 82 for control of cattle brucellosis in the Russian Federation. Anim. Health Res. Rev. 2011, 12, 113–121. [Google Scholar] [CrossRef]
- Shumilov, K.V.; Sklyarov, O.; Klimanov, A. Designing vaccines against cattle brucellosis. Vaccine 2010, 28, F31–F34. [Google Scholar] [CrossRef]
- Sklyarov, O.; Shumilov, K.; Klimanov, A.; Denisov, A. Targeted prevention of brucellosis in cattle, sheep, and goats in the Russian Federation. Vaccine 2010, 28, F54–F58. [Google Scholar] [CrossRef]
- Deqiu, S.; Donglou, X.; Jiming, Y. Epidemiology and control of brucellosis in China. Vet. Microbiol. 2002, 90, 165–182. [Google Scholar] [CrossRef]
- Sannikova, I.V.; Makhinya, O.V.; Maleev, V.V.; Deineka, D.A.; Golub, O.G.; Kovalchuk, I.V.; Lyamkin, G.I. Brutsellez v Stavropol’skom kraye: Rezul’taty 15-letnego nablyudeniya epidemiologicheskikh i klinicheskikh osobennostey. Ter. Arkh. 2015, 87, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Xin, X. Orally administrable brucellosis vaccine: Brucella suis strain 2 vaccine. Vaccine 1986, 4, 212–216. [Google Scholar] [CrossRef]
- Blasco, J.M.; Marín, C.; Jiménez de Bagüés, M.P.; Barberán, M. Efficacy of Brucella suis strain 2 vaccine against Brucella ovis in rams. Vaccine 1993, 11, 1291–1294. [Google Scholar] [CrossRef]
- Verger, J.-M.M.; Grayon, M.; Zundel, E.; Lechopier, P.; Olivier-Bernardin, V. Comparison of the efficacy of Brucella suis strain 2 and Brucella melitensis Rev. 1 live vaccines against a Brucella melitensis experimental infection in pregnant ewes. Vaccine 1995, 13, 191–196. [Google Scholar] [CrossRef]
- WHO. Report of WHO Working Group Meeting on Brucellosis Control and Eradication; WHO/CDS/VPH/92.109; Geneva, 2–4 June 1992; WHO: Geneva, Switzerland, 1992. [Google Scholar]
- Nicoletti, P. Vaccination. In Animal Brucellosis; Nielsen, K.H., Duncan, J.R., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 283–299. [Google Scholar]
- Moreno, E. Brucellosis in Central America. Vet. Microbiol. 2002, 90, 31–38. [Google Scholar] [CrossRef]
- Charbonnier, P. Brucelosis Bovina: Evaluación de los Sistemas de Vigilancia Epidemiológica Aplicados en Uruguay. Master’s Thesis, Universidad de la República de Uruguay, Montevideo, Uruguay, 2019. [Google Scholar]
- Olivares Castillo, R.; Vidal, P.; Sotomayor, C.; Norambuena, M.; Luppi Norambuena, M.; Silva Ojeda, F.; Cifuentes Díaz, M. Brucelosis en Chile: Descripción de una serie de 13 casos. Rev. Chil. Infectol. 2017, 34, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Moreno, E. Retrospective and prospective perspectives on zoonotic brucellosis. Front. Microbiol. 2014, 5, 213. [Google Scholar] [CrossRef] [Green Version]
- Berman, D.T.; Beach, B.A.; Irwin, M.R. Studies on repeated vaccination of cattle with Brucella abortus strain 19. III. The response of vaccinated and revaccinated cattle to conjunctival exposure with a virulent strain of Brucella abortus during the third gestation period. Am. J. Vet. Res. 1952, 13, 351–358. [Google Scholar]
- Cantú, A.; Aparicio, E.D.; Andrade, L.H.; Adams, G.L.; Güemes, F.S. Epidemiological study in a bovine herd with intermediate prevalence of brucellosis, vaccinated with RB51 and rfbK rough mutant strains of Brucella abortus. Vet. Méx. 2007, 38, 197–206. [Google Scholar]
- Leal Hernández, M.; Díaz-Aparicio, E.; Perez, R.; Hernández-Andrade, L.; Arellano-Reynoso, B.; Alfonseca, E.; Suárez-Guemes, F. Protection of Brucella abortus RB51 revaccinated cows, introduced in a herd with active brucellosis, with presence of atypical humoral response. Comp. Immunol. Microbiol. Infect. Dis. 2005, 28, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Gil, A.D.; Piaggio, J.M. Brucelosis bovina: Evaluación de las Pruebas Diagnósticas para Muestras Compuestas de Leche y Modelos Epidemiológicos de Difusión de la Enfermedad; Proyecto FPTA-245; FPTA; Unidad de Comunicación y Transferencia de Tecnología del INIA: Montevideo, Uruguay, 2013; Volume 36, Available online: http://www.inia.uy/Publicaciones (accessed on 1 December 2021).
- Edmonds, M.D.; Schurig, G.; Samartino, L.E.; Hoyt, P.G.; Walker, J.V.; Hagius, S.; Elzer, P.H. Biosafety of Brucella abortus strain RB51 for vaccination of mature bulls and pregnant heifers. Am. J. Vet. Res. 1999, 60, 722–725. [Google Scholar] [PubMed]
- Lopetegui, P. Erradicación de brucelosis bovina en Chile. Experiencias en el uso de la vacuna cepa RB51. In III Foro Nacional Brucelosis; SAGAR: Acapulco, Mexico, 1998; pp. 20–21. [Google Scholar]
- Palmer, M.V.; Olsen, S.C.; Cheville, N.F. Safety and immunogenicity of Brucella abortus strain RB51 vaccine in pregnant cattle. Am. J. Vet. Res. 1997, 58, 472–477. [Google Scholar]
- Uzal, F.A.; Samartino, L.E.; Schurig, G.; Carrasco, A.; Nielsen, K.H.; Cabrera, R.F.; Taddeo, H.R. Effect of vaccination with Brucella abortus strain RB51 on heifers and pregnant cattle. Vet. Res. Commun. 2000, 24, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Samartino, L.E.; Fort, M.; Gregoret, R.; Schurig, G. Use of Brucella abortus vaccine strain RB51 in pregnant cows after calfhood vaccination with strain 19 in Argentina. Prev. Vet. Med. 2000, 45, 193–199. [Google Scholar] [CrossRef]
- Cossaboom, C.M.; Kharod, G.A.; Salzer, J.S.; Tiller, R.V.; Campbell, L.P.; Wu, K.; Negron, M.E.; Ayala, N.; Evert, N.; Radowicz, J.; et al. Notes from the field: Brucella abortus vaccine strain RB51 infection and exposures associated with raw milk consumption—Wise County, Texas, 2017. MMWR. Morb. Mortal. Wkly. Rep. 2018, 67, 286. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.; Her, M.; Kim, J.; Hwang, I.; Ku, B.; Park, Y.; Yoo, H. Bovine brucellosis in Korea. J. Zoonoses 2010, 1, 1–15. [Google Scholar]
- Mainar-Jaime, R.C.; Marín, C.M.; de Miguel, M.J.; Muñoz, P.M.; Blasco, J.M. Experiences on the use of RB51 vaccine in Spain. In Proceedings of the Brucellosis 2008 International Conference, London, UK, 10–13 September 2008; pp. 10–13. [Google Scholar]
- Palmer, M.V.; Cheville, N.F.; Jensen, A.E. Experimental infection of pregnant cattle with the vaccine candidate Brucella abortus strain RB51: Pathologic, bacteriologic, and serologic findings. Vet. Pathol. 1996, 33, 682–691. [Google Scholar] [CrossRef] [Green Version]
- Sanz, C.; Sáez, J.L.; Álvarez, J.; Cortes, M.; Pereira, G.; Reyes, A.; Rubio, F.; Martín, J.; García, N.; Domínguez, L.; et al. Mass vaccination as a complementary tool in the control of a severe outbreak of bovine brucellosis due to Brucella abortus in Extremadura, Spain. Prev. Vet. Med. 2010, 97, 119–125. [Google Scholar] [CrossRef]
- Van Metre, D.C.; Kennedy, G.A.; Olsen, S.C.; Hansen, G.R.; Ewalt, D.R. Brucellosis induced by RB51 vaccine in a pregnant heifer. J. Am. Vet. Med. Assoc. 1999, 215, 1491–1493. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Adeno-tifo fever | Intermittent typhoid fever |
| Atypical infectious fever | Levant fever, |
| Atypical typhoid fever | Malta fever |
| Barcelona Fever | Mediterranean fever |
| Bruce septicemia | Mediterranean gastric remittent fever |
| Capricious fever | Ilo-tifo to sudoral form |
| Cartagena fever | Mediterranean tuberculosis |
| Cesspool fever, | Melitensis septicemia |
| Climatic fever | Melitococcia |
| Continuous epidemic fever | Melitosis |
| Corps disease | Mephitic fever, |
| Country fever, | Miliary fever |
| Crazy fever | Napolitan Fever |
| Cretan fever, | One 100 clinical form disease |
| Crimean fever | Phthisis |
| Cyprus fever, | Pseudo-tifo |
| Dust fever | Pythogenic septicemia |
| Faeco-malarial fever | Recurrent fever |
| Febricola typhosa | Remittent fever |
| Febris complicata | Rock fever |
| Febris melitensis | Sewage fever, |
| Febris sudoralis | Simple continued fever |
| Gastro-bilious fever | Town fever, |
| Gibraltar fever | Typho-malarial fever |
| Goat fever | Undulant fever |
| Requisite | Comments | |
|---|---|---|
| Animals | Brucellosis-free status | Any contact with the pathogen would biased results in impossible to predict and individually variable ways. In areas not free of brucellae, selection based on serology is not acceptable because of the test(s) DSe/DSp and latent carrier issues, particularly in young animals. |
| Other diseases | Healthy and rigorously proved to be free from confounding pathologies and infections, particularly those causing reproductive failure. | |
| Homogeneity | Breed, age, sex, and similar physiological status; pregnancy synchronized and monitored throughout the process. | |
| Challenge | Route | Through the conjunctiva because it reproduces a common natural infection route (subcutaneous, intravenous, or intramuscular are highly artificial). |
| Strain | In a previous test in the laboratory performing the experiments, the strain has to reproduce the multiplication in mouse spleens characteristic of virulent strains. Extreme precautions should be taken to avoid any degree of attenuation, including master seed/inoculum strategy and S to R dissociation controls for every inoculum. | |
| Vaccination-challenge interval | From 6 months to 1 year, depending on the age at vaccination (shorter periods do not provide information on sustained immunity). | |
| Pregnancy | Pregnant animals should be challenged when most susceptible to abortion (mid-pregnancy). Later times progressively reduce the development of lesions of a high enough intensity to damage fetus development. | |
| Assessments | Selective media | Strictly necessary. Commensal bacteria easily overgrow brucellae in milk, vaginal fluids, or semen. It is technically unfeasible to obtain perfectly clean necropsy samples of organs, even after surface disinfection, and microorganisms other than brucellae can be present within lymph nodes. The selective medium (better a combination of media) should be chosen, noting that some are inhibitory for some species or strains (Farrell’s for B. melitensis, B. ovis, and B. abortus RB51). |
| Detection threshold | As brucellae can be present in insufficient numbers and excreted and/or increase after sexual maturity or pregnancy, maximizing detection is critical. Thick homogenates in a minimal volume of diluent should be directly seeded (up to 0,5 mL of tissue/plate) on several plates (sensitivity up to 1 CFU/g of tissue), not only dilutions. | |
| Abortions | Protection against abortion with no complete protection is not valid as it can be counterproductive (non-aborted infected animals shed brucellae after delivery and give birth to infected offspring serologically negative latent carriers until first pregnancy when they abort and spread the disease). | |
| Controls | Non vaccinated | Mandatory. The infection rate should be close to 100%, and the infecting strain is widespread among organs/lymph nodes. |
| Reference vaccine | S19 and Rev 1 controls (OIE reference vaccines; CFU number and absence of dissociation assessed) are strictly necessary. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, E.; Blasco, J.-M.; Moriyón, I. Facing the Human and Animal Brucellosis Conundrums: The Forgotten Lessons. Microorganisms 2022, 10, 942. https://doi.org/10.3390/microorganisms10050942
Moreno E, Blasco J-M, Moriyón I. Facing the Human and Animal Brucellosis Conundrums: The Forgotten Lessons. Microorganisms. 2022; 10(5):942. https://doi.org/10.3390/microorganisms10050942
Chicago/Turabian StyleMoreno, Edgardo, José-María Blasco, and Ignacio Moriyón. 2022. "Facing the Human and Animal Brucellosis Conundrums: The Forgotten Lessons" Microorganisms 10, no. 5: 942. https://doi.org/10.3390/microorganisms10050942
APA StyleMoreno, E., Blasco, J.-M., & Moriyón, I. (2022). Facing the Human and Animal Brucellosis Conundrums: The Forgotten Lessons. Microorganisms, 10(5), 942. https://doi.org/10.3390/microorganisms10050942