
Physiologic, Genomic, and Electrochemical Characterization of Two Heterotrophic Marine Sediment Microbes from the Idiomarina Genus

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media, Cultivation, and Physiologic Tests
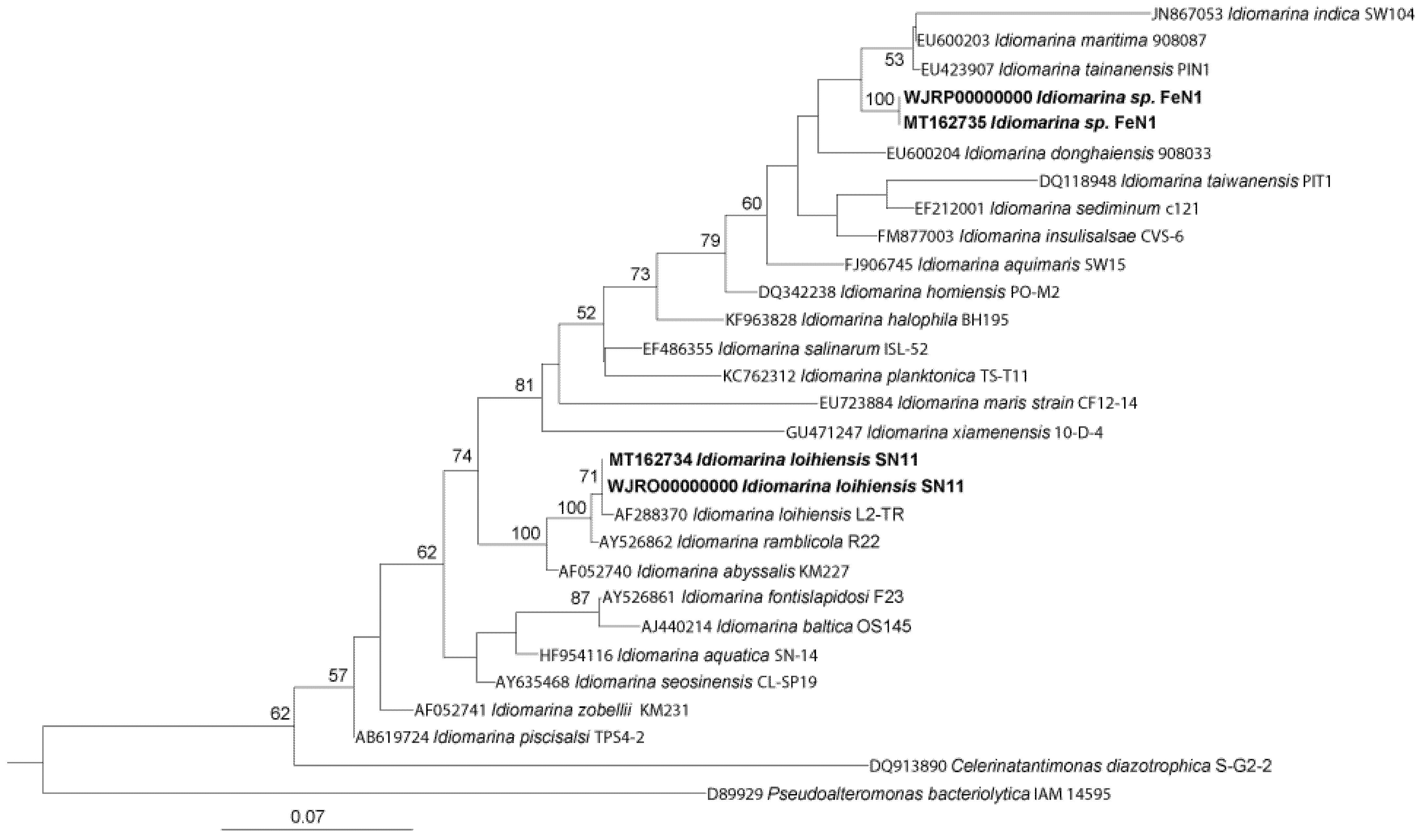
2.2. Phylogenetic Analysis
2.3. Genomic Sequencing and Analysis
2.4. Lipid Analysis
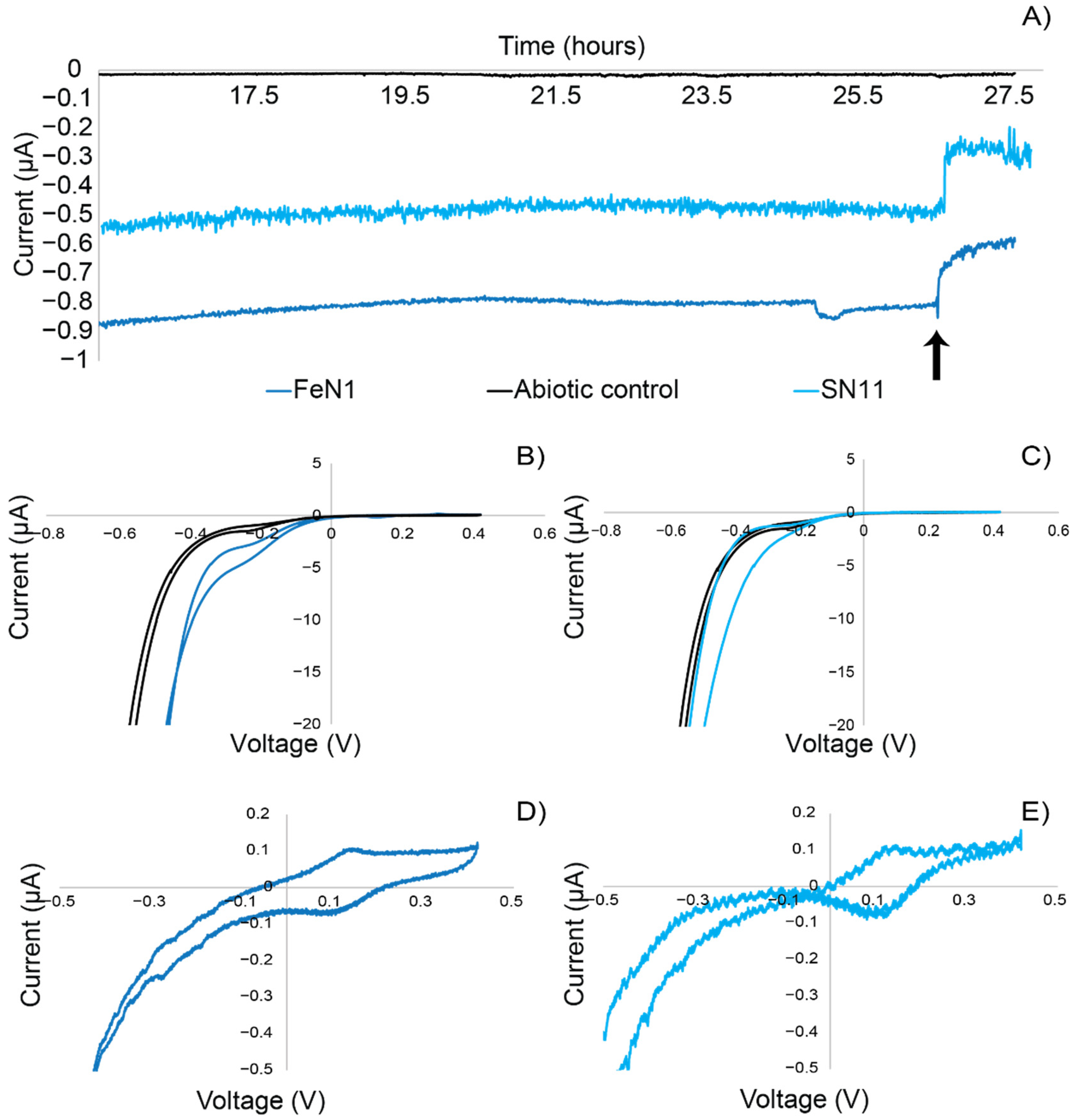
2.5. Electrochemical Techniques
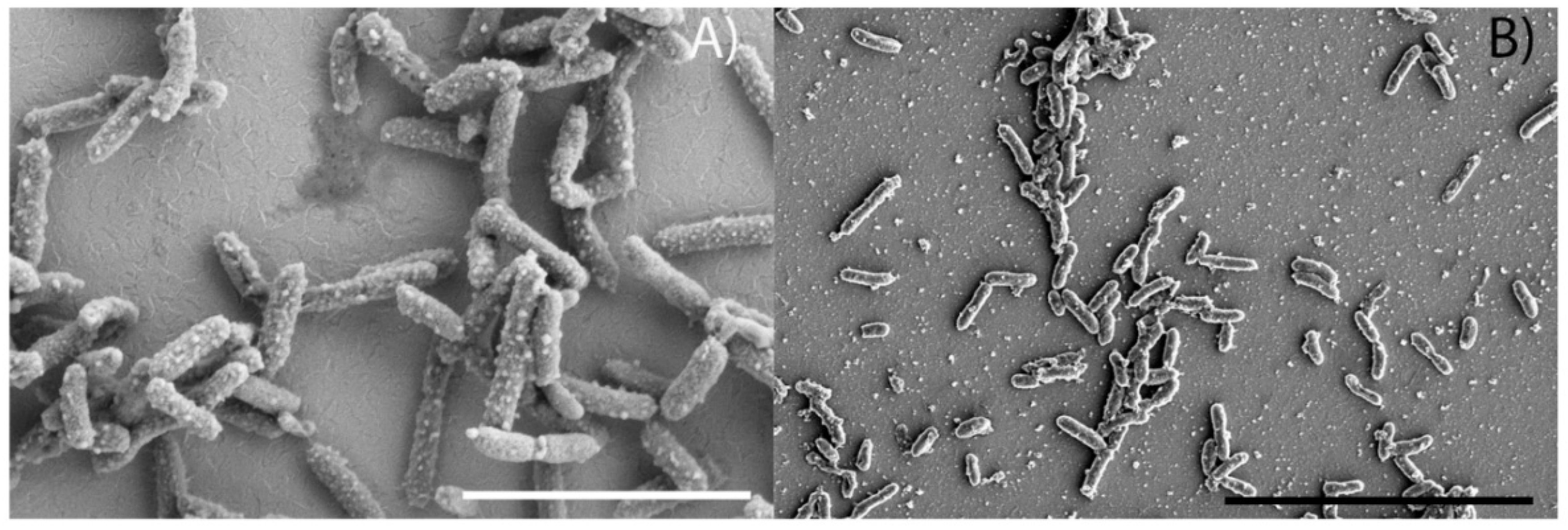
2.6. Scanning Electron Microscopy
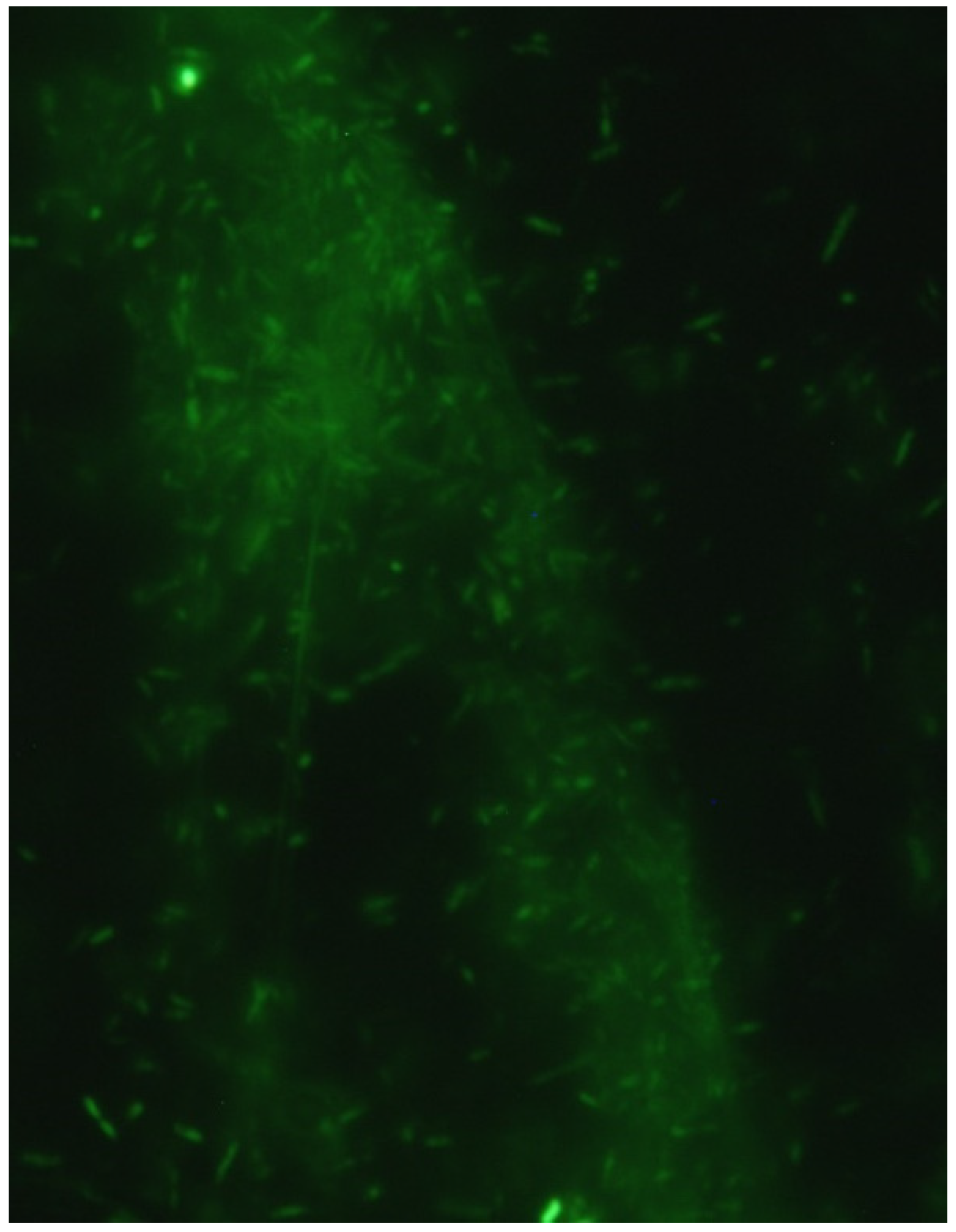
2.7. Fluorescence Microscopy
3. Results and Discussion
3.1. Lithoheterotrophic Isolates Fall within the Genus Idiomarina
3.2. Growth of Idiomarina Occurs Exclusively in the Presence of 11 Amino Acids
3.3. Streamlined Genome Supports a Narrow Environmental Niche for Growth
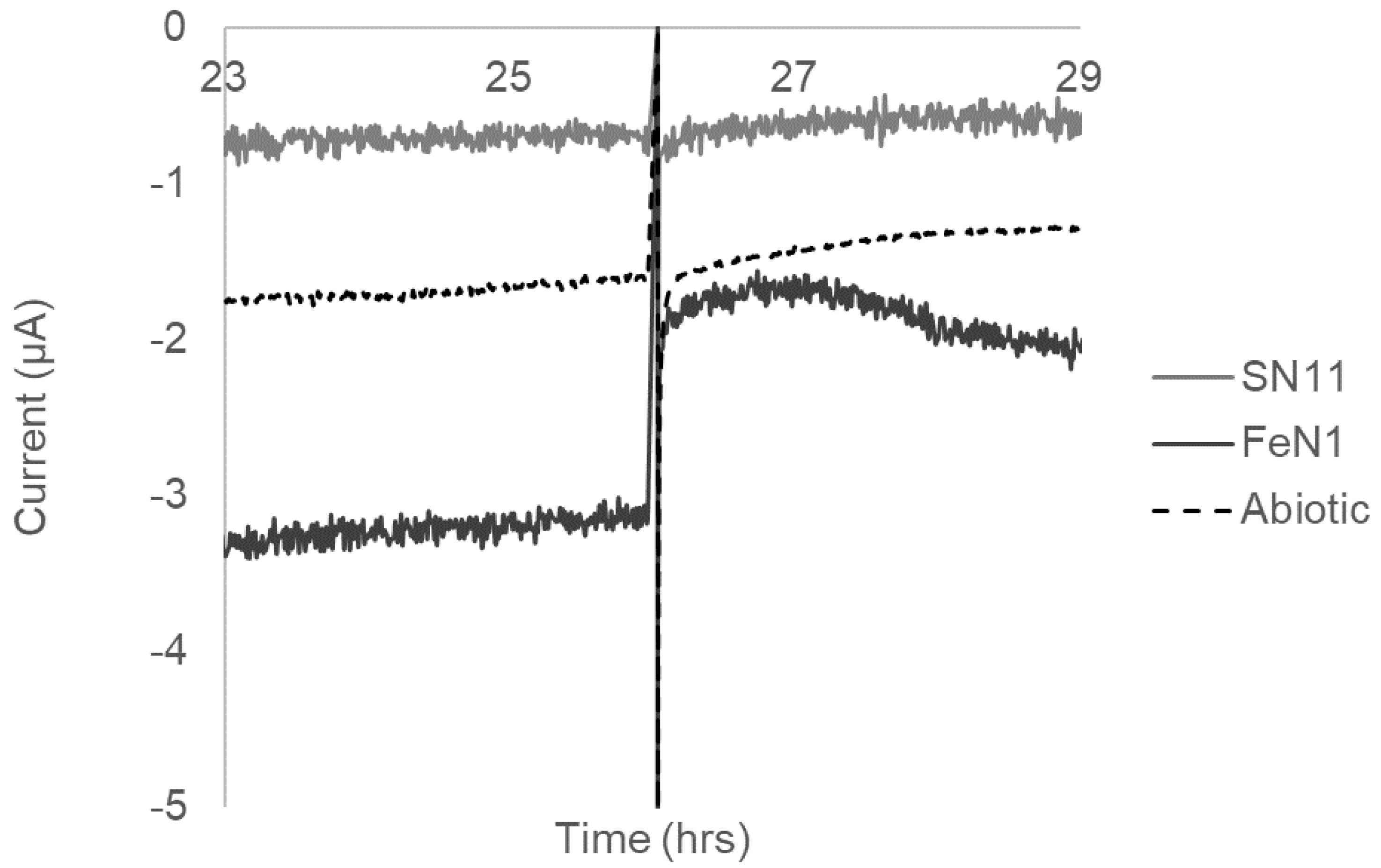
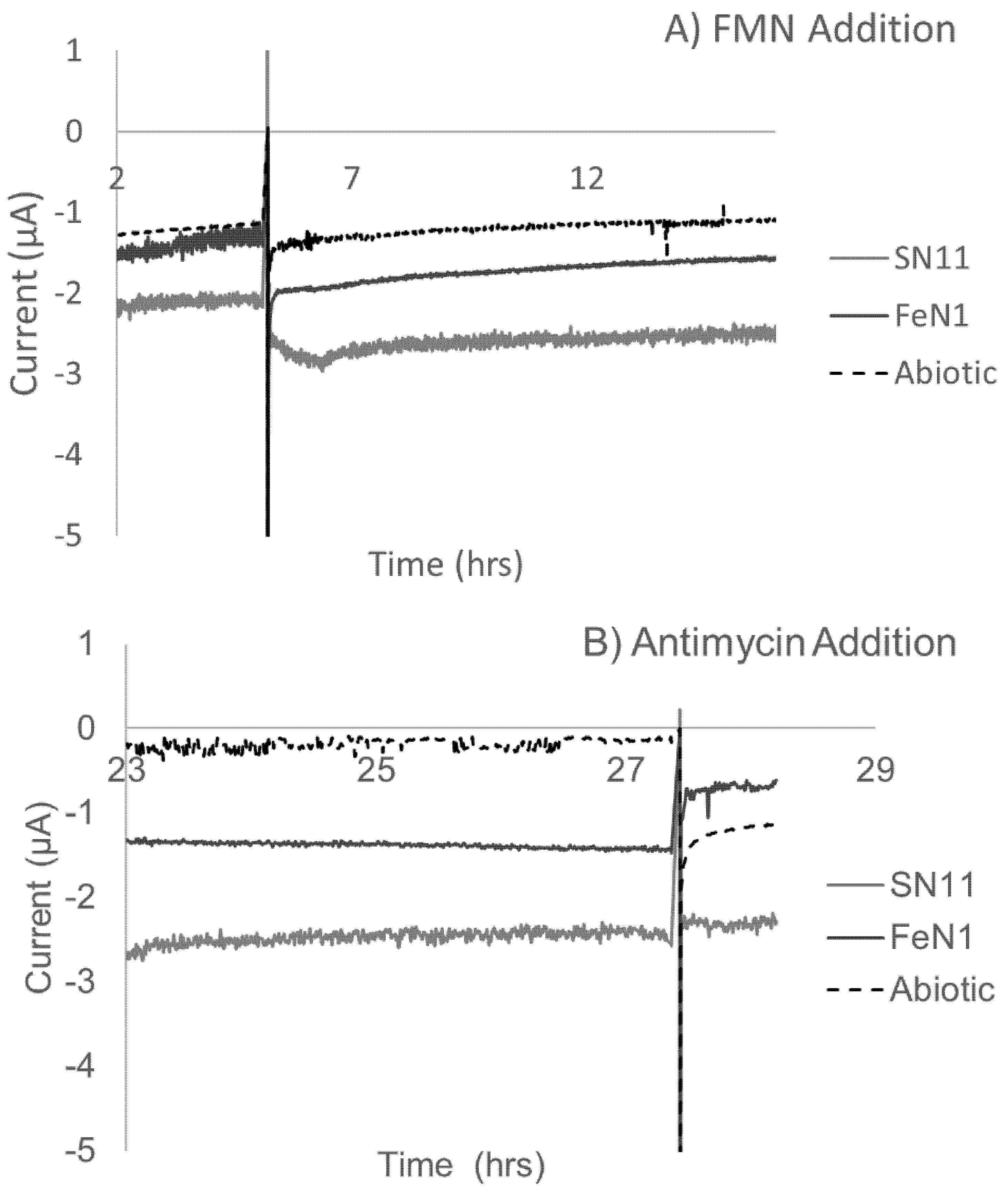
3.4. Electrochemical Activity Suggests That Electron Uptake Is Involved in Respiration (Not Growth)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ANI | 1 | 2 | 3 | 4 | 5 | |
|---|---|---|---|---|---|---|
| GGDC | ||||||
| 1 | 69.73% | 69.63% | 73.36% | 73.49% | ||
| 2 | 23.0% | 97.46% | 69.80% | 69.62% | ||
| 3 | 24.3% | 75.7% | 69.00% | 69.64% | ||
| 4 | 20.1% | 23.5% | 22.6% | 79.09% | ||
| 5 | 19.1% | 23.2% | 24.7% | 19.4% | ||



References
- Hau, H.H.; Gralnick, J.A. Ecology and Biotechnology of the Genus Shewanella. Annu. Rev. Microbiol. 2007, 61, 237–258. [Google Scholar] [CrossRef] [PubMed]
- Rabaey, K.; Rodríguez, J.; Blackall, L.L.; Keller, J.; Gross, P.; Batstone, D.; Verstraete, W.; Nealson, K.H. Microbial Ecology Meets Electrochemistry: Electricity-Driven and Driving Communities. ISME J. 2007, 1, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gralnick, J.A.; Newman, D.K. Extracellular Respiration. Mol. Microbiol. 2007, 65, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Dong, H.; Reguera, G.; Beyenal, H.; Lu, A.; Liu, J.; Yu, H.-Q.; Fredrickson, J.K. Extracellular Electron Transfer Mechanisms between Microorganisms and Minerals. Nat. Rev. Microbiol. 2016, 14, 651–662. [Google Scholar] [CrossRef]
- He, S.; Barco, R.A.; Emerson, D.; Roden, E.E. Comparative Genomic Analysis of Neutrophilic Iron(II) Oxidizer Genomes for Candidate Genes in Extracellular Electron Transfer. Front. Microbiol. 2017, 8, 1548. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, Z.; Belchik, S.M.; Edwards, M.J.; Liu, C.; Kennedy, D.W.; Merkley, E.D.; Lipton, M.S.; Butt, J.N.; Richardson, D.J.; et al. Identification and Characterization of MtoA: A Decaheme c-Type Cytochrome of the Neutrophilic Fe(II)-Oxidizing Bacterium Sideroxydans lithotrophicus ES-1. Front. Microbiol. 2012, 3, 37. [Google Scholar] [CrossRef] [Green Version]
- Keffer, J.L.; McAllister, S.M.; Garber, A.I.; Hallahan, B.J.; Sutherland, M.C.; Rozovsky, S.; Chan, C.S. Iron Oxidation by a Fused Cytochrome-Porin Common to Diverse Iron-Oxidizing Bacteria. MBio 2021, 12, e01074-21. [Google Scholar] [CrossRef]
- Barco, R.A.; Emerson, D.; Sylvan, J.B.; Orcutt, B.N.; Jacobson Meyers, M.E.; Ramírez, G.A.; Zhong, J.D.; Edwards, K.J. New Insight into Microbial Iron Oxidation as Revealed by the Proteomic Profile of an Obligate Iron-Oxidizing Chemolithoautotroph. Appl. Environ. Microbiol. 2015, 81, 5927–5937. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Dohmae, N.; Nealson, K.H.; Hashimoto, K.; Okamoto, A. Multi-Heme Cytochromes Provide a Pathway for Survival in Energy-Limited Environments. Sci. Adv. 2018, 4, eaao5682. [Google Scholar] [CrossRef] [Green Version]
- Scheller, S.; Yu, H.; Chadwick, G.L.; Mcglynn, S.E. Artificial Electron Acceptors Decouple Archaeal Methane Oxidation from Sulfate Reduction. Science 2016, 351, 1754–1756. [Google Scholar] [CrossRef] [Green Version]
- Mcglynn, S.E.; Chadwick, G.L.; Kempes, C.P.; Orphan, V.J. Single Cell Activity Reveals Direct Electron Transfer in Methanotrophic Consortia. Nature 2015, 526, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Shaw, D.R.; Ali, M.; Katuri, K.P.; Gralnick, J.A.; Van Niftrik, L.; Jetten, M.S.M.; Saikaly, P.E.; Reimann, J.; Mesman, R. Extracellular Electron Transfer-Depenent Anaerobic Oxidation of Ammonium by Anammox Bacteria. Nat. Commun. 2020, 11, 2058. [Google Scholar] [CrossRef] [PubMed]
- Rowe, A.R.; Chellamuthu, P.; Lam, B.; Okamoto, A.; Nealson, K.H. Marine Sediments Microbes Capable of Electrode Oxidation as a Surrogate for Lithotrophic Insoluble Substrate Metabolism. Front. Microbiol. 2015, 6, 784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, E.P.; Romanenko, L.A.; Chun, J.; Matte, M.H.; Matte, G.R.; Mikhailov, V.V.; Svetashev, V.I.; Huq, A.; Maugel, T.; Colwell, R.R. Idiomarina Gen. Nov., Comprising Novel Indigenous Deep-Sea Bacteria from the Pacific Ocean, Including Descriptions of Two Species, Idiomarina abyssalis Sp. Nov. and Idiomarina zobellii Sp. Nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Donachie, S.P.; Hou, S.; Gregory, T.S.; Malahoff, A.; Alam, M. Idiomarina loihiensis Sp. Nov., a Halophilic -Proteobacterium from the Lo’ihi Submarine Volcano, Hawai’i. Int. J. Syst. Evol. Microbiol. 2003, 53, 1873–1879. [Google Scholar] [CrossRef] [Green Version]
- Sitdhipol, J.; Visessanguan, W.; Benjakul, S.; Yukphan, P.; Tanasupawat, S. Idiomarina Piscisalsi Sp. Nov., from Fermented Fish (Pla-Ra) in Thailand. J. Gen. Appl. Microbiol. 2013, 59, 385–391. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.W.; Kim, B.Y.; Weon, H.Y.; Baek, Y.K.; Koo, B.S.; Go, S.J. Idiomarina homiensis Sp. Nov., Isolated from Seashore Sand in Korea. Int. J. Syst. Evol. Microbiol. 2006, 56, 2229–2233. [Google Scholar] [CrossRef]
- Chen, M.H.; Sheu, S.Y.; Chen, C.A.; Wang, J.T.; Chen, W.M. Idiomarina aquimaris Sp. Nov., Isolated from the Reef-Building Coral Isopora Palifera. Int. J. Syst. Evol. Microbiol. 2012, 62, 1536–1542. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Zhang, X.Y.; Zhao, H.L.; Zhou, M.Y.; Li, H.J.; Gao, Z.M.; Chen, X.L.; Dang, H.Y.; Zhang, Y.Z. Idiomarina maris Sp. Nov., a Marine Bacterium Isolated from Sediment. Int. J. Syst. Evol. Microbiol. 2012, 62, 370–375. [Google Scholar] [CrossRef] [Green Version]
- Erable, B.; Vandecandelaere, I.; Faimali, M.; Delia, M.L.; Etcheverry, L.; Vandamme, P.; Bergel, A. Marine Aerobic Biofilm as Biocathode Catalyst. Bioelectrochemistry 2010, 78, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Chang, R.; Bird, L.; Barr, C.; Osburn, M.; Wilbanks, E.; Nealson, K.; Rowe, A. Thioclava electrotropha Sp. Nov., a Versatile Electrode and Sulfur-Oxidizing Bacterium from Marine Sediments. Int. J. Syst. Evol. Microbiol. 2018, 68, 1652–1658. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate High-Throughput Multiple Sequence Alignment of Ribosomal RNA Genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- De Coster, W.; Hert, S.D.; Schultz, D.T.; Cruts, M.; Van Broeckhoven, C. Sequence Analysis NanoPack: Visualizing and Processing Long-Read Sequencing Data. Bioinformatics 2018, 34, 2666–2669. [Google Scholar] [CrossRef] [PubMed]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of Long, Error-Prone Reads Using Repeat Graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and Accurate Long-Read Alignment with Burrows–Wheeler Transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. Genome Analysis QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Tatusova, T.; Dicuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI Prokaryotic Genome Annotation Pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, O.S.; Cho, Y.J.; Lee, K.; Yoon, S.H.; Kim, M.; Na, H.; Park, S.C.; Jeon, Y.S.; Lee, J.H.; Yi, H.; et al. Introducing EzTaxon-e: A Prokaryotic 16s RRNA Gene Sequence Database with Phylotypes That Represent Uncultured Species. Int. J. Syst. Evol. Microbiol. 2012, 62, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome Sequence-Based Species Delimitation with Confidence Intervals and Improved Distance Functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, C.E.; Bailey, T.L.; Noble, W.S. FIMO: Scanning for Occurrences of a given Motif. Bioinformatics 2011, 27, 1017–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garber, A.I.; Nealson, K.H.; Okamoto, A.; McAllister, S.M.; Chan, C.S.; Barco, R.A.; Merino, N. FeGenie: A Comprehensive Tool for the Identification of Iron Genes and Iron Gene Neighborhoods in Genome and Metagenome Assemblies. Front. Microbiol. 2020, 11, 37. [Google Scholar] [CrossRef] [Green Version]
- Rowe, A.R.; Salimijazi, F.; Trutschel, L.; Sackett, J.; Adesina, O.; Anzai, I.; Kugelmass, L.H.; Baym, M.H.; Barstow, B. Identification of a Pathway for Electron Uptake in Shewanella oneidensis. Commun. Biol. 2021, 4, 957. [Google Scholar] [CrossRef]
- Weber, W., Jr.; Pirbazari, M.; Melson, G. Biological Growth on Activated Carbon: An Investigation by Scanning Electron Microscopy. Environ. Sci. Technol. 1978, 12, 817–819. [Google Scholar] [CrossRef]
- Rowe, A.R.; Rajeev, P.; Jain, A.; Pirbadian, S.; Okamoto, A.; Gralnick, J.A.; El-Naggar, M.Y.; Nealson, K.H. Tracking Electron Uptake from a Cathode into Shewanella Cells: Implications for Energy Acquisition from Solid-Substrate Electron Donors. mBio 2018, 9, e02203-17. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.H.; Shen, Y.Q.; Xu, X.W.; Wang, C.S.; Oren, A.; Wu, M. Pseudidiomarina donghaiensis Sp. Nov. and Pseudidiomarina maritima Sp. Nov., Isolated from the East China Sea. Int. J. Syst. Evol. Microbiol. 2009, 59, 1321–1325. [Google Scholar] [CrossRef] [Green Version]
- Hou, S.; Saw, J.H.; Lee, K.S.; Freitas, T.A.; Belisle, C.; Kawarabayasi, Y.; Donachie, S.P.; Pikina, A.; Galperin, M.Y.; Koonin, E.V.; et al. Genome Sequence of the Deep-Sea γ-Proteobacterium Idiomarina loihiensis Reveals Amino Acid Fermentation as a Source of Carbon and Energy. Proc. Natl. Acad. Sci. USA 2004, 101, 18036–18041. [Google Scholar] [CrossRef] [Green Version]
- Jisha, V.N.; Smitha, R.B.; Pradeep, S.; Sreedevi, S.; Unni, K.N.; Sajith, S.; Priji, P.; Sarath Josh, M.; Benjamin, S. Versatility of Microbial Proteases. Adv. Enzym. Res. 2013, 1, 39–51. [Google Scholar] [CrossRef]
- Saunders, S.H.; Tse, E.C.M.; Yates, M.D.; Otero, F.J.; Trammell, S.A.; Stemp, E.D.A.; Barton, J.K.; Tender, L.M.; Newman, D.K. Extracellular DNA Promotes Efficient Extracellular Electron Transfer by Pyocyanin in Pseudomonas aeruginosa Biofilms. Cell 2020, 182, 919–932.e19. [Google Scholar] [CrossRef] [PubMed]
- Kaden, J.; Galushko, A.S.; Schink, B. Cysteine-Mediated Electron Transfer in Syntrophic Acetate Oxidation by Cocultures of Geobacter sulfurreducens and Wolinella succinogenes. Arch. Microbiol. 2002, 178, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Jangir, Y.; El-Naggar, M.Y. Disentangling the Roles of Free and Cytochrome-Bound Flavins in Extracellular Electron Transport from Shewanella oneidensis MR-1. Electrochim. Acta 2016, 198, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Light, S.H.; Su, L.; Rivera-Lugo, R.; Cornejo, J.A.; Louie, A.; Iavarone, A.T.; Ajo-Franklin, C.M.; Portnoy, D.A. A Flavin-Based Extracellular Electron Transfer Mechanism in Diverse Gram-Positive Bacteria. Nature 2018, 562, 140–157. [Google Scholar] [CrossRef] [PubMed]



| Characteristics | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| Cell length (µm) | 1.0–2.0 | 1.0–2.0 | 0.7–1.8 1 | 1.0–1.4 2 | 1.4–2.0 2 |
| Temperature ranges (°C) | 4–40 | 4–40 | 6–39 | 7–42.5 | 7–45 |
| Optimum temperature (°C) | 35–40 | 25–30 | nt | nt | nt |
| NaCl ranges (%, w/v) | 0.1–18 | 0.1–18 | 0.25–17.5 | 0.1–12.5 | 0–13 |
| pH ranges | 6.1–10 | 6.1–9.4 | 6.5–9.4 | 6.1–10 | 6.5–9.4 |
| G + C content (mol.%) | 48.15 | 47.02 | 47.04 | 48.36 | 47.22 |
| Metabolic Tests | |||||
| Catalase | + | + | + | + | + |
| Oxidase | + | + | + | + | + |
| Hydrolysis of protein (gelatin) | + | − | + | + | − |
| Fatty Acid | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| C11:0 iso | 1.9 | 2.3 | 2.2 | 2.44 | 1.9 |
| C11:0 iso 3OH | 4.1 | 4.1 | 4.5 | 4.26 | 4.7 |
| c13:0 iso | 1.6 | 2.2 | 1.6 | 3.38 | 2.1 |
| c13:0 iso 3OH | 3.8 | 3.8 | 3.6 | 3.39 | 3.7 |
| c15:1 iso F | 1.8 | 3.2 | 1.6 | 5.42 | 5.9 |
| c15:0 iso | 26.0 | 38.4 | 30.4 | 36.82 | 32.8 |
| c16:0 | 7.4 | 7.2 | 1.1 | 2.88 | 3.8 |
| c17:0 | 2.0 | 1.5 | 1.3 | 0.35 | 5.2 |
| c17:0 iso | 15.0 | 11.8 | 17.1 | 13.2 | 14.2 |
| c17:1 iso | 11.5 | 12.0 | 11.6 | 9.53 | 14.0 |
| c18:0 | 2.7 | 1.1 | 1.8 | 2.22 | 2.0 |
| FeN1 | SN11 | |
|---|---|---|
| BioProject accession no. | PRJNA726532 | PRJNA726532 |
| Assembly accession no. | CP074112 | CP074073 |
| Assembly size (bp) | 2,900,277 | 2,834,848 |
| Estimated genome completeness (%) 1 | 99.66% | 100% |
| Estimated contamination (%) 1 | 0.9% | 0.17% |
| No. of contigs | 1 | 1 |
| No. of protein-coding genes | 2667 | 2616 |
| No. of tRNA genes | 57 | 56 |
| No. of rRNA operons | 4 | 4 |
| SN11 Locus Tag | FeN1 Locus Tag | Annotation | Query Coverage/Percentage Identity 1 | Localization (Score) |
|---|---|---|---|---|
| KF946_02970 | KGF88_02385 | Cytochrome c | 98/54 | Periplasmic (10.00) |
| KF946_04255 | KGF88_00295 | NAD(P)-dependent alcohol dehydrogenase | 99/61 | Cytoplasmic (9.97) |
| KF946_04965 | KGF88_04300 | Cytochrome-c oxidase, cbb3-type subunit III ccoP | 99/74 | Unknown |
| KF946_04975 | KGF88_04310 | Cytochrome-c oxidase, cbb3-type subunit II ccoO | 97/85 | Cytoplasmic (8.96) |
| KF946_05620 | 4a-Hydroxytetrahydrobiopterin dehydratase | Cytoplasmic (9.26) | ||
| KF946_05805 | KGF88_01080 | Cytochrome c | 94/60 | Unknown/periplasmic (9.84) |
| KF946_05965 | DUF3179 domain-containing protein | Cytoplasmic membrane (10.00) | ||
| KF946_06450 | KGF88_06085 | 2Fe–2S iron–sulfur cluster binding domain-containing protein | 100/67 | Unknown |
| KF946_06970 | KGF88_07065 | Molecular chaperone DnaJ | 99/79 | Cytoplasmic (9.97) |
| KF946_09505 | KGF88_09595 | Cytochrome c1 | 100/73 | Unknown—may have multiple localization sites |
| KF946_10365 | KGF88_10235 | Cytochrome c oxidase subunit II coxB | 98/74 | Cytoplasmic membrane (9.99) |
| KF946_10655 | KGF88_10495 | Cytochrome c5 family protein | 88/59 | Unknown |
| KF946_10745 | KGF88_12320 | Anaerobic ribonucleoside-triphosphate reductase activating protein nrdG | 92/72 | Cytoplasmic (9.26) |
| KF946_11095 | Cytochrome c/FTR1 family iron permease | Cytoplasmic membrane (10.00) | ||
| KF946_11525 | KGF88_11325 | Cytochrome c4 | 100/64 | Periplasmic (10.00) |
| KF946_12560 | KGF88_12535 | 50S ribosomal protein L31 rpmE | 97/70 | Cytoplasmic (9.26) |
| KF946_13430 | EAL and GGDEF domain-containing protein | Cytoplasmic membrane (7.88) | ||
| KGF88_01265 | dsbC family protein | Periplasmic (9.76) | ||
| KGF88_01425 | Thioredoxin family protein | Unknown—may have multiple localization sites | ||
| KGF88_01810 | Cryptochrome/photolyase family protein | Unknown | ||
| KGF88_05965 | Thioredoxin trxC | Cytoplasmic (9.26) | ||
| KGF88_07240 | Flp pilus assembly complex ATPase component TadA | Cytoplasmic (9.97) | ||
| KGF88_09010 | ISC system 2Fe-2S type ferredoxin | Cytoplasmic (8.96) | ||
| KGF88_09905 | Exinuclease ABC subunit UvrA | Cytoplasmic (9.97) | ||
| KGF88_10910 | Catalase | Periplasmic (10.00) | ||
| KGF88_11150 | Cytochrome c peroxidase | Periplasmic (10.00) | ||
| KGF88_11220 | DUF3365 domain-containing protein | Periplasmic (9.84) | ||
| KGF88_13155 | C-type cytochrome | Cytoplasmic membrane (10.00) | ||
| KGF88_13170 | C-type cytochrome | Periplasmic (10.00) | ||
| KGF88_13175 | C-type cytochrome | Periplasmic (10.00) | ||
| KGF88_13495 | Rhodanese-related sulfurtransferase | Cytoplasmic (8.96) | ||
| KGF88_13780 | C-type cytochrome | Periplasmic (10.00) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vinales, J.; Sackett, J.; Trutschel, L.; Amir, W.; Norman, C.; Leach, E.; Wilbanks, E.; Rowe, A. Physiologic, Genomic, and Electrochemical Characterization of Two Heterotrophic Marine Sediment Microbes from the Idiomarina Genus. Microorganisms 2022, 10, 1219. https://doi.org/10.3390/microorganisms10061219
Vinales J, Sackett J, Trutschel L, Amir W, Norman C, Leach E, Wilbanks E, Rowe A. Physiologic, Genomic, and Electrochemical Characterization of Two Heterotrophic Marine Sediment Microbes from the Idiomarina Genus. Microorganisms. 2022; 10(6):1219. https://doi.org/10.3390/microorganisms10061219
Chicago/Turabian StyleVinales, Jorge, Joshua Sackett, Leah Trutschel, Waleed Amir, Casey Norman, Edmund Leach, Elizabeth Wilbanks, and Annette Rowe. 2022. "Physiologic, Genomic, and Electrochemical Characterization of Two Heterotrophic Marine Sediment Microbes from the Idiomarina Genus" Microorganisms 10, no. 6: 1219. https://doi.org/10.3390/microorganisms10061219
APA StyleVinales, J., Sackett, J., Trutschel, L., Amir, W., Norman, C., Leach, E., Wilbanks, E., & Rowe, A. (2022). Physiologic, Genomic, and Electrochemical Characterization of Two Heterotrophic Marine Sediment Microbes from the Idiomarina Genus. Microorganisms, 10(6), 1219. https://doi.org/10.3390/microorganisms10061219