
Transmembrane Protein LMxysn_1693 of Serovar 4h Listeria monocytogenes Is Associated with Bile Salt Resistance and Intestinal Colonization

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Cell Lines and Animals
2.2. Bioinformatics Analysis
2.3. Mutant and Complemented Strains Construction
2.4. Evaluation of Biological Characteristics
2.5. Growth Curve Analysis
2.6. Determination of Biofilm Formation
2.7. Analysis of Gene Expression at Transcriptional Level
2.8. Adhesion, Invasion and Replication Capacity of Lm
2.9. Infection to Mice
2.10. Date Analysis
3. Results
3.1. Association with the Transporter System
3.2. Involvement in Resisting Bile Salt
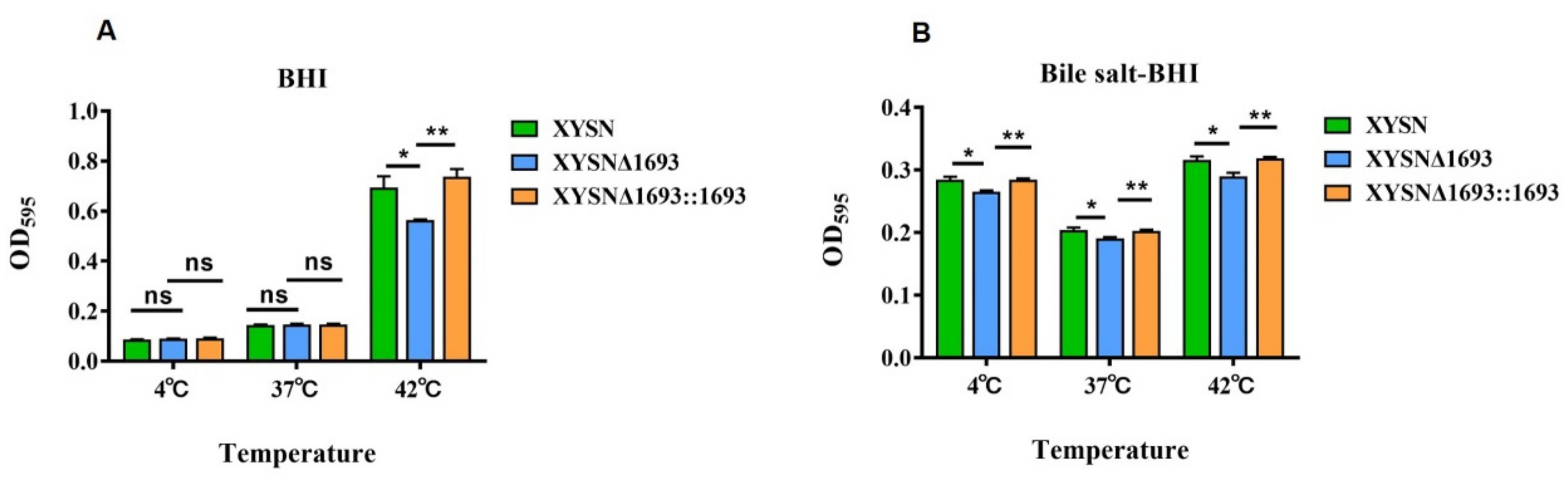
3.3. Contribution to Biofilm Formation
3.4. Interaction with Virulence and Bile-Salt-Resistant Genes
3.5. Enhancement Adhesion and Invasion Capacity
3.6. Promotion of Listeria Colonization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, S.; Meng, F.; Sun, X.; Yao, H.; Wang, Y.; Pan, Z.; Yin, Y.; Jiao, X. Epidemiology of human Listeriosis in China during 2008–2017. Foodborne Pathog. Dis. 2020, 17, 119–125. [Google Scholar] [CrossRef]
- Chawla, C.S.; Chen, H.; Donnelly, C.W. Mathematically modeling the repair of heat-injured Listeria monocytogenes as affected by temperature, pH, and salt concentration. Int. J. Food Microbiol. 1996, 30, 231–242. [Google Scholar] [CrossRef]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Núñez-Montero, K.; Leclercq, A.; Moura, A.; Vales, G.; Peraza, J.; Pizarro-Cerdá, J.; Lecuit, M. Listeria costaricensis sp. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 844–850. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Chen, Y.; Gorski, L.; Ward, T.J.; Osborne, J.; Kathariou, S. Listeria monocytogenes source distribution analysis indicates regional heterogeneity and ecological niche preference among serotype 4b clones. MBio 2018, 9, e00396-18. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Yao, H.; Doijad, S.; Kong, S.; Shen, Y.; Cai, X.; Tan, W.; Wang, Y.; Feng, Y.; Ling, Z.; et al. A hybrid sub-lineage of Listeria monocytogenes comprising hypervirulent isolates. Nat. Commun. 2019, 10, 4283. [Google Scholar] [CrossRef] [Green Version]
- Sheet, S.; Yesupatham, S.; Ghosh, K.; Choi, M.S.; Shim, K.S.; Lee, Y.S. Modulatory effect of low-shear modeled microgravity on stress resistance, membrane lipid composition, virulence, and relevant gene expression in the food-borne pathogen Listeria monocytogenes. Enzyme Microb. Technol. 2020, 133, 109440. [Google Scholar] [CrossRef]
- Cotter, P.D.; Gahan, C.G.; Hill, C. A glutamate decarboxylase system protects Listeria monocytogenes in gastric fluid. Mol. Microbiol. 2001, 40, 465–475. [Google Scholar] [CrossRef]
- Sebald, W.; Friedl, P.; Schairer, H.U.; Hoppe, J. Structure and genetics of the H+-conducting F0 portion of the ATP synthase. Ann. N. Y. Acad. Sci. 1982, 402, 28–44. [Google Scholar] [CrossRef] [Green Version]
- Chaturongakul, S.; Raengpradub, S.; Palmer, M.E.; Bergholz, T.M.; Orsi, R.H.; Hu, Y.; Ollinger, J.; Wiedmann, M.; Boor, K.J. Transcriptomic and phenotypic analyses identify coregulated, overlapping regulons among PrfA, CtsR, HrcA, and the alternative sigma factors sigmaB, sigmaC, sigmaH, and sigmaL in Listeria monocytogenes. Appl. Environ. Microbiol. 2011, 77, 187–200. [Google Scholar] [CrossRef] [Green Version]
- Begley, M.; Sleator, R.D.; Gahan, C.G.; Hill, C. Contribution of three bile-associated loci, bsh, pva, and btlB, to gastrointestinal persistence and bile tolerance of Listeria monocytogenes. Infect. Immun. 2005, 73, 894–904. [Google Scholar] [CrossRef] [Green Version]
- Pombinho, R.; Vieira, A.; Camejo, A.; Archambaud, C.; Cossart, P.; Sousa, S.; Cabanes, D. Virulence gene repression promotes Listeria monocytogenes systemic infection. Gut Microbes 2020, 11, 868–881. [Google Scholar] [CrossRef]
- Kovačević, J.; Mesak, L.R.; Allen, K.J. Occurrence and characterization of Listeria spp. in ready-to-eat retail foods from Vancouver, British Columbia. Food Microbiol. 2012, 30, 372–378. [Google Scholar] [CrossRef]
- Lee, S.; Parsons, C.; Chen, Y.; Hanafy, Z.; Brown, E.; Kathariou, S. Identification and characterization of a novel genomic island harboring cadmium and arsenic resistance genes in Listeria welshimeri. Biomolecules 2021, 11, 560. [Google Scholar] [CrossRef]
- Hochhut, B.; Jahreis, K.; Lengeler, J.W.; Schmid, K. CTnscr94, a conjugative transposon found in enterobacteria. J. Bacteriol. 1997, 179, 2097–2102. [Google Scholar] [CrossRef] [Green Version]
- Park, S.F.; Stewart, G.S. High-efficiency transformation of Listeria monocytogenes by electroporation of penicillin-treated cells. Gene 1990, 94, 129–132. [Google Scholar] [CrossRef]
- NicAogáin, K.; O’Byrne, C.P. The role of stress and stress adaptations in determining the fate of the bacterial pathogen Listeria monocytogenes in the food chain. Front. Microbiol. 2016, 7, 1865. [Google Scholar] [CrossRef] [Green Version]
- Shi, X.; Zhu, X. Biofilm formation and food safety in food industries. Trends Food Sci. Technol. 2009, 20, 407–413. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, L.; Han, J.; Dong, P.; Luo, X.; Zhang, Y.; Zhu, L. Inhibition of biofilm formation and related gene expression of Listeria monocytogenes in response to four natural antimicrobial compounds and sodium hypochlorite. Front. Microbiol. 2021, 11, 617473. [Google Scholar] [CrossRef]
- Weiler, C.; Ifland, A.; Naumann, A.; Kleta, S.; Noll, M. Incorporation of Listeria monocytogenes strains in raw milk biofilms. Int. J. Food Microbiol. 2013, 161, 61–68. [Google Scholar] [CrossRef]
- Dowd, G.C.; Joyce, S.A.; Hill, C.; Gahan, C.G. Investigation of the mechanisms by which Listeria monocytogenes grows in porcine gallbladder bile. Infect. Immun. 2011, 79, 369–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chand, D.; Avinash, V.S.; Yadav, Y.; Pundle, A.V.; Suresh, C.G.; Ramasamy, S. Molecular features of bile salt hydrolases and relevance in human health. Biochim. Biophys. Acta. Gen. Subj. 2017, 1861, 2981–2991. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, K.T.; Carleton, J.D.; Quillin, S.J.; Rollins, S.D.; Portnoy, D.A.; Leber, J.H. Hyperinduction of host beta interferon by a Listeria monocytogenes strain naturally overexpressing the multidrug efflux pump MdrT. Infect. Immun. 2012, 80, 1537–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Feng, Y.; Deng, L.; Feng, F.; Wang, L.; Zhou, Q.; Luo, Q. SigB plays a major role in Listeria monocytogenes tolerance to bile stress. Int. J. Food Microbiol. 2011, 145, 238–243. [Google Scholar] [CrossRef]
- Dussurget, O.; Cabanes, D.; Dehoux, P.; Lecuit, M.; Buchrieser, C.; Glaser, P.; Cossart, P.; European Listeria Genome Consortium. Listeria monocytogenes bile salt hydrolase is a PrfA-regulated virulence factor involved in the intestinal and hepatic phases of listeriosis. Mol. Microbiol. 2002, 45, 1095–1106. [Google Scholar]
- Sleator, R.D.; Wemekamp-Kamphuis, H.H.; Gahan, C.G.; Abee, T.; Hill, C. A PrfA-regulated bile exclusion system (BilE) is a novel virulence factor in Listeria monocytogenes. Mol. Microbiol. 2005, 55, 1183–1195. [Google Scholar] [CrossRef]
- Quillin, S.J.; Schwartz, K.T.; Leber, J.H. The novel Listeria monocytogenes bile sensor BrtA controls expression of the cholic acid efflux pump MdrT. Mol. Microbiol. 2011, 81, 129–142. [Google Scholar] [CrossRef]
- Vazquez-Armenta, F.J.; Hernandez-Oñate, M.A.; Martinez-Tellez, M.A.; Lopez-Zavala, A.A.; Gonzalez-Aguilar, G.A.; Gutierrez-Pacheco, M.M.; Ayala-Zavala, J.F. Quercetin repressed the stress response factor (sigB) and virulence genes (prfA, actA, inlA, and inlC), lower the adhesion, and biofilm development of L. monocytogenes. Food Microbiol. 2020, 87, 103377. [Google Scholar] [CrossRef]
- Lemon, K.P.; Freitag, N.E.; Kolter, R. The virulence regulator PrfA promotes biofilm formation by Listeria monocytogenes. J. Bacteriol. 2010, 192, 3969–3976. [Google Scholar] [CrossRef] [Green Version]
- Toledo-Arana, A.; Dussurget, O.; Nikitas, G.; Sesto, N.; Guet-Revillet, H.; Balestrino, D.; Loh, E.; Gripenland, J.; Tiensuu, T.; Vaitkevicius, K.; et al. The Listeria transcriptional landscape from saprophytism to virulence. Nature 2009, 459, 950–956. [Google Scholar] [CrossRef]
- Feng, Z.; Liu, D.; Wang, L.; Wang, Y.; Zang, Z.; Liu, Z.; Song, B.; Gu, L.; Fan, Z.; Yang, S.; et al. A putative efflux transporter of the ABC family, YbhFSR, in Escherichia coli functions in tetracycline efflux and Na+(Li+)/H+ transport. Front. Microbiol. 2020, 11, 556. [Google Scholar] [CrossRef] [PubMed]
- Benda, M.; Schulz, L.M.; Stülke, J.; Rismondo, J. Influence of the ABC transporter YtrBCDEF of Bacillus subtilis on competence, biofilm formation and cell wall thickness. Front. Microbiol. 2021, 12, 587035. [Google Scholar] [CrossRef] [PubMed]
- Poudyal, B.; Sauer, K. The ABC of biofilm drug tolerance: The MerR-like regulator BrlR is an activator of ABC transport systems, with PA1874-77 contributing to the tolerance of Pseudomonas aeruginosa biofilms to tobramycin. Antimicrob. Agents. Chemother. 2018, 62, e01981-17. [Google Scholar] [CrossRef] [PubMed]
- Pinweha, P.; Pumirat, P.; Cuccui, J.; Jitprasutwit, N.; Muangsombut, V.; Srinon, V.; Boonyuen, U.; Thiennimitr, P.; Vattanaviboon, P.; Cia, F.; et al. Inactivation of bpsl1039-1040 ATP-binding cassette transporter reduces intracellular survival in macrophages, biofilm formation and virulence in the murine model of Burkholderia pseudomallei infection. PLoS ONE 2018, 13, e0196202. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Geng, Y.; Ren, S.; Yu, T.; Li, Y.; Liu, G.; Wang, H.; Meng, H.; Shi, L. The VirAB-VirSR-AnrAB multicomponent system is involved in resistance of Listeria monocytogenes EGD-e to cephalosporins, bacitracin, nisin, benzalkonium chloride, and ethidium bromide. Appl. Environ. Microbiol. 2019, 85, e01470-19. [Google Scholar] [CrossRef]
- Grubaugh, D.; Regeimbal, J.M.; Ghosh, P.; Zhou, Y.; Lauer, P.; Dubensky, T.W., Jr.; Higgins, D.E. The VirAB ABC transporter is required for VirR regulation of Listeria monocytogenes virulence and resistance to nisin. Infect. Immun. 2018, 86, e00901-17. [Google Scholar] [CrossRef] [Green Version]
- Collins, B.; Curtis, N.; Cotter, P.D.; Hill, C.; Ross, R.P. The ABC transporter AnrAB contributes to the innate resistance of Listeria monocytogenes to nisin, bacitracin, and various beta-lactam antibiotics. Antimicrob. Agents Chemother. 2010, 54, 4416–4423. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; Wiedmann, M.; Boor, K.J.; Bergholz, T.M. VirR-mediated resistance of Listeria monocytogenes against food antimicrobials and cross-protection induced by exposure to organic acid salts. Appl. Environ. Microbiol. 2015, 81, 4553–4562. [Google Scholar] [CrossRef] [Green Version]
- Rismondo, J.; Schulz, L.M. Not just transporters: Alternative functions of ABC transporters in Bacillus subtilis and Listeria monocytogenes. Microorganisms 2021, 9, 163. [Google Scholar] [CrossRef]
- Matthysse, A.G.; Yarnall, H.A.; Young, N. Requirement for genes with homology to ABC transport systems for attachment and virulence of Agrobacterium tumefaciens. J. Bacteriol. 1996, 178, 5302–5308. [Google Scholar] [CrossRef] [Green Version]
- Soni, D.K.; Dubey, S.K.; Bhatnagar, R. ATP-binding cassette (ABC) import systems of Mycobacterium tuberculosis: Target for drug and vaccine development. Emerg. Microbes. Infect. 2020, 9, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Locher, K.P. Mechanistic diversity in ATP-binding cassette (ABC) transporters. Nat. Struct. Mol. Biol. 2016, 23, 487–493. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Domain | Location | Functional Prediction |
|---|---|---|---|
| LMxysn_1677 | None | Outside | exonuclease SbcD |
| LMxysn_1678 | PlsC Domain | Outside | 1-acyl-sn-glycerol-3-phosphate acyltransferase |
| LMxysn_1679 | Transmembrane Region | TM helix | Lipoprotein |
| LMxysn_1680 | Transmembrane Region | TM helix | Hypothetical Protein |
| LMxysn_1681 | AAA Domain | TM helix | ABC transporter ATP-binding protein/permease |
| LMxysn_1682 | AAA Domain | TM helix | ABC transporter ATP-binding protein/permease |
| LMxysn_1683 | Transmembrane Region | TM helix | Cellsurface protein |
| LMxysn_1684 | None | Outside | Proteolysis |
| LMxysn_1685 | None | Inside | Hypothetical Protein |
| LMxysn_1686 | Transmembrane Region | TM helix | Hypothetical Protein |
| LMxysn_1687 | None | Inside | Hypothetical Protein |
| LMxysn_1688 | Low complexity | Outside | DUF3221 Domain-containing Protein |
| LMxysn_1689 | None | Inside | Hypothetical Protein |
| LMxysn_1690 | Low complexity | Outside | Hypothetical Protein |
| LMxysn_1691 | Transmembrane Region | TM helix | Hypothetical Protein |
| LMxysn_1692 | Transmembrane Region | TM helix | Multisubunit Na+/H+ antiporter |
| LMxysn_1693 | Transmembrane Region | TM helix | Hypothetical Protein |
| LMxysn_1694 | None | Outside | Hypothetical Protein |
| LMxysn_1695 | SH3B Domain | Inside | N-acetylmuramoyl-L-alanine amidase |
| LMxysn_1696 | Phage_integrase | Outside | Belongs to the ‘phage’ integrase family |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, F.; Feng, Y.; Chen, C.; Yao, H.; Zhang, R.; Zhang, Q.; Meng, F.; Chen, X.; Jiao, X.; Yin, Y. Transmembrane Protein LMxysn_1693 of Serovar 4h Listeria monocytogenes Is Associated with Bile Salt Resistance and Intestinal Colonization. Microorganisms 2022, 10, 1263. https://doi.org/10.3390/microorganisms10071263
Jin F, Feng Y, Chen C, Yao H, Zhang R, Zhang Q, Meng F, Chen X, Jiao X, Yin Y. Transmembrane Protein LMxysn_1693 of Serovar 4h Listeria monocytogenes Is Associated with Bile Salt Resistance and Intestinal Colonization. Microorganisms. 2022; 10(7):1263. https://doi.org/10.3390/microorganisms10071263
Chicago/Turabian StyleJin, Fanxin, Youwei Feng, Chao Chen, Hao Yao, Renling Zhang, Qin Zhang, Fanzeng Meng, Xiang Chen, Xin’an Jiao, and Yuelan Yin. 2022. "Transmembrane Protein LMxysn_1693 of Serovar 4h Listeria monocytogenes Is Associated with Bile Salt Resistance and Intestinal Colonization" Microorganisms 10, no. 7: 1263. https://doi.org/10.3390/microorganisms10071263
APA StyleJin, F., Feng, Y., Chen, C., Yao, H., Zhang, R., Zhang, Q., Meng, F., Chen, X., Jiao, X., & Yin, Y. (2022). Transmembrane Protein LMxysn_1693 of Serovar 4h Listeria monocytogenes Is Associated with Bile Salt Resistance and Intestinal Colonization. Microorganisms, 10(7), 1263. https://doi.org/10.3390/microorganisms10071263