
Genome-Based Taxonomy of the Genus Stutzerimonas and Proposal of S. frequens sp. nov. and S. degradans sp. nov. and Emended Descriptions of S. perfectomarina and S. chloritidismutans


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Genome Sequencing
2.3. Phylogeny and Species Genomic Indices
2.4. Phenotypic Characterization
3. Results
3.1. Genome Characteristics of the New Sequenced Strains
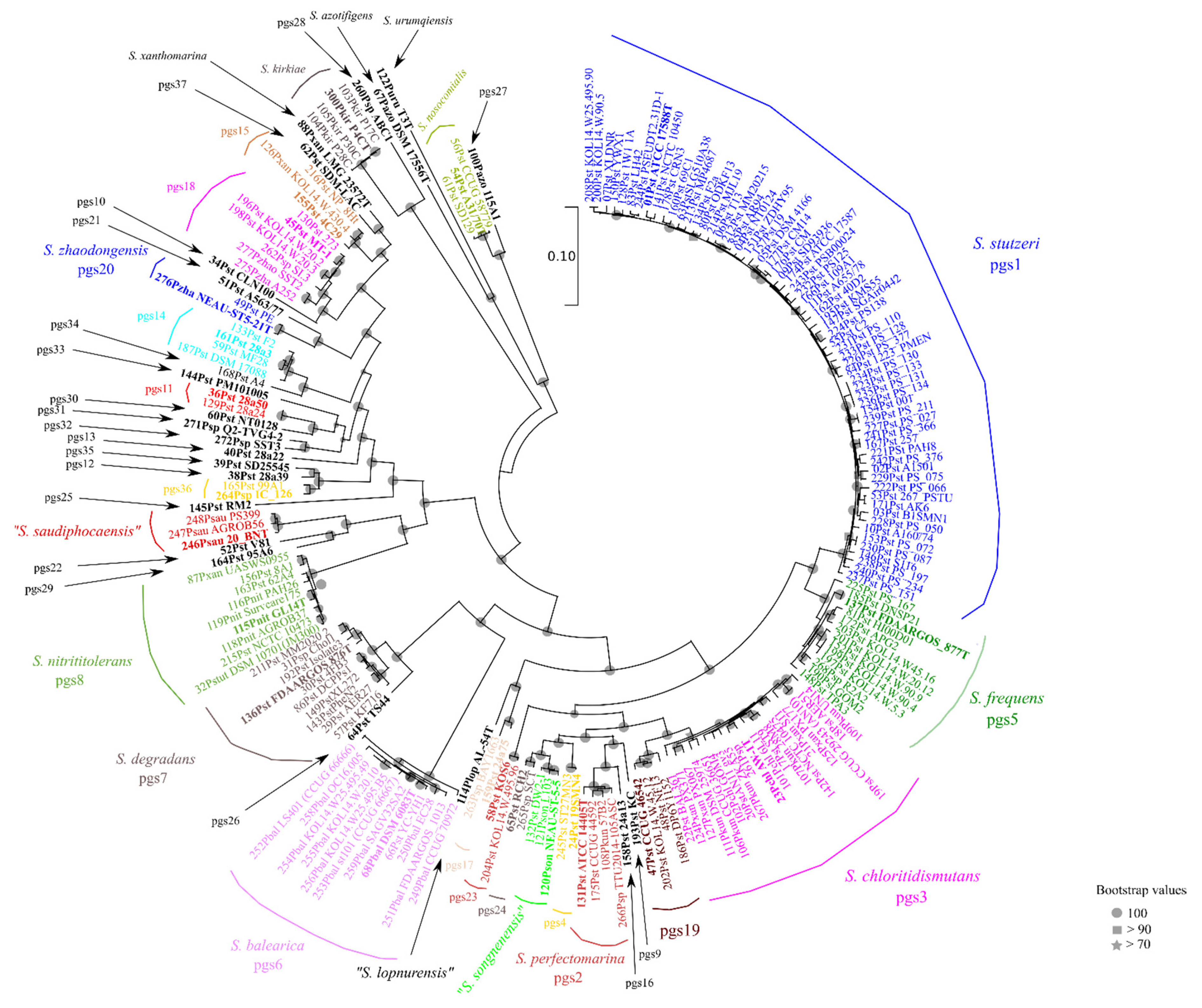
3.2. Core-Genome Phylogeny
3.3. Digital Species Delineation (ANIb, GGDC, and Phyletic Pattern)
3.4. Phenotypic Characterization and Species Descriptions
4. Discussion
5. Taxonomic Conclusions
5.1. Emended Description of S. perfectomarina (pgs2)
5.2. Emended Description of S. chloritidismutans (pgs3) and Synonymy with P. kunmingensis
5.3. Description of S. frequens sp. nov. (pgs5)
5.4. Description of S. degradans sp. nov. (pgs7)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Lalucat, J.; Gomila, M.; Mulet, M.; Zaruma, A.; García-Valdés, E. Past, present and future of the boundaries of the Pseudomonas genus: Proposal of Stutzerimonas gen. nov. Syst. Appl. Microbiol. 2022, 45, 126289. [Google Scholar] [CrossRef] [PubMed]
- Ursing, J.B.; Rosselló-Mora, R.A.; Garcia-Valdés, E.; Lalucat, L. Taxonomic note: A pragmatic approach to the nomenclature of phenotypically similar genomic groups. Int. J. Syst. Evol. 1995, 45, 604. [Google Scholar] [CrossRef] [Green Version]
- Lalucat, J.; Bennasar, A.; Bosch, R.; García-Valdés, E.; Palleroni, N.J. Biology of Pseudomonas stutzeri. Microbiol. Mol. Biol. Rev. 2006, 70, 510–547. [Google Scholar] [CrossRef] [Green Version]
- Oren, A.; Garrity, G.M. Valid publication of new names and new combinations effectively published outside the IJSEM. Int. J. Syst. Evol. Microbiol. 2022, 72, 005260. [Google Scholar] [CrossRef]
- Parks, D.H.; Chuvochina, M.; Waite, D.W.; Rinke, C.; Skarshewski, A.; Chaumeil, P.A.; Hugenholtz, P. A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat. Biotech. 2018, 36, 996–1004. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R.; Glöckner, F.O.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Sardà Carbasse, J.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acid Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef]
- Staley, J.T. Transitioning toward a universal species concept for the classification of all organisms. In The Species Problem; Pavlinov, I., Ed.; IntechOpen: London, UK, 2013. [Google Scholar]
- Avram, O.; Rapoport, D.; Portugez, S.; Pupko, T. M1CR0B1AL1Z3R—A user-friendly web server for the analysis of large-scale microbial genomics data. Nucleic Acids Res. 2019, 47, W88–W92. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-R, L.M.; Konstantinidis, K.T. The enveomics collection: A toolbox for specialized analyses of microbial genomes and metagenomes. Peer J. Prepr. 2016, 4, e1900v1. [Google Scholar]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Cowan, S.T. Cowan and Steel’s Manual for the Identification of Medical Bacteria, 2nd ed.; Cambridge University Press: Cambridge, UK, 1974. [Google Scholar]
- Lalucat, J. Analysis of Refractile (R) Bodies. In Methods in Microbiology; Mayer, F., Ed.; Academic Press: Cambridge, MA, USA, 1988; Volume 20, pp. 79–90. [Google Scholar]
- Rosselló-Mora, R.A.; Lalucat, J.; Dott, W.; Kämpfer, P. Biochemical and chemotaxonomic characterization of Pseudomonas stutzeri genomovars. J. Appl. Bacteriol. 1994, 76, 226–233. [Google Scholar] [CrossRef]
- Lalucat, J.; Mulet, M.; Gomila, M.; García-Valdés, E. Genomics in bacterial taxonomy: Impact on the genus Pseudomonas. Genes 2020, 11, 139. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Yang, Z.; Wang, W.; Li, W.; Zhang, G.; Yan, H. Comparative genomics of Pseudomonas stutzeri complex: Taxonomic assignments and genetic diversity. Front. Microbiol. 2022, 12, 755874. [Google Scholar] [CrossRef]
- Girard, L.; Lood, C.; Höfte, M.; Vandamme, P.; Rokni-Zadeh, H.; van Noort, V.; Lavigne, R.; De Mot, R. The ever-expanding Pseudomonas genus: Description of 43 new species and partition of the Pseudomonas putida group. Microorganisms 2021, 9, 1766. [Google Scholar] [CrossRef]
- Rosselló, R.; Garcia-Valdés, E.; Lalucat, J.; Ursing, J. Genotypic and phenotypic diversity of Pseudomonas stutzeri. Syst. Appl. Microbiol. 1991, 14, 150–157. [Google Scholar] [CrossRef]
- Scotta, C.; Gomila, M.; Mulet, M.; Lalucat, J.; García-Valdés, E. Whole-cell MALDI-TOF mass spectrometry and multilocus sequence analysis in the discrimination of Pseudomonas stutzeri populations: Three novel genomovars. Microb. Ecol. 2013, 66, 522–532. [Google Scholar] [CrossRef]
- Mulet, M.; Gomila, M.; Ramírez, A.; Lalucat, J.; Garcia-Valdes, E. Pseudomonas nosocomialis sp. nov., isolated from clinical specimens. Int. J. Syst. Evol. Microbiol. 2019, 69, 3392–3398, Erratum in Int. J. Syst. Evol. Microbiol.2022, 72, 005300. [Google Scholar] [CrossRef]
- Wolterink, A.F.W.M.; Jonker, A.B.; Kengen, S.W.M.; Stams, A.J.M. Pseudomonas chloritidismutans sp. nov., a non-denitrifying, chlorate-reducing bacterium. Int. J. Syst. Evol. Microbiol. 2002, 52 Pt 6, 2183–2190. [Google Scholar]
- Cladera, A.M.; García-Valdés, E.; Lalucat, J. Genotype versus phenotype in the circumscription of bacterial species: The case of Pseudomonas stutzeri and Pseudomonas chloritidismutans. Arch. Microbiol. 2006, 184, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Mulet, M.; Bennasar, A.; Lalucat, L.; García-Valdés, E. An rpoD-based PCR procedure for the identification of Pseudomonas species and for their detection in environmental samples. Mol. Cell Probes 2009, 23, 140–147. [Google Scholar] [CrossRef]
- Anwar, N.; Rozahon, M.; Zayadan, B.; Mamtimin, H.; Abdurahman, M.; Kurban, M.; Abdurusul, M.; Mamtimin, T.; Abdukerim, M.; Rahman, E. Pseudomonas tarimensis sp. nov., an endophytic bacteria isolated from Populus euphratica. Int. J. Syst. Evol. Microbiol. 2017, 67, 4372–4378. [Google Scholar] [CrossRef]
- Peng, J.S.; Liu, Y.; Yan, L.; Hou, T.T.; Liu, H.C.; Zhou, Y.G.; Liu, Z.P. Pseudomonas nitrititolerans sp. nov., a nitrite-tolerant denitrifying bacterium isolated from a nitrification/denitrification bioreactor. Int. J. Syst. Evol. Microbiol. 2019, 69, 2471–2476. [Google Scholar] [CrossRef]
- Zou, Y.; He, S.; Sun, Y.; Zhang, X.; Liu, Y.; Cheng, Q. Pseudomonas urumqiensis sp. nov., isolated from rhizosphere soil of Alhagi sparsifolia. Int. J. Syst. Evol. Microbiol. 2019, 69, 1760–1766. [Google Scholar] [CrossRef]
- Baumann, P.; Bowditch, R.D.; Baumann, L.; Beaman, B. Taxonomy of marine Pseudomonas species: P. stanieri sp. nov.; P. perfectomarina sp. nov., nom. rev.; P. nautica; and P. doudoroffii. Int. J. Syst. Bacteriol. 1983, 33, 857–865. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Pan, Y.; Wang, K.; Zhang, X.; Zhang, C.; Zhang, S.; Fu, X.; Jiang, J. Pseudomonas zhaodongensis sp. nov., isolated from saline and alkaline soils. Int. J. Syst. Evol. Microbiol. 2015, 65, 1022–1030. [Google Scholar] [CrossRef]
- Xie, F.; Ma, H.; Quan, S.; Liu, D.; Chen, G.; Chao, Y.; Qian, S. Pseudomonas kunmingensis sp. nov., an exopolysaccharide-producing bacterium isolated from a phosphate mine. Int. J. Syst. Evol. Microbiol. 2014, 64 Pt 2, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Zobell, C.E.; Upham, H.C. A list of marine bacteria including description of sixty species. Bull. Scripps Inst. Oceanogr. 1944, 5, 239–292. [Google Scholar]
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiol. Mol. Biol. Rev. 1997, 61, 533–616. [Google Scholar] [PubMed]
- Döhler, K.; Huss, V.A.R.; Zumft, W.G. Transfer of Pseudomonas perfectomarina Baumann, Bowditch, Baumann, and Beaman 1983 to Pseudomonas stutzeri (Lehmann and Neumann 1896) Sijderius 1946. Int. J. Syst. Evol. Microbiol. 1987, 37, 1–3. [Google Scholar] [CrossRef]
- Rosselló-Mora, R.A.; García-Valdés, E.; Lalucat, J. Taxonomic relationship between Pseudomonas perfectomarina ZoBell and Pseudomonas stutzeri. Int. J. Syst. Evol. Microbiol. 1993, 43, 852–854. [Google Scholar] [CrossRef] [Green Version]
- Peña, A.; Busquets, A.; Gomila, M.; Bosch, R.; Nogales, B.; García-Valdés, E.; Lalucat, J.; Bennasar, A. Draft genome of Pseudomonas stutzeri strain ZoBell (CCUG 16156), a marine isolate and model organism for denitrification studies. J. Bacteriol. 2012, 194, 1277–1278. [Google Scholar] [CrossRef] [Green Version]
- Stanier, R.Y.; Palleroni, N.J.; Doudoroff, M. The aerobic Pseudomonas: A taxonomic study. J. Gen. Microbiol. 1966, 43, 159. [Google Scholar] [CrossRef] [Green Version]
- Mehboob, F.; Junca, H.; Schraa, G.; Stams, A.J. Growth of Pseudomonas chloritidismutans AW-1(T) on n-alkanes with chlorate as electron acceptor. Appl. Microbiol. Biotechnol. 2009, 83, 739–747. [Google Scholar] [CrossRef] [Green Version]
- Youngblut, M.D.; Wang, O.; Barnum, T.P.; Coates, J.D. (Per)chlorate in Biology on Earth and Beyond. Annu. Rev. Microbiol. 2016, 70, 435–457. [Google Scholar] [CrossRef]
- Rosselló-Mora, R.A. Caracterización Taxonómica de Cepas de Pseudomonas stutzeri Degradadoras y no Degradadoras de Naftaleno. Ph.D. Thesis, Universitat de les Illes Balears, Ciutat de Mallorca, Spain, 1992. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Species Names Proposed in the Current Taxonomy (1) | Strain | Assignation to P. stutzeri Genomovar | GTDB Taxonomy | Proposed Species Assignation to Stutzerimonas |
|---|---|---|---|---|
| S. azotifigens | 6H33bT; ATCC BAA-1049T; DSM 17556T; JCM 12708T | - | Pseudomonas_N azotifigens | S. azotifigens |
| S. balearica | CCUG 44487T; CIP 105297T; DSM 6083; SP1402T | gv6 ref | Pseudomonas_A balearica | S. balearica |
| S. chloritidismutans | ATCC BAA-443T; AW-1T; DSM 13592T; NBRC 102227T | gv3 | Pseudomonas_A chloritidismutans | S. chloritidismutans |
| S. kirkiae | FRB 229T; LMG 31089T; NCPPB 4674T; P4CT | - | Pseudomonas_Q kirkiae | S. kirkiae |
| S. kunmingensis | CGMCC 1.12273T; DSM 25974T; HL22-2 T | gv3 | Pseudomonas_A kunmingensis | S. chloritidismutans |
| P. nitrititolerans | CGMCC 1.13874T; GL14T; NBRC 113853T | gv8 | Pseudomonas_A nitrititolerans | S. nitrititolerans |
| S. nosocomialis | A31/70T; CCUG 73638T; CECT 9881T | - | Pseudomonas_R nosocomialis | S. nosocomialis |
| P. perfectomarina | ZoBellT; ATCC 14405T; LMG 14935T | gv2 | Pseudomonas_A stutzeri_AC | S. perfectomarina |
| S. stutzeri | ATCC 17588T; CCUG 11256T; CFBP 2443T; CIP 103022T; DSM 5190T; IFO 14165T; JCM 5965T; LMG 11199T; NBRC 14165T | gv1 ref | Pseudomonas_A stutzeri | S. stutzeri |
| P. tarimensis | CCTCC AB 2013065T; KCTC 42447T; MA-69T | - | not found | S. tarimensis |
| S. urumqiensis | ACCC 60124T; JCM 32830T; T3T | - | Pseudomonas_N sp003640395 | S. urumqiensis |
| S. xanthomarina | CCUG 46543T; DSM 18231T; JCM 12468T; KMM 1447T; NRIC 617T | - | Pseudomonas_A xanthomarina | S. xanthomarina |
| S. zhaodongensis | ACCC 06362T; DSM 27559T; NEAU-ST5-21T | gv20 | Pseudomonas_A zhaondongensis | S. zhaodongensis |
| “P. lopnurensis” | AL-54T; CCTCC AB 2013066T; JCM 19136T; NRRL B-59987T | - | not found | “S. lopnurensis” |
| “P. saudiphocaensis” | 20_BNT | - | Pseudomonas_A saudiphocaensis | “S. saudiphocaensis” |
| “P. songnenensis” | ACCC 06361T; DSM 27560T; NEAU-ST5-5T | - | Pseudomonas_A songnenensis | “S. songnenensis” |
| P. stutzeri | CCUG 44592; ATCC 17591 | gv2 ref | Pseudomonas_A stutzeri_AC | S. perfectomarina |
| P. stutzeri | DSM 50227; CCUG 711, NRRL B-927, NCTC 10475, LMG 1228, CIP 59.54, DSM 50227, ATCC 11607, ICPB 2445, NCIB 9721 | gv3 ref | Pseudomonas_A chloritidismutans | S. chloritidismutans |
| P. stutzeri | 19SMN4; CCUG 44593; DSM 6084; LMG 18521 | gv4 ref | Pseudomonas_A stutzeri_A | Stutzerimonas pgs4 |
| P. stutzeri | DNSP21; CCUG 44521; DSM 6082; LMG 18520 | gv5 ref | Pseudomonas_A sp003205815 | S. frequens |
| P. stutzeri | DSM 50238 | gv7 ref | Pseudomonas_A sp000765155 | S. degradans |
| P. stutzeri | JM300 | gv8 ref | Pseudomonas_A stutzeri_AG | S. nitrititolerans |
| P. stutzeri | KC; DSM 7136 | gv9 ref | Pseudomonas_A stutzeri_AA | Stutzerimonas pgs9 |
| P. stutzeri | CLN100 | gv10 ref | not found | Stutzerimonas pgs10 |
| P. stutzeri | 28a50 | gv11 ref | Pseudomonas_A stutzeri_D | Stutzerimonas pgs11 |
| P. stutzeri | 28a39 | gv12 ref | Pseudomonas_A stutzeri_AI | Stutzerimonas pgs12 |
| P. stutzeri | 28a22 | gv13 ref | not found | Stutzerimonas pgs13 |
| P. stutzeri | 28a3 | gv14 ref | Pseudomonas_A stutzeri_AI | Stutzerimonas pgs14 |
| P. stutzeri | 4C29 | gv15 ref | Pseudomonas_A stutzeri_AB | Stutzerimonas pgs15 |
| P. stutzeri | 24a13 | gv16 ref | Pseudomonas_A stutzeri_AH | Stutzerimonas pgs16 |
| P. stutzeri | 24a75 | gv17 ref | Pseudomonas_A stutzeri_AF | Stutzerimonas pgs17 |
| P. stutzeri | MT-1; CCUG 50545 | gv18 ref | Pseudomonas_A sp004010935 | Stutzerimonas pgs18 |
| P. stutzeri | CCUG 46542 | gv19 ref | not found | Stutzerimonas pgs19 |
| P. stutzeri | PE | gv20 ref | not found | S. zhaodongensis |
| P. stutzeri | A563/77 | gv21 ref | not found | Stutzerimonas pgs21 |
| P. stutzeri | V81 | gv22 ref | not found | Stutzerimonas pgs22 |
| P. stutzeri | KOS6 | Pseudomonas_A stutzeri_B | Stutzerimonas pgs23 | |
| P. stutzeri | RCH2 | Pseudomonas_A stutzeri_AE | Stutzerimonas pgs24 | |
| P. stutzeri | RM2 | not found | Stutzerimonas pgs25 | |
| P. stutzeri | TS44 | Pseudomonas_A stutzeri_C | Stutzerimonas pgs26 | |
| P. azotifigens | 115A1 | Pseudomonas_R azotifigens | Stutzerimonas pgs27 | |
| Pseudomonas sp. | ABC1 | Pseudomonas_Q sp013395055 | Stutzerimonas pgs28 | |
| P. stutzeri | 95A6 | Pseudomonas_A stutzeri_AK | Stutzerimonas pgs29 | |
| P. stutzeri | NT0128 | Pseudomonas_A stutzeri_E | Stutzerimonas pgs30 | |
| Q2-TVG-42 | not found | Stutzerimonas pgs31 | ||
| Pseudomonas sp. | SST3 | Pseudomonas_A sp003325755 | Stutzerimonas pgs32 | |
| P. stutzeri | PM101005 | Pseudomonas_A stutzeri_R | Stutzerimonas pgs33 | |
| P. stutzeri | A4 | not found | Stutzerimonas pgs34 | |
| P. stutzeri | SD25545 | gv12 | not found | Stutzerimonas pgs35 (former gv12) |
| P. stutzeri | 99A1 | Pseudomonas_A sp004331835 | Stutzerimonas pgs36 | |
| P. stutzeri | SDM-LAC | Pseudomonas_A stutzeri_H | Stutzerimonas pgs37 | |
| P. stutzeri | AN10 | gv3 | Pseudomonas_A stutzeri_U | S. chloritidismutans |
| P. stutzeri | 28a24 | gv11 | Pseudomonas_A stutzeri_D | Stutzerimonas pgs11 |
| P. stutzeri | NF13 | gv19 | Pseudomonas_A stutzeri_G | Stutzerimonas pgs19 |
| Species Name | Stutzerimonas chloritidismutans |
|---|---|
| Species status | Validly published under the ICNP |
| Species etymology | chlo.ri.ti.dis.mu.tans. Gr. fem. n. chlôris, a female yellow bird; N.L. pres. part. dismutans, splitting; N.L. part. adj. chloritidismutans, chlorite-splitting |
| Designation of the type strain | AW-1T |
| Strain collection numbers | ATCC BAA-443; DSM 13592; NBRC 102227 |
| 16S rRNA gene accession number | AY017341 (1501 nt) |
| Alternative housekeeping genes | gyrB gene [AJ880092.2] and rpoD gene [AJ880091.1] |
| Genome accession number | AOFQ00000000.1 |
| Genome status | Draft |
| Genome size | 5.0 Mb |
| GC mol% | 62.5 |
| Country of origin | Netherlands |
| Region of origin | Wageningen |
| Other strains | CCUG 29243; ST-9; DSM 25974T |
| Date of isolation | 2002 |
| Source of isolation | Biomass of an anaerobic bioreactor treating chlorate- and bromate-polluted wastewater |
| Sampling date | 2002 |
| Heterotypic synonyms | Pseudomonas stutzeri gv3 (DSM 50227), Pseudomonas kunmingensis DSM 25974T |
| Source of isolation of non-type strains | Clinical specimens, before 1952 (DSM 50227); phosphate rock powder from a phosphate mining field; China; Yunnan Province, suburb of Kunming; isolated: 20.10.2009 (DSM 25974T); soil; sea water; contaminated marine sediment |
| Growth medium, incubation conditions used for standard cultivation | Luria-Bertani medium (LB) at 30 °C |
| Gram stain | Negative |
| Cell shape | Rods |
| Cell size (length or diameter) | Cells are 0.5–2 µm in size |
| Motility | Motile by a single polar flagellum |
| Colony morphology | Colonies growing in anaerobic agar are circular and pale orange. Colonies on nutrient broth plates are wrinkled, coherent, and have a pale brown or sometimes reddish, brown color. Smooth colonies may be produced. |
| Growth temperature range | 10–37 °C |
| Temperature optimum | 30 °C |
| Growth pH range | 7–9 |
| Growth NaCl range | 0–4 |
| Metabolism | Facultatively aerobic, strictly respiratory, able to grow anaerobically with nitrate or chlorate (some strains) |
| BIOLOG GEN III positive tests for the type strain | Substrates oxidized: alfa-D-glucose, Tween 40, dextrin, D-mannitol, methyl-pyruvate, gamma-amino-butyric acid, D-maltose, D-fructose, D-arabitol, L-alanine, alfa-hydroxy butyric acid, D-trehalose, D-gluconic acid, L-lactic acid, beta-Hydroxy-D-L-butyric acid, glycerol, L-aspartic acid, citric acid, alfa-keto-butyric acid, L-glutamic acid, alfa-keto glutaric acid, mucic acid, D-malic acid, propionic acid, L-pyroglutamic acid, L-malic acid, acetic acid, D-saccharic acid, formic acid, Growth: 1% NaCl, 1% sodium lactate, troleandomycin, lincomycin, vancomycin nalidixic acid, aztreonam, 4% NaCl, fusidic acid, rifamycin SV, guanidine HCl, tetrazolium violet, lithium chloride, sodium butyrate, D-serine, minocycline, niaproof 4, tetrazolium blue, potassium tellurite. |
| BIOLOG GEN III negative tests for the type strain | Not able to oxidize: D-raffinose, D-sorbitol, gelatine, pectin, p-hydroxy, phenylacetic acid, alfa D-lactose, D-mannose, glycyl-L-proline, D-galacturonic acid, D-melibiose, D, lactic acid methyl ester, beta-methyl-D-glucoside, D-galactose, myo-inositol, L-arginine, D-cellobiose, D-salicin, 3-methyl glucose, D-glucuronic acid, D-gentiobiose, N-acetyl-D-glucosamine, D-glucose-6-PO4, glucuronamide, acetoacetic acid, sucrose, N-acetyl, beta-D-mannosamine, L-fucose, D-fructose-6-PO4, L-histidine, turanose, N-acetyl-D-galactosamine, L-rhamnose, D-aspartic acid, quinic acid, stachyose, N-acetyl-neuraminic acid, inosine, D-serine, L-Serine, pH 5 |
| BIOLOG GEN III differential tests for strains DSM 50227 and DSM 25974T | Oxidized substrates: D-galactonic acid lactone, D-glucuronic acid, mucic acid, L-pyroglutamic acid, D-saccharic acid. |
| API 20NE positive tests for the type strain | Reduction of nitrate to nitrite and nitrite to N2, glucose, mannitol, maltose, gluconate, caprate, malate, citrate. |
| API 20NE negative tests for the type strain | Indole production, glucose fermentation. Presence of arginine dihydrolase, urease, hydrolysis of aesculin, hydrolysis of gelatin, beta-galactosidase- Assimilation of: Arabinose, Mannose, N-acetyl-D-glucosamine, Adipate, Phenylacetate. |
| API 20NE differential tests for strains DSM 50227 and DSM 25974T | Assimilation of Mannitol |
| Energy metabolism of strains in the species | Chemoorganotroph, strictly oxidative, anaerobic with nitrate or chlorate. Thiosulfate oxidation in 2 strains. |
| Oxidase | Positive |
| Catalase | Positive |
| Pigment production on King A and King B media | Negative |
| Major fatty acids (mean values of 10 strains) [17] | 0.10% decanoate (10:0); 2.79% 3-hydroxydecanoate (3OH 10:0); 0.00% 10-methyldodecanoate (a-13:0); 7.69% dodecanoate (12:0); 2.64% 3-hydroxydodecanoate (3OH 12:0); 0.98% tetradecanoate (14:0); 0.00% pentadecanoate (15:0); 28.77% cis 9 hexadecanoate (16:1); 21.45% hexadecanoate (16:0); 0.61% 15 methylhexadecenoate (17:0); 1.26% cis 9,10 methylenehexadecanoate (17:0c); 29.58% cis 9,11 octadecenoate (18:1); 1.46% octadecanoate (18:0); 1.45% cis 9,10 methyleneoctadecanoate (19:0c). |
| Biosafety level | 1 |
| Habitat | Soil, wastewater sludge, marine sediment, rhizosphere, clinical specimens |
| Biotic relationship | Free-living |
| Known pathogenicity | None |
| Species Name | Stutzerimonas perfectomarina |
|---|---|
| Species status | nom. rev. |
| Species etymology | per.fec.to.ma.rí.na. L. masc. perf. part. perfectus, complete, perfect; L. fem. adj. marina, of the sea, marine; N.L. fem. adj. perfectomarina, completely marine |
| Previous validly published names (Synonyms) | Bacterium stutzeri (Lehmann and Neumann 1896), Pseudomonas perfectomarinus (ZoBell and Upham 1944), Pseudomonas perfectomarina (Baumann et al., 1983) trasferred to Pseudomonas stutzeri (Döhler et al., 1983), member of genomovar 2 of P. stutzeri (Rosselló-Móra et al., 1993) |
| Designation of the type strain | ZoBell 632 |
| Type strain collection numbers | Baumann 218; ATCC 14405; CCUG 16156; CECT 4899; JCM 20778; LMG 2243 |
| 16S rRNA gene accession number | U26420 (1456 nt), U65012 (rRNA operon) |
| Alternative housekeeping genes | gyrB gene [AJ617564], and rpoD gene [AGSL01000000] |
| Genome accession number | AGSL01000000 |
| Genome status | Draft |
| Genome size (pb) | 4.7 Mb |
| GC mol% | 61.4 |
| Country of origin | USA |
| Region of origin | California, Pacific Ocean |
| Date of isolation | 1944 |
| Source of isolation | Marine water |
| Sampling date | 1944 |
| Other strains | Stanier 224 (=ATCC 17591, =CIP 107689, =CCUG 44592), Stanier 220a (=ATCC 17587), Stanier 225a (=ATCC 17592), Stanier 228a (=ATCC 17595) |
| Source of isolation of non-type strains | Clinical specimens, indoor dust and Bos taurus feces |
| Growth medium, incubation conditions used for standard cultivation | Luria-Bertani medium (LB) at 30 °C |
| Gram stain | Negative |
| Cell shape | Straight or slightly curved rod |
| Cell size (length or diameter) | 1.1 to 1.9 µm long and 0.5 to 0.7 µm wide |
| Motility | Motile by a single polar flagellum |
| Colony morphology | Colonies are mucous, beige-colored, from round to oval, with irregular margins. 2–3 mm in diameter after growth for 48 h, 2–5 mm after 72 h at 30 °C on LB plates. Smooth and rough colonial morphologies may be produced. |
| Growth temperature range | 4–40 °C |
| Temperature optimum | 30 °C |
| Growth pH range | 7–10 |
| Growth NaCl range | 0–8% |
| Metabolism | Facultatively aerobic, strictly respiratory, able to grow anaerobically with nitrate |
| BIOLOG GEN III positive tests for the type strain | Growth: 1–4–8% NaCl, 1% sodium lactate, troleandomycin, lincomycin, vancomycin, nalidixic acid, aztreonam, pH 6, fusidic acid, rifamycin SV, guanidine HCl, tetrazolium violet, lithium chloride, sodium butyrate, D-serine, minocycline, niaproof 4, tetrazolium blue, potassium tellurite. Substrates oxidized: alfa-D-glucose, tween 40, dextrin, D-galacturonic acid, methyl pyruvate, D-maltose, D-fructose, D-arabitol, L-alanine, D-galactonic acid lactone, alfa-hydroxy butyric acid, L-lactic acid, beta-hydroxy-D, L-butyric acid, glycerol, L-aspartic acid, D-glucuronic acid, citric acid, alfa-keto-butyric acid, L-glutamic acid, alfa-keto glutaric acid, mucic acid, D-malic acid, propionic acid, L-pyroglutamic acid, L-malic acid, acetic acid, D-saccharic acid, and formic acid. |
| BIOLOG GEN III negative tests for the type strain | No growth at pH 5. Not able to oxidize D-raffinose, D-sorbitol, gelatin, pectin, p-hydroxy-phenylacetic acid, alfa-D-lactose, D-mannose, D-mannitol, glycyl-L-proline, gamma-amino-butyric acid, D-melibiose, D-lactic acid methyl ester, D-trehalose, beta-methyl-D-glucoside, D-galactose, myo-Inositol, L-arginine, D-gluconic acid, D-cellobiose, D-salicin, 3-methyl glucose, gentiobiose, N-acetyl-D-galactosamine, D-fucose, D-glucose-6-PO4, acetoacetic acid, sucrose, N-acetyl-beta-D-mannosamine, L-fucose, D-fructose-6-PO4, D-fructose-6-PO4, turanose, N-acetyl-D-galactosamine, L-rhamnose, D-aspartic acid, quinic acid, stachyose, N-acetyl-neuraminic acid, inosine, N-serine, and bromo-succinic acid. |
| BIOLOG GEN III differential tests for strain ATCC 17591 | Oxidation of D-mannitol, D-galactonic acid, D-trehalose, D-gluconic acid, mucic acid, D-saccaric acid, and formic acid. |
| API 20NE positive tests for the type strain | Reduction of nitrate to nitrite and nitrite to N2, the assimilation of glucose, caprate, malate, and citrate. |
| API 20NE negative tests for the type strain | Indole production, glucose fermentation, arginine dihydrolase, urease, aesculin hydrolysis, hydrolysis of gelatin, and beta-galactosidase. Assimilation of arabinose, mannose, mannitol, N-acetyl-D-glucosamine, maltose, gluconate, adipate, phenyl-acetate. |
| API 20NE differential tests for strain ATCC 17591 | Mannitol, maltose, gluconate. |
| Energy metabolism | Chemoorganotrophic, denitrifier with nitrate, strictly oxidative |
| Oxidase | Positive |
| Catalase | Positive |
| Pigment production on King A and King B media | Negative |
| Major fatty acids (mean values of 5 strains) [17] | 0.06% decanoate (10:0); 2.92% 3-hydroxydecanoate (3OH 10:0); 0.00 10-methyldodecanoate (a-13:0); 9.33% dodecanoate (12:0); 2.77% 3-hydroxydodecanoate (3OH 12:0); 0.72% tetradecanoate (14:0); 0.03% pentadecanoate (15:0); 25.55% cis 9 hexadecanoate (16:1); 20.93% hexadecanoate (16:0); 0.4% 15 methylhexadecenoate (17:0); 0.26% cis 9,10 methylenehexadecanoate (17:0c); 34.64% cis 9,11 octadecenoate (18:1); 1.80% octadecanoate (18:0); 0.46% cis 9,10 methyleneoctadecanoate (19:0c). |
| Biosafety level | 1 |
| Habitat | Marine, human urine |
| Biotic relationship | Free-living |
| Known pathogenicity | Not known pathogenicity but present in clinical specimens |
| Species Name | Stutzerimonas frequens |
|---|---|
| Species status | sp. nov. |
| Species etymology | fre’quens. L. adj. frequens, frequently isolated from different habitats |
| Designation of the type strain | DNSP21 |
| Type strain collection numbers | DSM 6082; LMG 1852; CCUG 44594 |
| 16S rRNA gene accession number | U26414.1 (1456 nt) |
| Alternative housekeeping genes | gyrB gene [AJ620493.1] and rpoD gene [AJ631335.1] |
| Genome accession number | POUJ01000000 and NZ CP065720 |
| Genome status | Draft |
| Genome size (pb) | 4.4 Mb |
| GC mol% | 58.7 |
| Country of origin | Spain |
| Region of origin | Mallorca (Santa Ponça, wastewater treatment plant) |
| Date of isolation | 1988 |
| Source of isolation | Wastewater |
| Sampling date | 1988 |
| Geographic location | Wastewater treatment plant, Santa Ponça, |
| Latitude | 39°31′18.8″ N |
| Longitude | 2°30′45.4″ E |
| Other strains | St103, St104, HI00D01, R2A2, KOL14.W.45.16, KOL14.W.90.4, KOL14.W.5.3, KOL14.W.20.12, KOL14.W.90.9, THAF7b, TPA3, ChG 5-2 |
| Source of isolation of non-type strains | Pacific Ocean, near Hawaii (HI00D01); groundwater South-west Taiwan; hydrothermal vent Greece (KOL14.W.45.16, KOL14.W.90.4, KOL14.W.5.3, KOL14.W.20.12, KOL14.W.90.9); microplastics (THAF7b); soil (TPA3); Phragmites australis (St103), and Spartina patens (St104) rhizosphere; Black Sea 120m depth (ChG 5-2). |
| Growth medium, incubation conditions used for standard cultivation | Luria-Bertani medium (LB) at 30 °C |
| Gram stain | Negative |
| Cell shape | Rod |
| Cell size (length or diameter) | 2 − 5 × 0.8 − 1 µm |
| Motility | Motile, one polar inserted flagellum |
| Colony morphology | Colonies are round, dry, and wrinkled, beige-colored, with irregular margins. Its size was 2–3 mm to 3–5 mm in diameter after growth for 48 h at 30 °C on LB plates. Colonies strongly attached to the agar. |
| Growth temperature range | 4–37 °C |
| Temperature optimum | 30 °C |
| Growth pH range | 6–10 |
| Growth NaCl concentration | 0–8 |
| Metabolism | Facultatively aerobic, strictly respiratory, able to grow anaerobically with nitrate |
| BIOLOG GENIII positive tests for the type strain | The strains have the ability to oxidize: alfa-D-glucose, Tween 40, dextrin, D-mannitol, methyl-pyruvate, gamma-amino-butyric acid, D-maltose, D-fructose, L-alanine, alfa-hydroxy-butyric acid, D-gluconic acid, L-lactic acid, beta-hydroxy-D, L-butyric acid, glycerol, citric acid, alfa-keto-butyric acid, L-glutamic acid, alfa-keto glutaric acid, D-malic acid, propionic acid, L-malic and acid, acetic acid, bromo-succinic acid, formic acid. Able to grow at 1% NaCl, 4% NaCl 8% NaCl, 1% sodium lactate, troleandomycin, lincomycin, vancomycin, nalidixic acid, aztreonam, pH 6, fusidic acid, rifamycin SV, guanidine HCl, tetrazolium violet, lithium chloride, sodium butyrate, D-serine, minocycline, niaproof 4, potassium tellurite, sodium bromate. |
| BIOLOG GENIII negative tests for the type strain | The strains were unable to oxidize: D-raffinose, D-sorbitol, gelatin, pectin, p-hydroxy-phenylacetic acid, alfa-D-lactose, D-mannose, glycyl-L-proline, D-galacturonic acid, D-melibiose, D-arabitol, D-galactonic acid lactone, D-lactic acid methyl ester, D-trehalose, beta-methyl-D-glucoside, D-galactose, myo-inositol, L-arginine, D-cellobiose, D-salicin, 3-methyl glucose, L-aspartic acid, D-glucuronic acid, gentiobiose, N-acetyl-D-glucosamine, D-fucose, D-glucose-6-PO4, acetoacetic acid, sucrose, N-acetyl-beta-D-mannosamine, L-fucose, D-fructose-6-PO4, L-histidine, mucic acid, turanose, N-acetyl-D-galactosamine, L-rhamnose, D-aspartic acid, L-pyroglutamic acid, quinic acid, stachyose, N-acetyl-neuraminic acid, inosine, D-serine, L-serine, D-saccharic acid Unable to grow at pH 5. |
| API 20NE positive tests for the type strain | Reduction of nitrate to nitrite and nitrite to N2, assimilation of glucose, maltose, gluconate, caprate, malate, and citrate. |
| API 20NE negative tests for the type strain | Indole production, glucose fermentation, arginine dihydrolase, urease, hydrolysis of aesculin, hydrolysis of gelatine, beta-galactosidase. No assimilation of arabinose, mannose, mannitol, N-acetyl-D-glucosamine, adipate, and phenylacetate. |
| Energy metabolism | Chemoorganotroph, denitrifier, strictly oxidative |
| Oxidase | Positive |
| Catalase | Positive |
| Pigment production on King A and King B media | Negative |
| Major fatty acids of the type strain [17] | 0.00% decanoate (10:0); 3.42% 3-hydroxydecanoate (3OH 10:0); 0.00% 10-methyldodecanoate (a-13:0); 8.46% dodecanoate (12:0); 2.88% 3-hydroxydodecanoate (3OH 12:0); 0.05% tetradecanoate (14:0); 0.00% pentadecanoate (15:0); 24.78% cis 9 hexadecanoate (16:1); 21.57% hexadecanoate (16:0); 0.40% 15 methylhexadecenoate (17:0); 0.28% cis 9,10 methylenehexadecanoate (17:0c); 35.79% cis 9,11 octadecenoate (18:1); 1.04% octadecanoate (18:0); 0.32% cis 9,10 methyleneoctadecanoate (19:0c). |
| Biosafety level | 1 |
| Habitat | Marine water, ground water, hydrothermal vent, and soil |
| Biotic relationship | Free-living |
| Known pathogenicity | None |
| Species Name | Stutzerimonas degradans |
|---|---|
| Species status | sp. nov. |
| Species etymology | de.gra’dans. L. part adj. degradans, returning to the original order, referring to the ability of the type strain to degrade contaminants |
| Designation of the type strain | DSM 50238 |
| Type strain collection numbers | ATCC 17832, ICPB 2737-419, CCUG 44596, LMG 14935, CIP 107692, Stanier 419 |
| 16S rRNA gene accession number | U26416.1 (1456 bp) |
| Alternative housekeeping genes | gyrB gene [AJ631262.1] and rpoD gene [AJ631339.1] |
| Genome accession number | CP065721 |
| Genome status | Complete |
| Genome size (pb) | 4.0 Mb |
| GC mol% | 64.5 |
| Country of origin | USA |
| Region of origin | Berkeley, California |
| Other strains | AER2.7, 4FB3, 2FB7A, Chol1, DCP-Ps1, KF716 |
| Other strains in this pgs | HMP271, KF716, DCP-Ps1, PheN2, JL972, XL272, MM2020_2, D-134-1 |
| Date of isolation | Before 1966 |
| Source of isolation | Isolated from the soil by L-(+)-tartrate enrichment |
| Geographic location | California |
| Region of origin | Berkeley |
| Source of isolation of non-type strains | Aircraft-oil contaminated soil (AER2.7); estuarine sediment (4FB3, 2FB7A); soil (Chol1, DCP-Ps1, KF716); coal cutting; lab enrichment culture in anaerobic medium (HMP271), biphenyl contaminated soil (KF716), dichlorophenol enrichment culture inoculated with agricultural soil (DCP-Ps1), anaerobic environments (PheN2), aquatic sediment (JL972), cucumber rhizosphere (XL272), freshwater sediment (MM2020_2), soil (D-134-1) |
| Growth medium, incubation conditions used for standard cultivation | Luria-Bertani medium (LB) at 30 °C |
| Gram stain | Negative |
| Cell shape | Rods |
| Cell size (length or diameter) | 3.5 − 4.8 × 0.3 − 0.7 µm |
| Motility | Motile, one polar inserted flagellum |
| Colony morphology | Freshly isolated strains: rough, wrinkled, dry and coherent, light brown in color. Colonies are round, beige color. Its size is 2–3 mm in diameter after growth for 48 h at 30 °C on LB plates. Colonies with regular margins and translucent may be produced. |
| Growth temperature range | 10–42 °C |
| Temperature optimum | 30 °C |
| Growth pH range | 6–10 |
| Growth NaCl range | 0–8% |
| Metabolism | Facultatively aerobic, strictly respiratory, able to grow anaerobically with nitrate |
| BIOLOG GENIII positive tests for the type strain | Oxidized substrates: alfa-D-glucose, Tween 40, dextrin, methyl pyruvate, gamma-amino-butyric acid, D-maltose, L-alanine, alfa-hydroxy butyric acid, L-lactic acid, beta-hydroxy-D, L-butyric acid, glycerol, L-aspartic acid, citric acid, alfa-keto butyric acid. L-glutamic acid, alfa-keto glutaric acid, D-malic acid, propionic acid, L-malic acid, acetic acid, L-serin, bromo-succinic acid. Growth with 1–4–8% NaCl, pH 6, 1% sodium lactate, troleandomycin, lincomycin, vancomycin, nalidixic acid, aztreonam, fusidic acid, rifamycin SV, guanidine HCl, tetrazolium violet, lithium chloride, sodium butyrate, D-serine, minocycline, niaproof 4, tetrazolium blue and potassium tellurite. |
| BIOLOG GENIII negative tests for the type strain | Not oxidized substrates: D-raffinose, D-sorbitol, gelatin, pectin, p-hydroxy-phenylacetic acid, alfa-D-lactose, D-mannose, D-mannitol, glycyl-L-proline, D-galacturonic acid, D-melibiose, D-fructose, D-arabitol, D-galactonic acid lactone, D-lactic acid methyl ester, D-trehalose, beta-methyl-D-glucoside, D-galactose, myo-inositol, L-arginine, D-gluconic acid, D-cellobiose, D-salicin, 3-methylglucose, D-glucuronic acid, gentiobiose, N-acetyl-D-glucosamine, D-glucose-6-PO4, glucuronamide, acetoacetic acid, N-acetyl-beta-D-mannosamine, L-fucose, D-fructose-6-PO4, L-histidine, mucic acid, turanose, N-acetyl-D-galactosamine, L-rhamnose, D-aspartic acid, L-pyroglutamic acid, quinic acid, stachyose, N-acetyl-neuraminic acid, inosine, D-serine, and D-saccharic acid, formic acid. Strains were unable to grow at pH 5, nor with sodium bromate. |
| API 20NE positive tests for the type strain | Reduction of nitrate to nitrite and nitrite to N2, the assimilation of glucose, maltose, caprate, malate, and citrate. |
| API 20NE negative tests for the type strain | Indole production, glucose fermentation, arginine dihydrolase, urease, hydrolysis of aesculin, Hydrolysis of gelatin, beta-galactosidase, arabinose, mannose, mannitol, and N-acetyl-D-glucosamine, gluconate, adipate, phenylacetate |
| Energy metabolism | Chemoorganotroph, denitrifier, strictly oxidative |
| Oxidase | Positive |
| Catalase | Positive |
| Pigment production on King A and King B media | Negative |
| Major fatty acids [17] | 0.00% decanoate (10:0); 2.76% 3-hydroxydecanoate (3OH 10:0); 0.00 10-methyldodecanoate (a-13:0); 10.80% dodecanoate (12:0); 2.51% 3-hydroxydodecanoate (3OH 12:0); 1.04% tetradecanoate (14:0); 0.00% pentadecanoate (15:0); 29.60% cis 9 hexadecanoate (16:1); 22.47% hexadecanoate (16:0); 0.76% 15 methylhexadecenoate (17:0); 1.64% cis 9,10 methylenehexadecanoate (17:0c); 26.15% cis 9,11 octadecenoate (18:1); 1.37% octadecanoate (18:0); 0.89% cis 9,10 methyleneoctadecanoate (19:0c). |
| Biosafety level | 1 |
| Habitat | Soil, estuarine sediment |
| Biotic relationship | Free-living |
| Known pathogenicity | None |
| New Name Combination and Etymology | Basonym | Description | Type Strain and Culture Collection Numbers |
|---|---|---|---|
| Stutzerimonas nitrititolerans nitritum, nitrite; L. pres. part. tolerans, tolerating; N.L. part. adj. nitrititolerans, tolerating nitrite | Pseudomonas nitrititolerans | The description of this taxon is as given by Peng et al., 2019 [28] | CGMCC 1.13874T; GL14T; NBRC 113853T |
| Stutzerimonas nosocomialis no.so.co.mi.a’lis. N.L. neut. n. nosocomium, a hospital, infirmary; L. masc./fem. adj. suff. -alis, suffix used with the sense of pertaining to; N.L. masc./fem. adj. nosocomialis, pertaining to a hospital | Pseudomonas nosocomialis | The description of this taxon is as given by Mulet et al., 2019; and 2022 [23] | A31/70T; CCUG 73638T; CECT 9881T |
| Stutzerimonas tarimensis ta.rim.en’sis. N.L. masc./fem. adj. tarimensis, pertaining to Tarim basin in Xinjiang Uyghur autonomous region of China, where the type strain was isolated | Pseudomonas tarimensis | The description of this taxon is as given by Anwar et al., 2017 [27] | CCTCC AB 2013065T; KCTC 42447T; MA-69T |
| Stutzerimonas urumqiensis u.rum.qi.en’sis. N.L. masc./fem. adj. urumqiensis, pertaining to Urumqi, in Xinjiang Uyghur Autonomous Region of China, 45°5′6″ N, 94°58′36″ E, where the sample was collected | Pseudomonas urumqiensis | The description of this taxon is as given by Zou et al., 2019 [29] | ACCC 60124T; JCM 32830T; T3T |
| Stuzerimonas perfectomarina per.fec.to.ma.ri.na. L. masc. perf. Part. perfectus, complete, perfect; L. fem. adj. marina, of the sea, marine; N.L. fem. adj. perfectomarina, completely marine | Pseudomonas perfectomarina | The description of this taxon is as given by Bauman et al., 1983 [30] and by the present study | ZoBell 632T; Baumann 218T; ATCC 14405T; CCUG 16156T; CECT 4899T; JCM 20778T; LMG 2243T |
| Stutzerimonas zhaodongensis zhao.dong.en’sis. N.L. masc./fem. adj. zhaodongensis, pertaining to Zhaodong City, China, where the type strain was isolated | Pseudomonas zhaodongensis | The description of this taxon is as given by Zhang et al., 2015 [31] | ACCC 06362T; DSM 27559T; NEAU-ST5-21T |
| Heterotypic synonym: | |||
| Stutzerimonas chloritidismutans | Pseudomonas kunmingensis (heterotypic synonym) | The description of this taxon is as given by Xie et al., 2014 [32] and the present study | ATCC BAA-443T; DSM 13592T; NBRC 102227T; AW-1T |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gomila, M.; Mulet, M.; García-Valdés, E.; Lalucat, J. Genome-Based Taxonomy of the Genus Stutzerimonas and Proposal of S. frequens sp. nov. and S. degradans sp. nov. and Emended Descriptions of S. perfectomarina and S. chloritidismutans. Microorganisms 2022, 10, 1363. https://doi.org/10.3390/microorganisms10071363
Gomila M, Mulet M, García-Valdés E, Lalucat J. Genome-Based Taxonomy of the Genus Stutzerimonas and Proposal of S. frequens sp. nov. and S. degradans sp. nov. and Emended Descriptions of S. perfectomarina and S. chloritidismutans. Microorganisms. 2022; 10(7):1363. https://doi.org/10.3390/microorganisms10071363
Chicago/Turabian StyleGomila, Margarita, Magdalena Mulet, Elena García-Valdés, and Jorge Lalucat. 2022. "Genome-Based Taxonomy of the Genus Stutzerimonas and Proposal of S. frequens sp. nov. and S. degradans sp. nov. and Emended Descriptions of S. perfectomarina and S. chloritidismutans" Microorganisms 10, no. 7: 1363. https://doi.org/10.3390/microorganisms10071363
APA StyleGomila, M., Mulet, M., García-Valdés, E., & Lalucat, J. (2022). Genome-Based Taxonomy of the Genus Stutzerimonas and Proposal of S. frequens sp. nov. and S. degradans sp. nov. and Emended Descriptions of S. perfectomarina and S. chloritidismutans. Microorganisms, 10(7), 1363. https://doi.org/10.3390/microorganisms10071363