
Pandemic Clones of CTX-M-15 Producing Klebsiella pneumoniae ST15, ST147, and ST307 in Companion Parrots

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Bacterial Strains
2.2. Antimicrobial Susceptibility Tests
2.3. Virulence and Genotypic Profile of Klebsiella pneumoniae Strains
2.4. Plasmid Analysis
2.5. Whole Genome Sequencing and Analysis of ESBL-Producing K. pneumoniae Strains
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Wyres, K.L.; Holt, K.E. Klebsiella pneumoniae Population Genomics and Antimicrobial-Resistant Clones. Trends Microbiol. 2016, 24, 944–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyres, K.L.; Lam, M.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Antimicrobial Resistance. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 7 December 2020).
- Lee, M.; Choi, T.J. Antimicrobial resistance caused by KPC-2 encoded by promiscuous plasmids of the Klebsiella pneumoniae ST307 strain. Ann. Lab. Med. 2021, 41, 86–94. [Google Scholar] [CrossRef]
- Han, Y.; Huang, L.; Liu, C.; Huang, X.; Zheng, R.; Lu, Y.; Wenying, X.; Fang, N.; Yaning, M.; Liu, G. Characterization of carbapenem-resistant Klebsiella pneumoniae st15 clone coproducing kpc-2, ctx-m-15 and shv-28 spread in an intensive care unit of a tertiary hospital. Infect. Drug Resist. 2021, 14, 767–773. [Google Scholar] [CrossRef]
- Rad, Z.R.; Rad, Z.R.; Goudarzi, H.; Goudarzi, M.; Alizade, H.; Mazraeh, F.N.; Sharahi, J.Y.; Ardebili, A.; Hashemi, A. Detection of NDM-1 producing Klebsiella pneumoniae ST15 and ST147 in Iran during 2019–2020. Acta Microbiol. Imm. Hung. 2021, 68, 177–182. [Google Scholar] [CrossRef]
- Hayakawa Ito de Sousa, A.T.; dos Santos Costa, M.T.; Makino, H.; Cândido, S.L.; de Godoy Menezes, I.; Lincopan, N.; Nakazato, L.; Dutra, V. Multidrug-resistant mcr-1 gene-positive Klebsiella pneumoniae ST307 causing urinary tract infection in a cat. Braz. J. Microbiol. 2021, 52, 1043–1046. [Google Scholar] [CrossRef]
- Hidasi, H.W.; Hidasi Neto, J.; Moraes, M.D.C.; Linhares, G.F.C.; Jayme, V.D.S.; Andrade, M.A. Enterobacterial Detection and Escherichia coli Antimicrobial Resistance in Parrots Seized from the Illegal Wildlife Trade. J. Zoo Wildl. Med. 2013, 44, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Davies, Y.M.; Cunha, M.P.V.; Oliveira, M.G.X.; Oliveira, M.C.V.; Philadelpho, N.; Romeiro, D.C.; Milanelo, L.; Guimaraes, M.B.; Ferreira, A.J.P.; Moreno, A.M.; et al. Virulence and antimicrobial resistance of Klebsiella pneumoniae isolated from passerines and psittacine birds. Avian Pathol. 2016, 45, 194–201. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.M.; Tang, H.L.; Chiou, C.S.; Tung, K.C.; Lu, M.C.; Lai, Y.C. Colonization dynamics of Klebsiella pneumoniae in the pet animals and human owners in a single household. Vet. Microbiol. 2021, 256, 109050. [Google Scholar] [CrossRef]
- Silva, G.G.D.C.; Campana, E.H.; Vasconcelos, P.C.; Silva, N.M.V.D.; Santos Filho, L.; Leite, E.L.; Oliveira, C.J.B.D. Occurrence of KPC-Producing Escherichia coli in Psittaciformes Rescued from Trafficking in Paraíba, Brazil. Int. J. Environ. Res. Public Health 2021, 18, 95. [Google Scholar] [CrossRef]
- Kuhnert, P.; Bisgaard, M.; Korczak, B.M.; Scwendener, S.; Christensen, H.; Frey, J. Identification of animal Pasteurellaceae by MALDI-TOF mass spectrometry. Methods Mol. Biol. 2015, 1247, 235–243. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- McLauchlin, J.; Narayanan, G.L.; Mithani, V.; O’Neill, G. The detection of enterotoxins and toxic shock síndrome toxin genes in Staphylococcus aureus by polymerase chain reaction. J. Food Prot. 2000, 63, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Van Belkum, A.; Tassios, P.T.; Dijkshoorn, L.; Haeggman, S.; Cookson, B.; Fry, N.K.; Fussing, V.; Green, J.; Feil, E.; Gerner-Smidt, P.; et al. Guidelines for the validation and application of typing methods for use in bacterial epidemiology. Clin. Microbiol. Infect. 2007, 13, 1–46. [Google Scholar] [CrossRef]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Villa, L.; García-Fernández, A.; Fortini, D.; Carattoli, A. Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J. Antimicrob. Chemother. 2010, 65, 2518–2529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Wick, R.R.; Heinz, E.; Holt, K.E.; Wyres, K.L. Kaptive Web: User-friendly capsule and lipopolysaccharide serotype prediction for Klebsiella genomes. J. Clin. Microbiol. 2018, 56, e00197-18. [Google Scholar] [CrossRef] [Green Version]
- Gato, E.; Vázquez Ucha, J.C.; Rumbo-Feal, S.; Alvarez Fraga, L.; Vallejo, J.; Martínez-Guitián, M.; Beceiro, A.; Ramos-Vivas, J.; Campoy, P.; Perez-Vazquez, M.; et al. Kpi, a chaperone-usher pili system associated with the worldwide-disseminated high-risk clone Klebsiella pneumoniae ST-15. Proc. Natl. Acad. Sci. USA 2020, 117, 17249–17259. [Google Scholar] [CrossRef]
- Cunha, M.P.; Davies, Y.M.; Cerdeira, L.; Dropa, M.; Lincopan, N.; Knöbl, T. Complete DNA sequence of an IncM1 plasmid bearing the novel qnrE1 plasmid-mediated quinolone resistance variant and bla CTX-M-8 from Klebsiella pneumoniae sequence type 147. Antimicrob. Agents Chemother. 2017, 61, e00592-17. [Google Scholar] [CrossRef] [Green Version]
- Holt, K.E.; Wertheim, H.; Zadoks, R.N.; Baker, S.; Whitehouse, C.A.; Dance, D. Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. USA 2015, 112, E3574–E3581. [Google Scholar] [CrossRef] [Green Version]
- Padmini, N.; Ajilda, A.A.K.; Sivakumar, N.; Selvakumar, G. Extended spectrum β-lactamase producing Escherichia coli and Klebsiella pneumoniae: Critical tools for antibiotic resistance pattern. J. Basic Microbiol. 2017, 57, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Diekema, D.J.; Hsueh, P.R.; Mendes, R.E.; Pfaller, M.A.; Rolston, K.V.; Sader, H.S.; Jones, R.N. The Microbiology of Bloodstream Infection: 20-Year Trends from the SENTRY Antimicrobial Surveillance Program. Antimicrob. Agents Chemother. 2019, 63, e00355-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, C.; Menezes, J.; Belas, A.; Aboim, C.; Cavaco-Silva, P.; Trigueiro, G.; Telo Gama, L.; Pomba, C. Klebsiella pneumoniae causing urinary tract infections in companion animals and humans: Population structure, antimicrobial resistance and virulence genes. J. Antimicrob. Chemother. 2019, 74, 594–602. [Google Scholar] [CrossRef]
- Valencia-Bacca, J.; Silva, M.M.; Cerdeira, L.; Esposito, F.; Cardoso, B.; Muñoz, M.E.; Jiménez-Villegas, T.; Cardenas-Arias, A.; Pessoa, D.A.N.; Lincopan, N. Detection and Whole-Genome Analysis of a High-Risk Clone of Klebsiella pneumoniae ST340/CG258 Producing CTX-M-15 in a Companion Animal. Microb. Drug Resist. 2020, 26, 611–615. [Google Scholar] [CrossRef]
- Sellera, F.P.; Fuga, B.; Fontana, H.; Esposito, F.; Cardoso, B.; Konno, S.; Berl, C.; Cappellanes, M.H.; Cortez, M.; Ikeda, M.; et al. Detection of IncN-pST15 one-health plasmid harbouring blaKPC-2 in a hypermucoviscous Klebsiella pneumoniae CG258 isolated from an infected dog, Brazil. Transbound Emerg. Dis. 2021, 68, 3083–3088. [Google Scholar] [CrossRef]
- Garcia-Fierro, R.; Drapeau, A.; Dazas, M.; Saras, E.; Rodrigues, C.; Brisse, S.; Madec, J.Y.; Haenni, M. Comparative phylogenomics of ESBL-, AmpC- and carbapenemase-producing Klebsiella pneumoniae originating from companion animals and humans. J. Antimicrob. Chemother. 2022, 77, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, M.S.; O’connor, C.; Miller, V.L. Yersiniabactin is a virulence factor for Klebsiella pneumoniae during pulmonary infection. Infect. Immun. 2007, 75, 1463–1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the Offense with a Strong Defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Wyres, K.L.; Nguyen, T.N.; Lam, M.; Judd, L.M.; van Vinh Chau, N.; Dance, D.A.; Margaret, L.P.; Karkey, A.; Ling, C.L.; Miliya, T.; et al. Genomic surveillance for hypervirulence and multi-drug resistance in invasive Klebsiella pneumoniae from South and Southeast Asia. Genome Med. 2020, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Villa, L.; Feudi, C.; Fortini, D.; Brisse, S.; Passet, V.; Bonura, C.; Endimiani, A.; Mammina, C.; Ocampo, A.M.; Jimenez, J.N.; et al. Diversity, virulence, and antimicrobial resistance of the KPC-producing Klebsiella pneumoniae ST307 clone. Microb. Genom. 2017, 3, e000110. [Google Scholar] [CrossRef]
- Haller, S.; Kramer, R.; Becker, K.; Bohnert, J.A.; Eckmanns, T.; Hans, J.B.; Hecht, J.; Heidecken, C.; Hübner, N.; Kramer, A.; et al. Extensively drug-resistant Klebsiella pneumoniae ST307 outbreak, north-eastern Germany, June to October 2019. Eurosurveillance 2019, 24, 1900734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonstra, M.B.; Spijkerman, D.C.; Voor, A.F.; van der Laan, R.J.; Bode, L.G.; van Vianen, W.; Klaassen, C.H.W.; Vos, M.C.; Severin, J.A. An outbreak of ST307 extended-spectrum beta-lactamase (ESBL)–producing Klebsiella pneumoniae in a rehabilitation center: An unusual source and route of transmission. Infect. Control Hosp. Epidemiol. 2020, 41, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Dropa, M.; Lincopan, N.; Balsalobre, L.C.; Oliveira, D.E.; Moura, R.A.; Fernandes, M.R.; Da Silva, Q.M.; Matté, G.R.; Sato, M.I.Z.; Matté, M.H. Genetic background of novel sequence types of CTX-M-8- and CTX-M-15-producing Escherichia coli and Klebsiella pneumoniae from public wastewater treatment plants in São Paulo, Brazil. Environ. Sci. Pollut. Res. 2016, 23, 4953–4958. [Google Scholar] [CrossRef] [PubMed]
- Sartori, L.; Sellera, F.P.; Moura, Q.; Cardoso, B.; Cerdeira, L.; Lincopan, N. Multidrug-resistant CTX-M-15-positive Klebsiella pneumoniae ST307 causing urinary tract infection in a dog in Brazil. J. Glob. Antimicrob. Resist. 2019, 19, 96–97. [Google Scholar] [CrossRef]
- Ewers, C.; Stamm, I.; Pfeifer, Y.; Wieler, L.H.; Kopp, P.A.; Schinning, K.; Prenger-Berninghoff, E.; Scheufen, S.; Stolle, I.; Gunther, S.; et al. Clonal spread of highly successful ST15-CTX-M-15 Klebsiella pneumoniae in companion animals and horses. J. Antimicrob. Chemother. 2014, 69, 2676–2680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ovejero, C.M.; Escudero, J.A.; Thomas-Lopez, D.; Hoefer, A.; Moyano, G.; Montero, N.; Espada, C.M.; Gonzalez-Zorn, B. Highly tigecycline-resistant Klebsiella pneumoniae sequence type 11 (ST11) and ST147 isolates from companion animals. Antimicrob. Agents Chemother. 2017, 61, e02640-16. [Google Scholar] [CrossRef] [Green Version]
- Calbo, E.; Garau, J. The changing epidemiology of hospital outbreaks due to ESBL-producing Klebsiella pneumoniae: The CTX-M-15 type consolidation. Future Microbiol. 2005, 10, 1063–1075. [Google Scholar] [CrossRef]
- Carattoli, A. Plasmids and the spread of resistance. Int. J. Med. Microbiol. 2013, 303, 298–304. [Google Scholar] [CrossRef]
- Naseer, U.; Sundsfjord, A. The CTX-M conundrum: Dissemination of plasmids and Escherichia coli clones. Microbial. Drug Resist. 2011, 17, 83–97. [Google Scholar] [CrossRef]
- Zhao, W.H.; Hu, Z.Q. Epidemiology and genetics of CTX-M extended-spectrum β-lactamases in Gram-negative bacteria. Crit. Rev. Microbiol. 2013, 39, 79–101. [Google Scholar] [CrossRef]
- Cerdeira, L.; Silva, K.C.; Fernandes, M.R.; Lenne, S.; Souza, T.A.; Garcia, D.O.; Moreno, A.M.; Lincopan, N. Draft genome sequence of a CTX-M-15-producing Klebsiella pneumoniae sequence type 340 (clonal complex 258) isolated from food-producing animal. J. Glob. Antimicrob. Resist. 2016, 7, 67–68. [Google Scholar] [CrossRef] [PubMed]
- Norizuki, C.; Wachino, J.I.; Suzuki, M.; Kawamura, K.; Nagano, N.; Kimura, K.; Arakawa, Y. Specific bla CTX-M-8/IncI1 plasmid transfer among genetically diverse Escherichia coli isolates between humans and chickens. Antimicrob. Agents Chemother. 2017, 61, e00663-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albornoz, E.; Tijet, N.; De Belder, D.; Gomez, S.; Martino, F.; Corso, A.; Melano, R.G.; Petroni, A. qnrE1, a Member of a New Family of Plasmid-located Quinolone Resistance Genes Originated from the Chromosome of Enterobacter spp. Antimicrob. Agents Chemother. 2017, 61, e02555-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
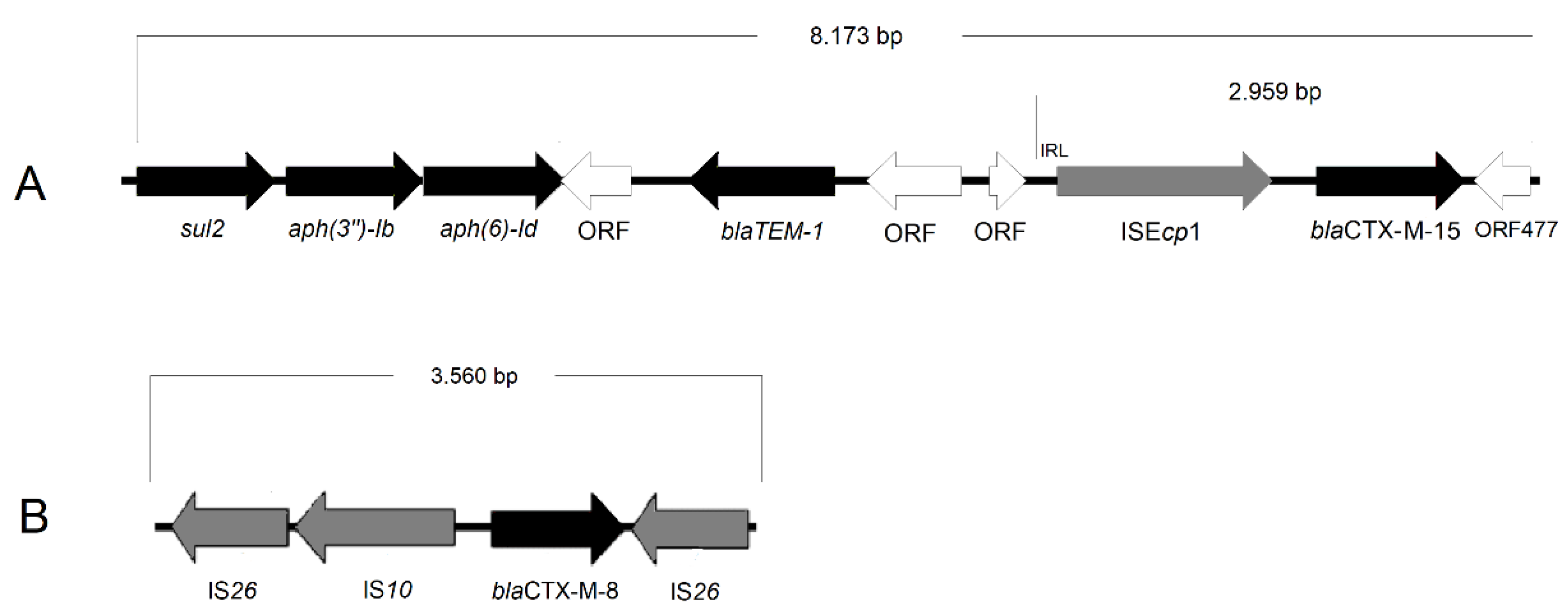{kind=link}
| Isolates Information | Strains | ||
|---|---|---|---|
| Kp41 | Kp58 | Kp137 | |
| Host | Amazona aestiva (nasal secretion) | Psittacara leucophtalmus (oropharynx) | Amazona aestiva (sepsis) |
| Codified Regions (CDS) | 5548 | 5351 | 5436 |
| RNAs | 93 | 96 | 86 |
| GC Content | 57.1% | 57.2% | 57.1% |
| Sequence type | ST147 | ST15 | ST307 |
| Plasmids-group (size) | FII(K) (~220 kb) L/M (~70 kb) ColRNAI (3 kb) | FII(K)-FIA-FIB (~250 kb) | FIB (K) (~145 kb) NT (48 kb) ColRNAI (4 kb) |
| Capsule | K64 | K60 | K173 |
| Resistance genes | |||
| Aminoglycosides | aac(3)-VIa, aph(3″)-Ib, aac(3)-IIa, aadA1, aph(6)-Id | aph(3″)-Ib, aac(6′)Ib-cr, aph(6)-Id, aadA2 | aac(3)-IIa, aph(3″)-Ib, aac(6′)Ib-cr, aph(6)-Id |
| Beta-lactams | blaSHV-11, blaCTX-M-8, blaCTX-M-15, blaTEM-1 A, blaOXA-1, blaOXA-9 | blaSHV-28, blaCTX-M-15, blaTEM-1 B, blaOXA-1 | blaSHV-28, blaCTX-M-15, blaTEM-1 B, blaOXA-1 |
| Quinolones | qnrE, oqxAB, gyrA (mutation) | aac(6′)Ib-cr, oqxAB, gyrA (mutation) | aac(6′)Ib-cr, oqxAB, qnrB66, gyrA (mutation) |
| Fosfomycin | fosA | fosA | fosA |
| Tetracyclines | tet(A) | - | |
| Sulfonamides | sul1, sul2 | sul1, sul2 | sul2 |
| Trimethoprim | dfrA14 | dfrA14, dfrA12 | dfrA14 |
| Phenicols | catB4 | catA1, catB4 | catB4 |
| Strain | Plasmid Sizes | MIC (mg/L) | |||
|---|---|---|---|---|---|
| Cefotaxime | Ceftazidime | Cefepime | Ceftiofur | ||
| Kp41 | 220 kb 70 kb 3 kb | >128 | 128 | >128 | >128 |
| Tc-41a | 220 kb | >128 | 32 | >128 | >128 |
| Kp58 | 250 kb | >128 | 128 | >128 | >128 |
| Tc-58a | 250 kb | >128 | 64 | >128 | >128 |
| Kp137 | 145 Kb 48 kb 4 kb | >128 | 128 | >128 | >128 |
| Tc-137a | 145 Kb | >128 | 64 | >128 | >128 |
| EC-C600 | - | 0.125 | 0.5 | 0.125 | 0.125 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davies, Y.M.; Cunha, M.P.V.; Dropa, M.; Lincopan, N.; Gomes, V.T.M.; Moreno, L.Z.; Sato, M.I.Z.; Moreno, A.M.; Knöbl, T. Pandemic Clones of CTX-M-15 Producing Klebsiella pneumoniae ST15, ST147, and ST307 in Companion Parrots. Microorganisms 2022, 10, 1412. https://doi.org/10.3390/microorganisms10071412
Davies YM, Cunha MPV, Dropa M, Lincopan N, Gomes VTM, Moreno LZ, Sato MIZ, Moreno AM, Knöbl T. Pandemic Clones of CTX-M-15 Producing Klebsiella pneumoniae ST15, ST147, and ST307 in Companion Parrots. Microorganisms. 2022; 10(7):1412. https://doi.org/10.3390/microorganisms10071412
Chicago/Turabian StyleDavies, Yamê Miniero, Marcos Paulo Vieira Cunha, Milena Dropa, Nilton Lincopan, Vasco Túlio Moura Gomes, Luisa Zanoli Moreno, Maria Inês Zanoli Sato, Andrea Micke Moreno, and Terezinha Knöbl. 2022. "Pandemic Clones of CTX-M-15 Producing Klebsiella pneumoniae ST15, ST147, and ST307 in Companion Parrots" Microorganisms 10, no. 7: 1412. https://doi.org/10.3390/microorganisms10071412
APA StyleDavies, Y. M., Cunha, M. P. V., Dropa, M., Lincopan, N., Gomes, V. T. M., Moreno, L. Z., Sato, M. I. Z., Moreno, A. M., & Knöbl, T. (2022). Pandemic Clones of CTX-M-15 Producing Klebsiella pneumoniae ST15, ST147, and ST307 in Companion Parrots. Microorganisms, 10(7), 1412. https://doi.org/10.3390/microorganisms10071412