
Exploiting the Potential of Bioreactors for Creating Spatial Organization in the Soil Microbiome: A Strategy for Increasing Sustainable Agricultural Practices

 , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
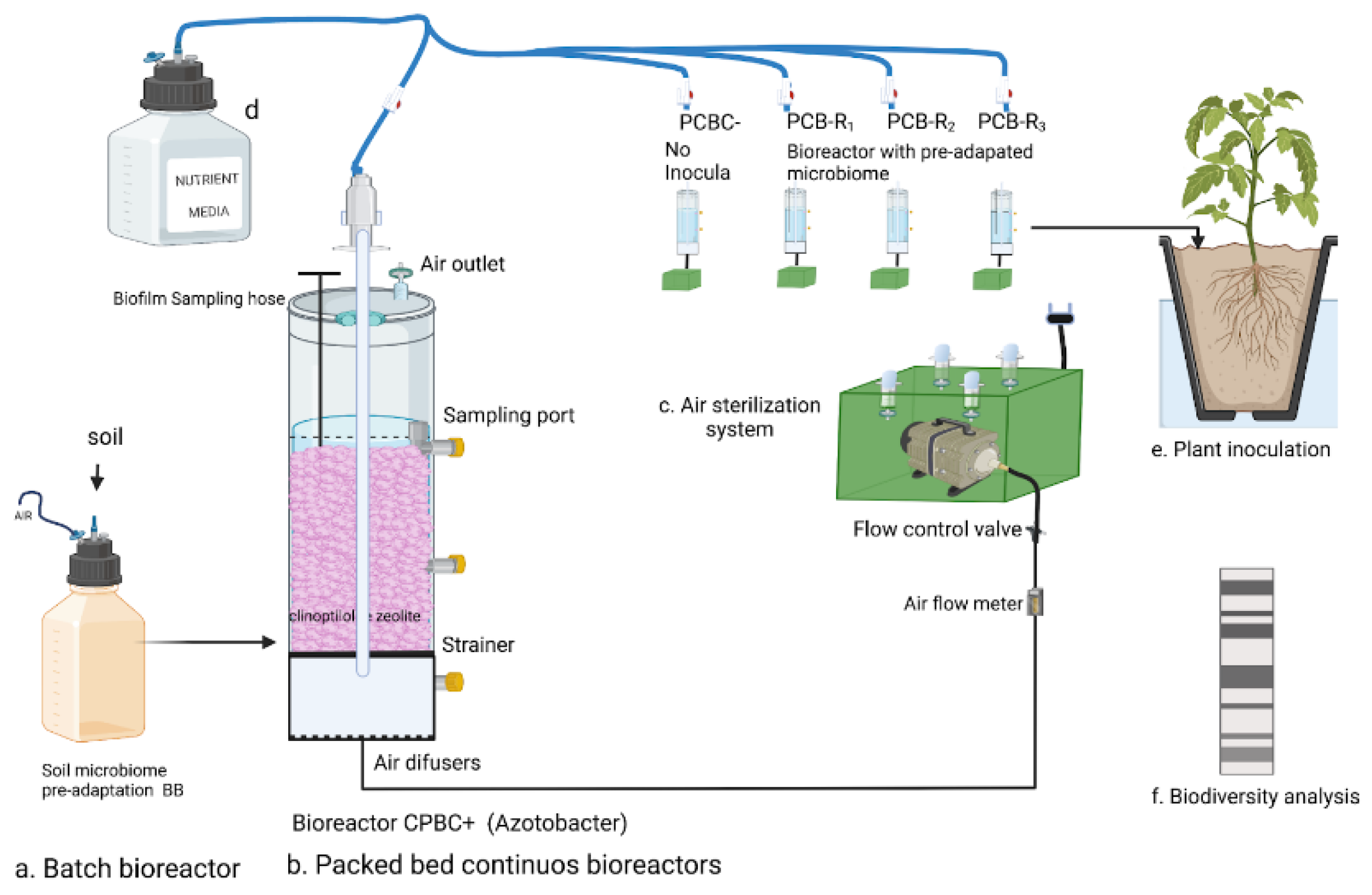
2.1. Microbial Inoculum Pre-Adaptation in Batch Reactors
2.2. Packed-Bed Bioreactor’s Configuration
2.3. Reactor Sampling and Processing
2.4. Fertilizer’s Solution Preparation
2.5. Soil and Seed Selection for Planting
2.6. Tomato Plant Growth Assessment
2.7. DNA Extraction, Sequencing, and Metabarcoding Analysis
2.8. Statistical Analysis
3. Results
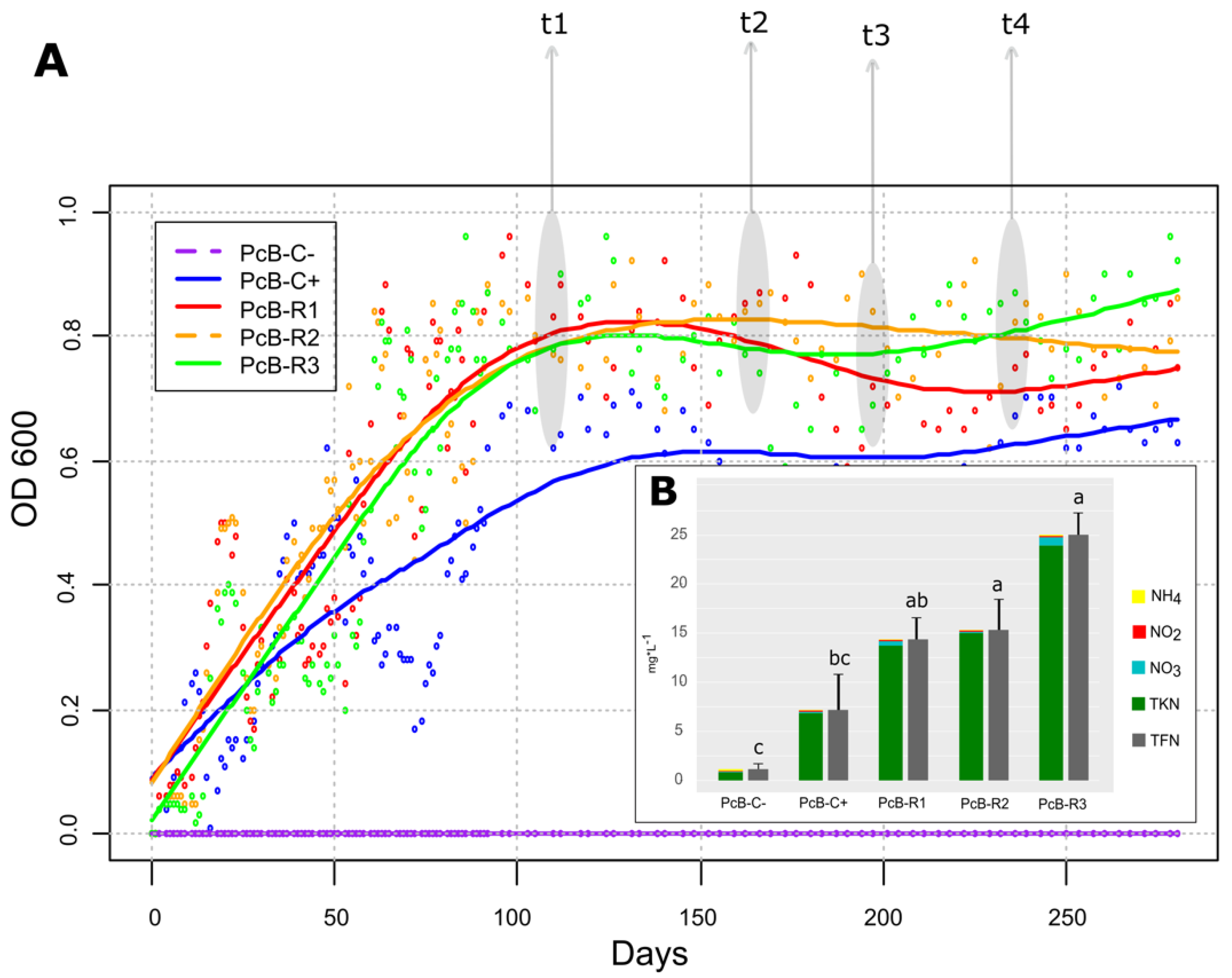
3.1. Reactor’s Startup and Performance
3.2. Bioinoculant Characteristics
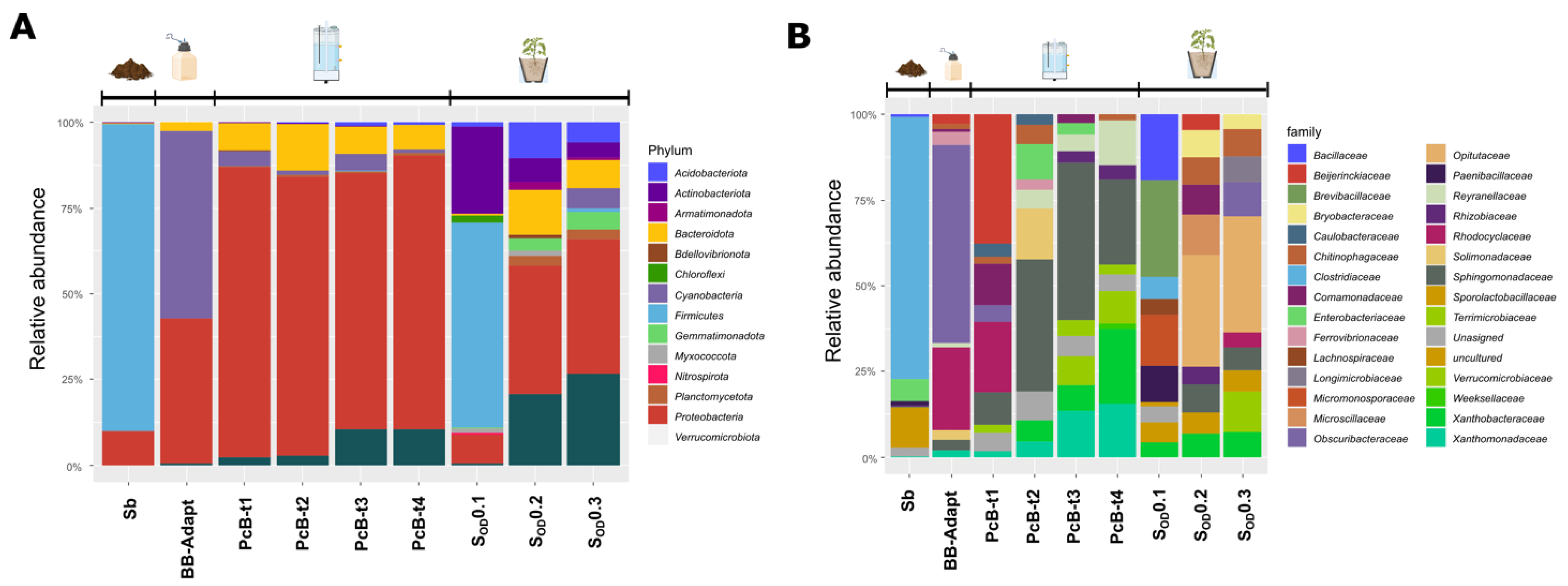
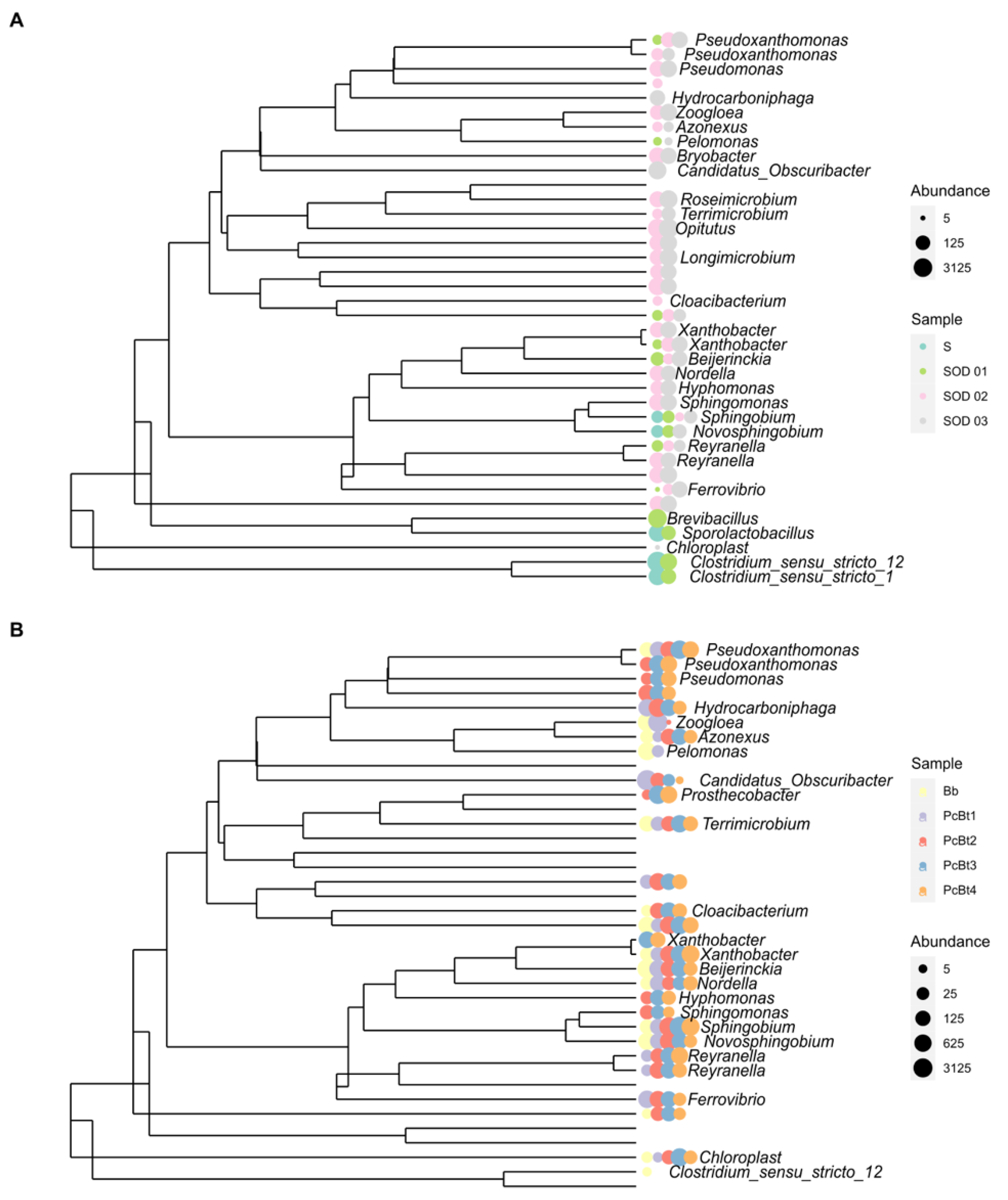
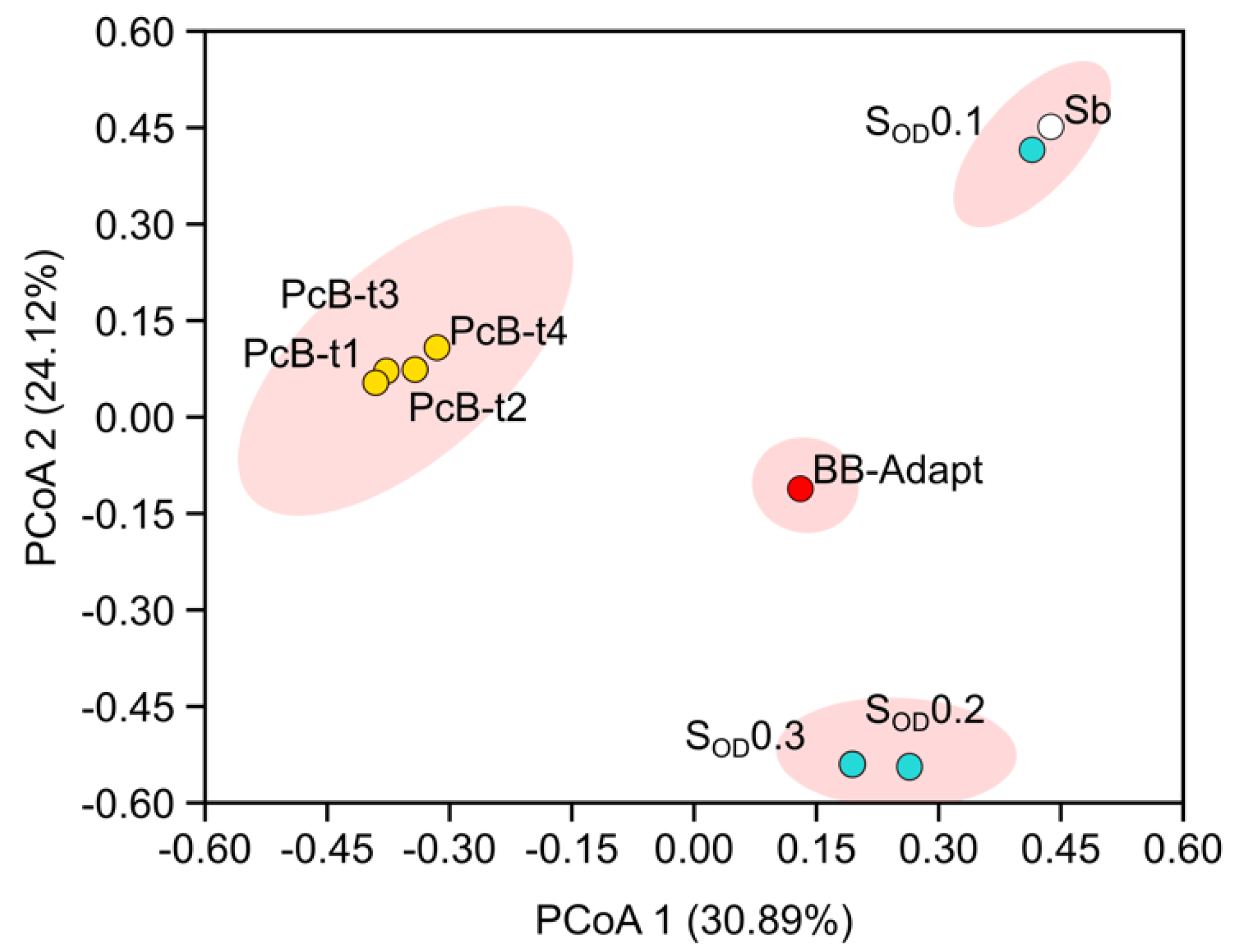
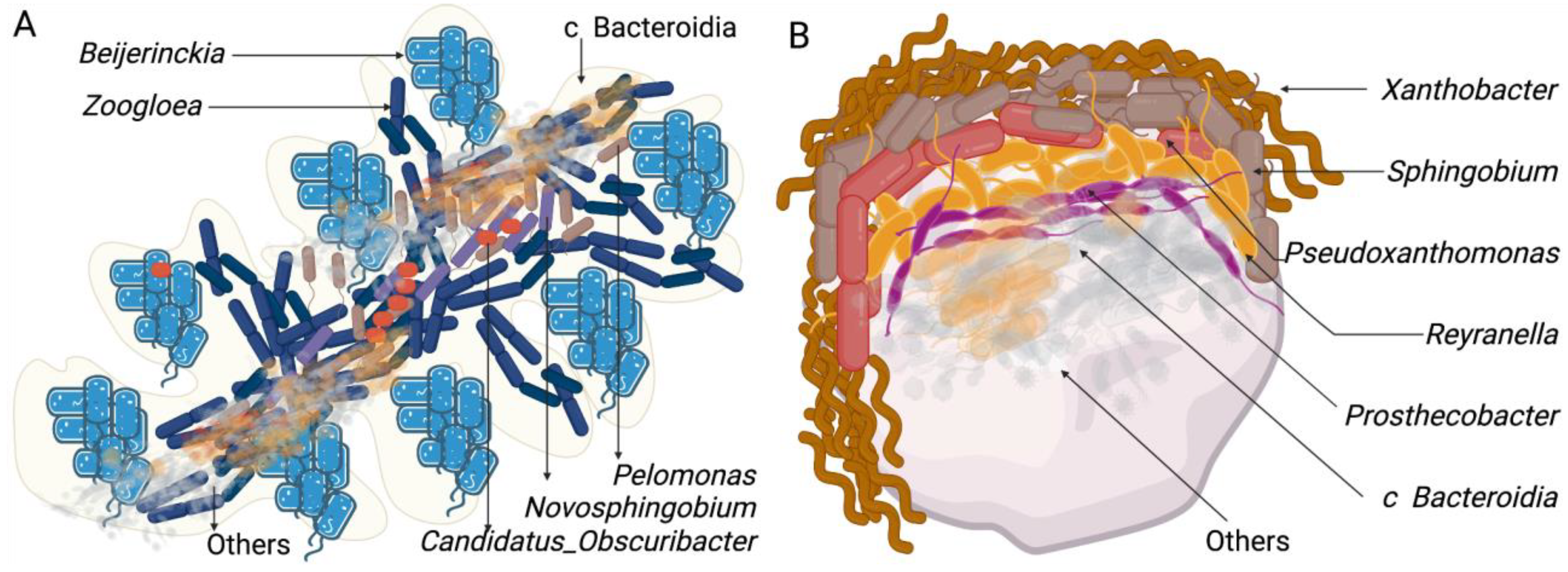
3.3. Composition of the Bacterial Community in Bioreactors
3.4. Composition of the Bacterial Community in the Soil
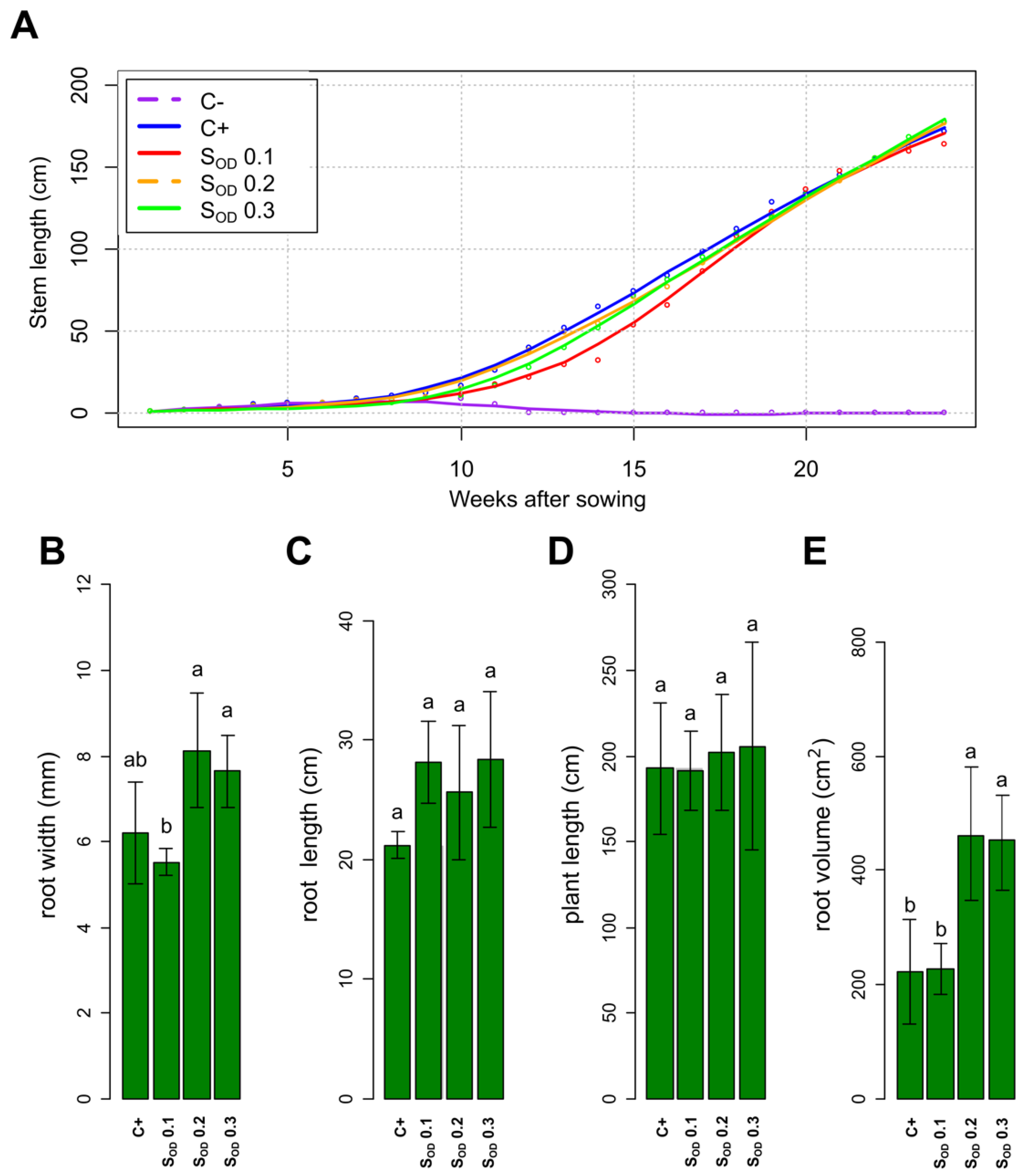
3.5. The Phenotypic Response of the Tomato Plants
4. Discussion
4.1. Bioreactors and Microbial Biodiversity
4.2. Rhizosphere Soil Microbiome and Plant Growth Performance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, M.; Khan, M.A.; Mohsin, I.; Wicks, J.; Ip, A.H.; Sumon, K.Z. Can sustainable ammonia synthesis pathways compete with fossil-fuel based Haber–Bosch processes? Energy Environ. Sci. 2021, 14, 2535–2548. [Google Scholar] [CrossRef]
- Bowles, T.M.; Atallah, S.S.; Campbell, E.E.; Gaudin, A.C.M.; Wieder, W.R.; Grandy, A.S. Addressing agricultural nitrogen losses in a changing climate. Nat. Sustain. 2018, 1, 399–408. [Google Scholar] [CrossRef]
- Fernandes, A.P.G.; Machado, J.; Fernandes, T.R.; Vasconcelos, M.W.; Carvalho, S.M.P. Water and nitrogen fertilization management in light of climate change: Impacts on food security and product quality. In Plant Nutrition and Food Security in the Era of Climate Change; Kumar, V., Srivastava, A.K., Suprasanna, P., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 147–178. [Google Scholar]
- Hunter, M.C.; Smith, R.G.; Schipanski, M.E.; Atwood, L.W.; Mortensen, D.A. Agriculture in 2050: Recalibrating targets for sustainable intensification. Bioscience 2017, 67, 386–391. [Google Scholar] [CrossRef] [Green Version]
- Food & Agriculture Organization. The Future of Food and Agriculture 2018: Alternative Pathways to 2050; Food & Agriculture Organization: Yokohama, Japan, 2018. [Google Scholar]
- Duchene, O.; Vian, J.-F.; Celette, F. Intercropping with legume for agroecological cropping systems: Complementarity and facilitation processes and the importance of soil microorganisms. A review. Agric. Ecosyst. Environ. 2017, 240, 148–161. [Google Scholar] [CrossRef]
- Celestina, C.; Hunt, J.R.; Sale, P.W.G.; Franks, A.E. Attribution of crop yield responses to application of organic amendments: A critical review. Soil Tillage Res. 2019, 186, 135–145. [Google Scholar] [CrossRef]
- Pan, M.; Chu, L.M. Transfer of antibiotics from wastewater or animal manure to soil and edible crops. Environ. Pollut. 2017, 231, 829–836. [Google Scholar] [CrossRef]
- Meemken, E.-M.; Qaim, M. Organic agriculture, food security, and the environment. Annu. Rev. Resour. Econ. 2018, 10, 39–63. [Google Scholar] [CrossRef] [Green Version]
- Timmusk, S.; Behers, L.; Muthoni, J.; Muraya, A.; Aronsson, A.-C. Perspectives and challenges of microbial application for crop improvement. Front. Plant Sci. 2017, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Malusà, E.; Pinzari, F.; Canfora, L. Efficacy of biofertilizers: Challenges to improve crop production. In Microbial Inoculants in Sustainable Agricultural Productivity; Singh, D.P., Singh, H.B., Prabha, R., Eds.; Functional Applications; Springer: New Delhi, India, 2016; Volume 2, pp. 17–40. [Google Scholar]
- Fierer, N.; Ladau, J.; Clemente, J.C.; Leff, J.W.; Owens, S.M.; Pollard, K.S. Reconstructing the microbial diversity and function of pre-agricultural tallgrass prairie soils in the United States. Science 2013, 342, 621–624. [Google Scholar] [CrossRef] [Green Version]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef] [Green Version]
- Sheth, R.U.; Cabral, V.; Chen, S.P.; Wang, H.H. Manipulating bacterial communities by in situ microbiome engineering. Trends Genet. 2016, 32, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the root microbiome by plant molecules: The basis for targeted disease suppression and plant growth promotion. Front. Plant Sci. 2019, 10, 1741. [Google Scholar] [CrossRef]
- Landazuri, C.F.G.; Gomez, J.S.; Raaijmakers, J.M.; Oyserman, B.O. Restoring degraded microbiome function with self-assembled communities. FEMS Microbiol. Ecol. 2020, 96, 12. [Google Scholar] [CrossRef]
- Rodriguez-Gonzalez, C.; Ospina-Betancourth, C.; Sanabria, J. High resistance of a sludge enriched with nitrogen-fixing bacteria to ammonium salts and its potential as a biofertilizer. Bioengineering 2021, 8, 55. [Google Scholar] [CrossRef]
- Ospina-Betancourth, C.; Acharya, K.; Allen, B.; Entwistle, J.; Head, I.M.; Sanabria, J. Enrichment of nitrogen-fixing bacteria in a nitrogen-deficient wastewater treatment system. Environ. Sci. Technol. 2020, 54, 3539–3548. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. Encyclopedia of Soil Science; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Carranzo, I.V. Standard methods for examination of water and wastewater. In Anales De Hidrología Médica; Universidad Complutense de Madrid: Madrid, Spain, 2012; Volume 5, pp. 185–186. Available online: https://search.proquest.com/openview/85626245e0feccc3314bcce5a84957f3/1?pq-origsite=gscholar&cbl=54817 (accessed on 15 February 2020).
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants Without Soil. Circular California Agricultural Experiment Station. Available online: https://www.cabdirect.org/cabdirect/abstract/19500302257 (accessed on 30 August 2019).
- Kamilova, F.; Kravchenko, L.V.; Shaposhnikov, A.I.; Azarova, T.; Makarova, N.; Lugtenberg, B. Organic acids, sugars, and L-tryptophane in exudates of vegetables growing on stonewool and their effects on activities of rhizosphere bacteria. Mol. Plant-Microbe Interact. 2006, 19, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baird, R.; Rice, E.W.; Eaton, A.D.; Bridgewater, L. Water Environment Federation. In Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, WA, USA, 2017. [Google Scholar]
- Barillot, C.D.C.; Sarde, C.-O.; Bert, V.; Tarnaud, E.; Cochet, N. A standardized method for the sampling of rhizosphere and rhizoplan soil bacteria associated to a herbaceous root system. Ann. Microbiol. 2013, 63, 471–476. [Google Scholar] [CrossRef]
- Hall, M.; Beiko, R.G. 16S rRNA gene analysis with QIIME2. Methods Mol. Biol. 2018, 1849, 113–129. [Google Scholar]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods. 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing. 2020. Available online: https://www.R.-project.org/ (accessed on 30 January 2022).
- Hu, B.-L.; Shen, L.-D.; Liu, S.; Cai, C.; Chen, T.-T.; Kartal, B. Enrichment of an anammox bacterial community from a flooded paddy soil. Environ. Microbiol. Rep. 2013, 5, 483–489. [Google Scholar] [CrossRef]
- Kallistova, A.; Nikolaev, Y.; Grachev, V.; Beletsky, A.; Gruzdev, E.; Kadnikov, V. New Insight Into the Interspecies Shift of Anammox Bacteria Ca. “Brocadia” and Ca. “Jettenia” in Reactors Fed With Formate and Folate. Front. Microbiol. 2021, 12, 802201. [Google Scholar] [CrossRef]
- Hess, S.; Suthaus, A.; Melkonian, M. “Candidatus finniella” (rickettsiales, alphaproteobacteria), novel endosymbionts of viridiraptorid amoeboflagellates (cercozoa, rhizaria). Appl. Environ. Microbiol. 2016, 82, 659–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutler, D.W.; Bal, D.V. Influence of protozoa on the process of nitrogen fixation by Azotobacter chroococcum. Ann. Appl. Biol. 1926, 13, 516–534. [Google Scholar] [CrossRef]
- Danso, S.K.; Keya, S.O.; Alexander, M. Protozoa and the decline of rhizobium populations added to soil. Can J. Microbiol. 1975, 21, 884–895. [Google Scholar] [CrossRef] [PubMed]
- Hurek, T.; Reinhold-Hurek, B. Azoarcus sp. strain BH72 as a model for nitrogen-fixing grass endophytes. J. Biotechnol. 2003, 106, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.D.; Lawson, P.A.; Collins, M.D.; Falsen, E.; Tanner, R.S. Cloacibacterium normanense gen. nov., sp. nov., a novel bacterium in the family Flavobacteriaceae isolated from municipal wastewater. Int. J. Syst. Evol. Microbiol. 2006, 56, 1311–1316. [Google Scholar] [CrossRef] [Green Version]
- Kuypers, M.M.M.; Marchant, H.K.; Kartal, B. The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 2018, 16, 263–276. [Google Scholar] [CrossRef]
- Xie, C.-H.; Yokota, A. Zoogloea oryzae sp. nov., a nitrogen-fixing bacterium isolated from rice paddy soil, and reclassification of the strain ATCC 19623 as Crabtreella saccharophila gen. nov., sp. nov. Int. J. Syst. Evol. Microbiol. 2006, 56, 619–624. [Google Scholar] [CrossRef]
- Nielsen, P.H.; Thomsen, T.R.; Nielsen, J.L. Bacterial composition of activated sludge-importance for floc and sludge properties. Water Sci. Technol. 2004, 49, 51–58. [Google Scholar] [CrossRef]
- Becking, J.H. The genus beijerinckia. In The Prokaryotes: Proteobacteria: Alpha and Beta Subclasses; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 5, pp. 151–162. [Google Scholar]
- Gomila, M.; Gascó, J.; Busquets, A.; Gil, J.; Bernabeu, R.; Buades, J.M. Identification of culturable bacteria present in haemodialysis water and fluid. FEMS Microbiol. Ecol. 2005, 52, 101–114. [Google Scholar] [CrossRef] [Green Version]
- Soo, R.M.; Skennerton, C.T.; Sekiguchi, Y.; Imelfort, M.; Paech, S.J.; Dennis, P.G. An expanded genomic representation of the phylum cyanobacteria. Genome Biol. Evol. 2014, 6, 1031–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Kim, D.-U.; Lee, S.; Park, S.; Yoon, J.-H.; Seong, C.N. Reyranella terrae sp. nov., isolated from an agricultural soil, and emended description of the genus reyranella. Int. J. Syst. Evol. Microbiol. 2017, 67, 2031–2035. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sakimoto, K.K.; Colón, B.C.; Silver, P.A.; Nocera, D.G. Ambient nitrogen reduction cycle using a hybrid inorganic-biological system. Proc. Natl. Acad. Sci. USA 2017, 114, 6450–6455. [Google Scholar] [CrossRef] [Green Version]
- Bu, X.; Gu, X.; Zhou, X.; Zhang, M.; Guo, Z.; Zhang, J. Extreme drought slightly decreased soil labile organic C and N contents and altered microbial community structure in a subtropical evergreen forest. For. Ecol. Manag. 2018, 429, 18–27. [Google Scholar] [CrossRef]
- Welz, P.J.; Ramond, J.-B.; Braun, L.; Vikram, S.; Le Roes-Hill, M. Bacterial nitrogen fixation in sand bioreactors treating winery wastewater with a high carbon to nitrogen ratio. J. Environ. Econ. Manag. 2018, 207, 192–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Li, Y.; Wu, J. Bacterial community response to different nitrogen gradients of swine wastewater in surface flow constructed wetlands. Chemosphere 2021, 265, 129106. [Google Scholar] [CrossRef]
- Bao, Y.; Dolfing, J.; Guo, Z.; Chen, R.; Wu, M.; Li, Z. Important ecophysiological roles of non-dominant actinobacteria in plant residue decomposition, especially in less fertile soils. Microbiome 2021, 9, 84. [Google Scholar] [CrossRef]
- Lima, J.V.; Tinôco, R.S.; Olivares, F.L.; Chia, G.S.; de Melo Júnior, J.A.G.; da Silva, G.B. Rhizobacteria modify root architecture and improve nutrient uptake in oil palm seedlings despite reduced fertilizer. Rhizosphere 2021, 19, 100420. [Google Scholar] [CrossRef]
- Li, B.; Zhang, C.; Qi, M.; Zheng, X.; Mustafad, N.S.; Ahmed, N. Effects of plant growth-promoting rhizobacteria on uptake and utilization of phosphorus and root architecture in apple seedlings under water limited regimes. Int. J. Appl. Exp. Biol. 2022, 1, 1–8. [Google Scholar]
- Herms, C.H.; Hennessy, R.C.; Bak, F.; Dresbøll, D.B.; Nicolaisen, M.H. Back to our roots: Exploring the role of root morphology as a mediator of beneficial plant-microbe interactions. Environ. Microbiol. 2022. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez, C.F.; Rodríguez-Romero, N.; Egan, S.; Holmes, E.; Sanabria, J. Exploiting the Potential of Bioreactors for Creating Spatial Organization in the Soil Microbiome: A Strategy for Increasing Sustainable Agricultural Practices. Microorganisms 2022, 10, 1464. https://doi.org/10.3390/microorganisms10071464
Gutiérrez CF, Rodríguez-Romero N, Egan S, Holmes E, Sanabria J. Exploiting the Potential of Bioreactors for Creating Spatial Organization in the Soil Microbiome: A Strategy for Increasing Sustainable Agricultural Practices. Microorganisms. 2022; 10(7):1464. https://doi.org/10.3390/microorganisms10071464
Chicago/Turabian StyleGutiérrez, Carlos Fernando, Nicolás Rodríguez-Romero, Siobhon Egan, Elaine Holmes, and Janeth Sanabria. 2022. "Exploiting the Potential of Bioreactors for Creating Spatial Organization in the Soil Microbiome: A Strategy for Increasing Sustainable Agricultural Practices" Microorganisms 10, no. 7: 1464. https://doi.org/10.3390/microorganisms10071464
APA StyleGutiérrez, C. F., Rodríguez-Romero, N., Egan, S., Holmes, E., & Sanabria, J. (2022). Exploiting the Potential of Bioreactors for Creating Spatial Organization in the Soil Microbiome: A Strategy for Increasing Sustainable Agricultural Practices. Microorganisms, 10(7), 1464. https://doi.org/10.3390/microorganisms10071464