
Whole Genome Sequencing and Molecular Analysis of Carbapenemase-Producing Escherichia coli from Intestinal Carriage in Elderly Inpatients


 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain Identification and Antimicrobial Susceptibility Testing (AST)
2.2. Whole Genome Sequencing and In Silico Analysis
3. Results
3.1. Antimicrobial Susceptibility Testing and Characterization of Resistance Genes
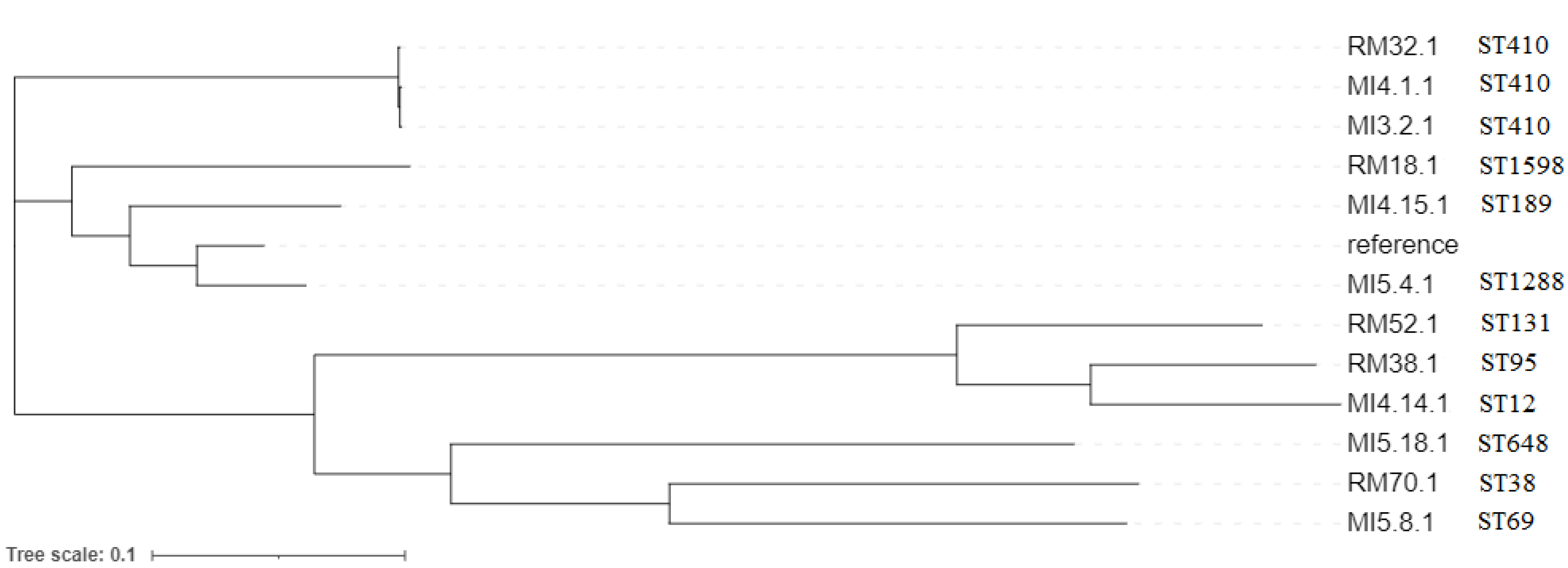
3.2. WGS Analysis of CP-E. coli
3.3. Plasmid Content of CP-E. coli
3.4. Resistome of CP-E. coli
3.5. Virulence Genes of CP-E. coli
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef] [Green Version]
- Latour, K.; Huang, T.D.; Jans, B.; Berhin, C.; Bogaerts, P.; Noel, A.; Nonhoff, C.; Dodémont, M.; Denis, O.; Ieven, M.; et al. Prevalence of multidrug-resistant organisms in nursing homes in Belgium in 2015. PLoS ONE 2019, 14, e0214327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorrie, C.L.; Mirceta, M.; Wick, R.R.; Judd, L.M.; Wyres, K.L.; Thomson, N.R.; Strugnell, R.A.; Pratt, N.F.; Garlick, J.S.; Watson, K.M.; et al. Antimicrobial-Resistant Klebsiella pneumoniae Carriage and Infection in Specialized Geriatric Care Wards Linked to Acquisition in the Referring Hospital. Clin. Infect. Dis. 2018, 67, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Tumbarello, M.; Viale, P.; Viscoli, C.; Trecarichi, E.M.; Tumietto, F.; Marchese, A.; Spanu, T.; Ambretti, S.; Ginocchio, F.; Cristini, F.; et al. Predictors of mortality in bloodstream infections caused by Klebsiella pneumoniae carbapenemase-producing K. pneumoniae: Importance of combination therapy. Clin. Infect. Dis. 2012, 55, 943–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Regional Office for Europe/European Centre for Disease Prevention and Control. Antimicrobial Resistance Surveillance in Europe 2022—2020 Data; WHO Regional Office for Europe: Copenhagen, Denmark, 2022. [Google Scholar]
- Jiang, Y.; Wang, Y.; Hua, X.; Qu, Y.; Peleg, A.Y.; Yu, Y. Pooled plasmid sequencing reveals the relationship between mobile genetic elements and antimicrobial resistance genes in clinically isolated Klebsiella pneumoniae. Genom. Proteom. Bioinform. 2020, 18, 539–548. [Google Scholar] [CrossRef]
- Kiratisin, P.; Kazmierczak, K.; Stone, G.G. In vitro activity of ceftazidime/avibactam and comparators against carbapenemase-producing Enterobacterales and Pseudomonas aeruginosa isolates collected globally between 2016 and 2018. J. Glob. Antimicrob. Resist. 2021, 27, 132–141. [Google Scholar] [CrossRef]
- Yu, W.; Xiong, L.; Luo, Q.; Chen, Y.; Ji, J.; Ying, C.; Liu, Z.; Xiao, Y. In Vitro Activity Comparison of Ceftazidime-Avibactam and Aztreonam-Avibactam against Bloodstream Infections with Carbapenem-Resistant Organisms in China. Front. Cell Infect. Microbiol. 2021, 11, 780365. [Google Scholar] [CrossRef] [PubMed]
- Di Bella, S.; Giacobbe, D.R.; Maraolo, A.E.; Viaggi, V.; Luzzati, R.; Bassetti, M.; Luzzaro, F.; Principe, L. Resistance to ceftazidime/avibactam in infections and colonisations by KPC-producing Enterobacterales: A systematic review of observational clinical studies. J. Glob. Antimicrob. Resist. 2021, 25, 268–281. [Google Scholar] [CrossRef]
- Pakbin, B.; Brück, W.M.; Rossen, J.W.A. Virulence Factors of Enteric Pathogenic Escherichia coli: A Review. Int. J. Mol. Sci. 2021, 22, 9922. [Google Scholar] [CrossRef] [PubMed]
- Gruber, I.; Heudorf, U.; Werner, G.; Pfeifer, Y.; Imirzalioglu, C.; Ackermann, H.; Brandt, C.; Besier, S.; Wichelhaus, T.A. Multidrug-resistant bacteria in geriatric clinics, nursing homes, and ambulant cared prevalence and risk factors. Int. J. Med. Microbiol. 2013, 303, 405.e9. [Google Scholar] [CrossRef]
- Lim, C.J.; Cheng, A.C.; Kennon, J.; Spelman, D.; Hale, D.; Melican, G.; Sidjabat, H.E.; Paterson, D.L.; Kong, D.C.M.; Peleg, A.Y. Prevalence of multidrug-resistant organisms and risk factors for carriage in long-term care facilities: A nested caseecontrol study. J. Antimicrob. Chemother. 2014, 69, 1972–1980. [Google Scholar] [CrossRef] [PubMed]
- Aschbacher, R.; Pagani, E.; Confalonieri, M.; Farina, C.; Fazii, P.; Luzzaro, F.; Montanera, P.G.; Piazza, A.; Pagani, L. Review on colonization of residents and staff in Italian long-term care facilities by multidrug-resistant bacteria compared with other European countries. Antimicrob. Resist. Infect. Control 2016, 5, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinelli, M.; Rossini, A.; Scudeller, L.; Zabzuni, D.; Errico, G.; Fogato, E.; D’Angelo, R.; Gentiloni Silverj, F.; Cesana, E.; Bergamaschini, L.C. Dynamics of carbapenemase-producing Enterobacterales intestinal colonization in the elderly population after hospital discharge, Italy, 2018–2020. Int. J. Antimicrob. Agents 2022, 59, 106594. [Google Scholar] [CrossRef] [PubMed]
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 12.0; 2022. Available online: http://www.eucast.org (accessed on 8 April 2022).
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Grundmann, H.; Glasner, C.; Albiger, B.; Aanensen, D.M.; Tomlinson, C.T.; Andrasević, A.T.; Cantón, R.; Carmeli, Y.; Friedrich, A.W.; Giske, C.G. Occurrence of carbapenemase-producing Klebsiella pneumoniae and Escherichia coli in the European survey of carbapenemase-producing Enterobacteriaceae (EuSCAPE): A prospective, multinational study. Lancet Infect. Dis. 2017, 17, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Viale, P.; Tumietto, F.; Giannella, M.; Bartoletti, M.; Tedeschi, S.; Ambretti, S.; Cristini, F.; Gibertoni, C.; Venturi, S.; Cavalli, M.; et al. Impact of a hospital–wide multifaceted programme for reducing carbapenem-resistant Enterobacteriaceae infections in a large teaching hospital in northern Italy. Clin. Microbiol. Infect. 2015, 21, 242–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giufrè, M.; Mazzolini, E.; Cerquetti, M.; Brusaferro, S. CCM2015 One-Health ESBL-producing Escherichia coli Study Group. Extended-spectrum β-lactamase-producing Escherichia coli from extraintestinal infections in humans and from food-producing animals in Italy: A ‘One Health’ study. Int. J. Antimicrob. Agents 2021, 58, 106433. [Google Scholar] [CrossRef]
- Giufrè, M.; Errico, G.; Accogli, M.; Monaco, M.; Villa, L.; Distasi, M.A.; Del Gaudio, T.; Pantosti, A.; Carattoli, A.; Cerquetti, M. Emergence of NDM-5 producing Escherichia coli sequence type 167 clone in Italy. Int. J. Antimicrob. Agents 2018, 52, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Giufrè, M.; Accogli, M.; Ricchizzi, E.; Barbanti, F.; Farina, C.; Fazii, P.; Mattei, R.; Sarti, M.; Barozzi, A.; Buttazzi, R.; et al. Multidrug-resistant infections in long-term care facilities: Extended-spectrum β-lactamase-producing Enterobacteriaceae and hypervirulent antibiotic resistant Clostridium difficile. Diagn. Microbiol. Infect. Dis. 2018, 91, 275–281. [Google Scholar] [CrossRef]
- Giufrè, M.; Ricchizzi, E.; Accogli, M.; Barbanti, F.; Monaco, M.; Pimentel de Araujo, F.; Farina, C.; Fazii, P.; Mattei, R.; Sarti, M.; et al. Colonization by multidrug-resistant organisms in long-term care facilities in Italy: A point-prevalence study. Clin. Microbiol. Infect. 2017, 23, 961–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thaden, J.T.; Pogue, J.M.; Kaye, K.S. Role of newer and re-emerging older agents in the treatment of infections caused by carbapenem-resistant Enterobacteriaceae. Virulence 2017, 8, 403–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsson, M.; Lutgring, J.D.; Ansari, U.; Lawsin, A.; Albrecht, V.; McAllister, G.; Daniels, J.; Lonsway, D.; McKay, S.; Beldavs, Z.; et al. Molecular Characterization of Carbapenem-Resistant Enterobacterales Collected in the United States. Microb. Drug Resist. 2022, 28, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, K.L.; Ellaby, N.; Ellington, M.J.; Doumith, M.; Mustafa, N.; Meunier, D.; Woodford, N. Diversity of carbapenemase-producing Enterobacterales in England as revealed by whole-genome sequencing of isolates referred to a national reference laboratory over a 30-month period. J. Med. Microbiol. 2022, 71, 001518. [Google Scholar] [CrossRef] [PubMed]
- Nicolas-Chanoine, M.H.; Bertrand, X.; Madec, J.Y. Escherichia coli ST131, an intriguing clonal group. Clin. Microbiol. Rev. 2014, 27, 543–574. [Google Scholar] [CrossRef] [Green Version]
- Peirano, G.; Bradford, P.A.; Kazmierczak, K.M.; Badal, R.E.; Hackel, M.; Hoban, D.J.; Pitout, J.D. Global incidence of carbapenemase-producing Escherichia coli ST131. Emerg. Infect. Dis. 2014, 20, 1928–1931. [Google Scholar] [CrossRef] [PubMed]
- Roer, L.; Overballe-Petersen, S.; Hansen, F.; Hansen, F.; Schønning, K.; Wang, M.; Hansen, D.S.; Justesen, U.S.; Andersen, L.P.; Fulgsang-Damgaard, D.; et al. Escherichia coli Sequence Type 410 is causing new international high-risk clones. mSphere 2018, 3, e00337-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global extraintestinal pathogenic Escherichia coli (ExPEC) lineages. Clin. Microbiol. Rev. 2019, 32, e00135-18. [Google Scholar] [CrossRef]
- Adams-Sapper, S.; Diep, B.A.; Perdreau-Remington, F.; Riley, L.W. Clonal composition and community clustering of drug-susceptible and resistant Escherichia coli isolates from blood stream infections. Antimicrob. Agents Chemother. 2013, 57, 490–497. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.D.; Irshad, P.; Anusree, M.; Rekha, I.; Shailaja, S.; Suresh, J.; Aishwarya, G.; Shrestha, S.; Shome, B.R. Whole genome global insight of antibiotic resistance gene repertoire and virulome of high-risk multidrug-resistant Uropathogenic Escherichia coli. Microb. Pathog. 2021, 161, 105256. [Google Scholar] [CrossRef] [PubMed]
- Kopotsa, K.; Osei Sekyere, J.; Mbelle, N.M. Plasmid evolution in carbapenemase-producing Enterobacteriaceae: A review. Ann. N. Y. Acad. Sci. 2019, 1457, 61–91. [Google Scholar] [CrossRef]
- Sora, V.M.; Meroni, G.; Martino, P.A.; Soggiu, A.; Bonizzi, L.; Zecconi, A. Extraintestinal Pathogenic Escherichia coli: Virulence Factors and Antibiotic Resistance. Pathogens 2021, 10, 1355. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, S.; Xia, Y.; Sondén, B.; Göransson, M.; Hacker, J.; Uhlin, B.E. Regulatory Interactions among adhesin gene systems of uropathogenic Escherichia coli. Infect. Immun. 2008, 76, 771–780. [Google Scholar] [CrossRef] [Green Version]
- Chu, B.C.; Garcia-Herrero, A.; Johanson, T.H.; Krewulak, K.D.; Lau, C.K.; Peacock, R.S.; Slavinskaya, Z.; Vogel, H.J. Siderophore uptake in bacteria and the battle for iron with the host; a bird’s eye view. Biometals 2010, 23, 601–611. [Google Scholar] [CrossRef]
- Chen, B.; Berglund, B.; Wang, S.; Börjesson, S.; Bi, Z.; Nilsson, M.; Yin, H.; Zheng, B.; Xiao, Y.; Bi, Z.; et al. Rapid increase in occurrence of carbapenem-resistant Enterobacteriaceae in healthy rural residents in Shandong Province, China, from 2015 to 2017. J. Glob. Antimicrob. Resist. 2022, 28, 38–42. [Google Scholar] [CrossRef]
- Liu, X.; Li, X.; Yang, A.W.; Tang, B.; Jian, Z.J.; Zhong, Y.M.; Li, H.L.; Li, Y.M.; Yan, Q.; Liang, X.H.; et al. Community Fecal Carriage and Molecular Epidemiology of Extended-Spectrum β-Lactamase- and Carbapenemase-Producing Escherichia coli from Healthy Children in the Central South China. Infect. Drug Resist. 2022, 15, 1601–1611. [Google Scholar] [CrossRef] [PubMed]
- Yen, N.T.P.; Nhung, N.T.; Phu, D.H.; Dung, N.T.T.; Van, N.T.B.; Kiet, B.T.; Hien, V.B.; Larsson, M.; Olson, L.; Campbell, J.; et al. Prevalence of carbapenem resistance and its potential association with antimicrobial use in humans and animals in rural communities in Vietnam. JAC Antimicrob. Resist. 2022, 4, dlac038. [Google Scholar] [CrossRef] [PubMed]
- Hernández-García, M.; Pérez-Viso, B.; Carmen Turrientes, M.; Díaz-Agero, C.; López-Fresneña, N.; Bonten, M.; Malhotra-Kumar, S.; Ruiz-Garbajosa, P.; Cantón, R. Characterization of carbapenemase-producing Enterobacteriaceae from colonized patients in a university hospital in Madrid, Spain, during the R-GNOSIS project depicts increased clonal diversity over time with maintenance of high-risk clones. J. Antimicrob. Chemother. 2018, 73, 3039–3043. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Isolate | Serotype | ST | CC | fimH | Plasmid Content | pMLST * | β-Lactamases | Aminoglycoside Modifying Enzimes | FQs | MLS | Phenicol | Sulphonamide | TET | Trimethoprim |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MI3.2.1 | O8:H9 | 410 | ST23 | 24 | IncFIA, IncFIB, IncFII, IncQ1 | IncF[F1:A1:B49] | NDM-5, CTX-M-15, CMY-2, OXA-1, TEM-1B | aac(6′)-Ib-cr, aadA2, aadA5, aph(6)-Id, aph(3″)-Ib, aac(3)-IId | aac(6′)-Ib-cr | mph(A) | catB3 | sul1, sul2 | tet(B) | dfrA12, dfrA17 |
| MI4.1.1 | O8:H9 | 410 | ST23 | 24 | IncFIA, IncFIB, IncFII, Col (BS512) | IncF[F1:A1:B49] | NDM-5, CTX-M-15, CMY-2 | aac(6′)-Ib-cr, aadA2, aph(6)-Id, aph(3″)-Ib, aac(3)-IId | aac(6′)-Ib-cr | mph(A) | sul1, sul2 | dfrA12 | ||
| RM32.1 | O8:H9 | 410 | ST23 | 21 | IncFIA, IncFIB, IncFII, IncQ1, IncX3, IncY, Col (BS512), ColKP3 | IncF[F1:A1:B49] | OXA-181, CTX-M-15CMY-2, OXA-1, TEM-1B | aac(6′)-Ib-cr, aadA5, aph(6)-Id, aph(3″)-Ib, aac(3)-IId | aac(6′)-Ib-cr, qnrS1 | mph(A) | catB3 | sul1, sul2 | tet(B) | dfrA17 |
| MI4.14.1 | -:H5 | 12 | ST12 | 5 | IncFIA, IncFIB, IncFII, IncX1, IncB/O/K/Z, Col156, ColKP3 | IncF[F1:A6:B20] | OXA-181, TEM-1B | aadA5, aph(6)-Id, aph(3″)-Ib, aac(3)-IId | mph(A) | sul1, sul2 | tet(A), tet(C) | dfrA17 | ||
| MI4.15.1 | O188:H21 | 189 | ST165 | 54 | IncX3 | KPC-3, SHV-182 | aadA5 | sul2 | tet(A) | dfrA17 | ||||
| MI5.4.1 | O9:H9 | 1288 | - | 54 | IncFIB, IncFII, ColpVC | IncF[K2:A-:B-] | KPC-2 | aph(3″)-Ib, aph(6)-Id | tet(B) | |||||
| MI5.8.1 | O15:H18 | 69 | ST69 | 27 | IncA, IncB/O/K/Z | IncA/C [12] | VIM-1, SHV-12 | aac(6′)-Ib-cr, aadA1, aph(6)-Id, aph(3″)-Ib, aac(6′)-Ib3 | aac(6′)-Ib-cr, qnrS1 | mph(A) | catB2 | sul1, sul2 | dfrA14 | |
| MI5.18.1 | -:H6 | 648 | ST648 | 27 | IncFIB, IncFII | IncF[K2:A-:B-] | KPC-2, CTX-M-15, OXA-9 | |||||||
| RM18.1 | O9:H4 | 1598 | - | - | IncFIB, IncFII, IncI1-I, IncR, IncX3, Col (BS512) | IncF[K2:A-:B53], IncI1[154] | KPC-3, CTX-M-15, OXA-9, SHV-182, TEM-1B | aph(3″)-Ib, aph(6)-Id | qnrS1 | sul2 | tet(A) | dfrA14 | ||
| RM38.1 | O1:H7 | 95 | ST95 | 30 | IncFIB, IncFII | IncF[F2:A-:B1] | KPC-3, OXA-9, TEM-1A | |||||||
| RM52.1 | O25:H4 | 131 | ST131 | 30 | IncFIA, IncFIB, IncFII | IncF[K2:A4:B1] | KPC-3, CTX-M-15, OXA-1, OXA-9, TEM-1A | aac(6′)-Ib-cr, aadA5, aac(3)-IIa | aac(6′)-Ib-cr | mph(A) | catB3 | sul1 | tet(A) | dfrA17 |
| RM70.1 | -:H9 | 38 | ST38 | 24 | IncFIB, IncFII | IncF[K2:A-:B-] | KPC-3, OXA-9, TEM-1A |
| Isolate | Adhesins | Capsule | Siderophores | Serum Resistance | Toxins | Other |
|---|---|---|---|---|---|---|
| MI3.2.1 | fimH, lpfA | gad, terC | ||||
| MI4.1.1 | fimH, lpfA | gad, terC | ||||
| RM32.1 | fimH, lpfA | gad, terC | ||||
| MI4.14.1 | fimH, focC, papA/C, sfaD, yfcV | kpsE/MII | chuA, fyuA, iroN, irp2, sitA | iss, ompT, traT | cea, senB, usp, vat | gad, terC |
| MI4.15.1 | fimH, lpfA | gad, terC | ||||
| MI5.4.1 | fimH | sitA | traT | gad, terC | ||
| MI5.8.1 | fimH, lpfA | kpsE/MII | chuA, fyuA, irp2, sitA | iss, ompT, traT | gad, terC | |
| MI5.18.1 | fimH, lpfA, papA/C, yfcV | kpsE/MII, neuC | chuA, fyuA, irp2 | ompT, traT | gad, terC | |
| RM18.1 | sitA | iss, traT | gad, terC | |||
| RM38.1 | fimH, papA/C, yfcV | kpsE/MII, neuC | chuA, fyuA, ireA, iroN, irp2, iucC, sitA | iss, ompT, traT | cia, hlyF, usp, vat | gad, terC |
| RM52.1 | afaA/C/D, fimH, hra, iha, nfaE, papA/C, yfcV | kpsE/MII | chuA, fyuA, irp2, iucC, sitA | iss, ompT, traT | cnf1, sat, usp | gad, terC |
| RM70.1 | fimH, hra | kpsE/MII | chuA, sitA | traT | gad, terC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giufrè, M.; Errico, G.; Monaco, M.; Del Grosso, M.; Sabbatucci, M.; Pantosti, A.; Cerquetti, M.; Pagnotta, M.; Marra, M.; Carollo, M.; et al. Whole Genome Sequencing and Molecular Analysis of Carbapenemase-Producing Escherichia coli from Intestinal Carriage in Elderly Inpatients. Microorganisms 2022, 10, 1561. https://doi.org/10.3390/microorganisms10081561
Giufrè M, Errico G, Monaco M, Del Grosso M, Sabbatucci M, Pantosti A, Cerquetti M, Pagnotta M, Marra M, Carollo M, et al. Whole Genome Sequencing and Molecular Analysis of Carbapenemase-Producing Escherichia coli from Intestinal Carriage in Elderly Inpatients. Microorganisms. 2022; 10(8):1561. https://doi.org/10.3390/microorganisms10081561
Chicago/Turabian StyleGiufrè, Maria, Giulia Errico, Monica Monaco, Maria Del Grosso, Michela Sabbatucci, Annalisa Pantosti, Marina Cerquetti, Michela Pagnotta, Manuela Marra, Maria Carollo, and et al. 2022. "Whole Genome Sequencing and Molecular Analysis of Carbapenemase-Producing Escherichia coli from Intestinal Carriage in Elderly Inpatients" Microorganisms 10, no. 8: 1561. https://doi.org/10.3390/microorganisms10081561
APA StyleGiufrè, M., Errico, G., Monaco, M., Del Grosso, M., Sabbatucci, M., Pantosti, A., Cerquetti, M., Pagnotta, M., Marra, M., Carollo, M., Rossini, A., Fogato, E., Cesana, E., Gentiloni Silverj, F., Zabzuni, D., & Tinelli, M. (2022). Whole Genome Sequencing and Molecular Analysis of Carbapenemase-Producing Escherichia coli from Intestinal Carriage in Elderly Inpatients. Microorganisms, 10(8), 1561. https://doi.org/10.3390/microorganisms10081561