
Bacillus velezensis QA2 Potentially Induced Salt Stress Tolerance and Enhanced Phosphate Uptake in Quinoa Plants




Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling
2.2. Isolation, Screening, and Purification of Phosphate Solubilizing Rhizobacteria on Plates
2.3. Screening for Salt Tolerance
2.4. Quantitative Assay of P Solubilization in Liquid Media
2.5. DNA Amplification and Phylogenetic Identification of Selected Rhizobacterium
2.6. In Vitro Evaluation of PGP Traits
2.6.1. Indole-3-Acetic Acid (IAA) Production Assay
2.6.2. Biofilm Formation Assay
2.6.3. Qualitative Siderophores Production Assay
2.6.4. Ammonia Production Assay
2.6.5. Zinc Solubilization Assay
2.6.6. HCN Production Assay
2.6.7. Extracellular Enzymes Production Assay
2.6.8. Antibacterial Activity Assay
2.6.9. Antifungal Activity Assay
2.6.10. Bacterial Antibiotic Resistance Pattern
2.7. In-Vivo Inoculation Experiments
2.7.1. Seed Germination Assay
- Germination rate (%) = (Number of germinated seeds/Total number of seeds) × 100
- Vigor index = Germination rate (%) × Total seedling length (cm)
2.7.2. In Vivo Pot Experiment under Saline Irrigation Conditions
Inoculant Preparation and Seeds Treatment
Irrigation Treatment
Plant Vegetative Attributes Measurement
Plant Physiological Attributes Analysis
2.8. Bibliometric Analysis of Bacillus velezensis Strains
2.9. Statistical Analysis
3. Results
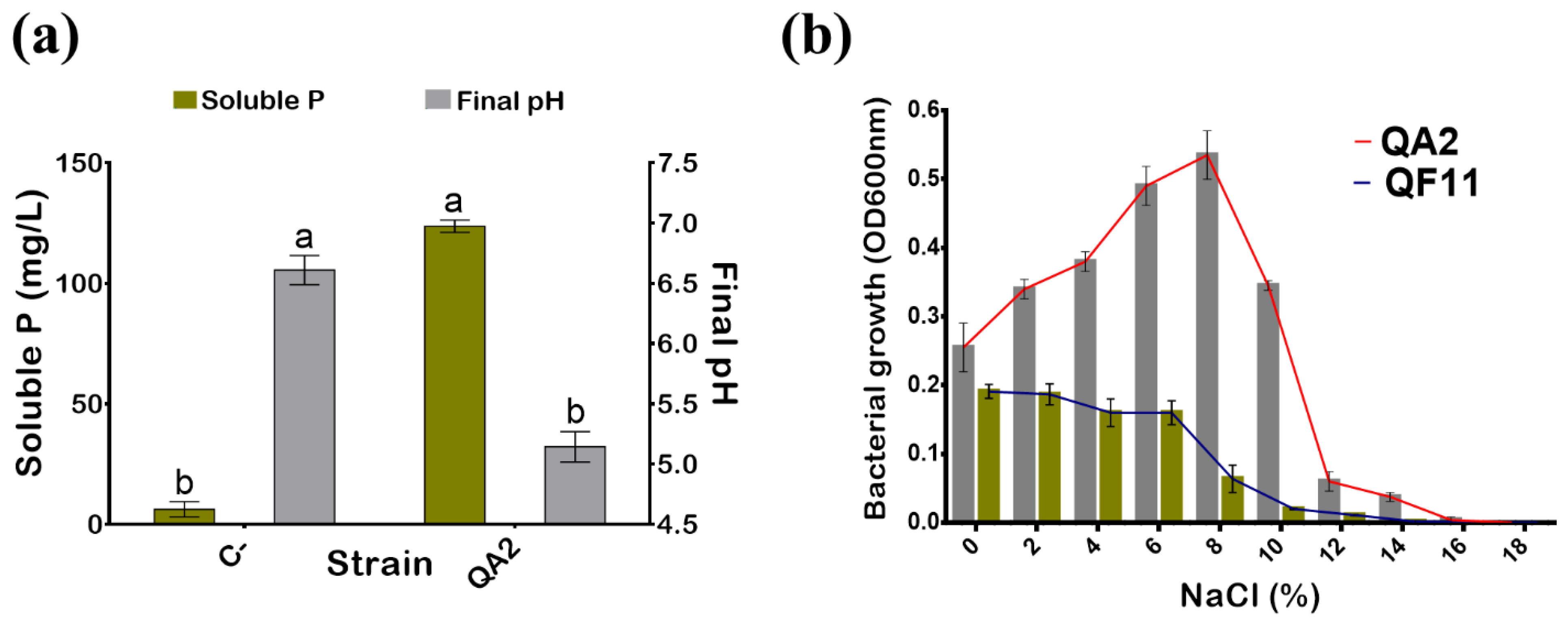
3.1. The QA2 Isolate Solubilizes a High Level of Phosphate
3.2. The QA2 Isolate Is Halotolerant, Mesophilic, and Sensitive to Antibiotics
3.3. The QA2 Isolate Is Endowed with High IAA Production and Exhibits a Low Zinc Solubilization Activity
3.4. The QA2 Isolate Is a High Siderophores Producer
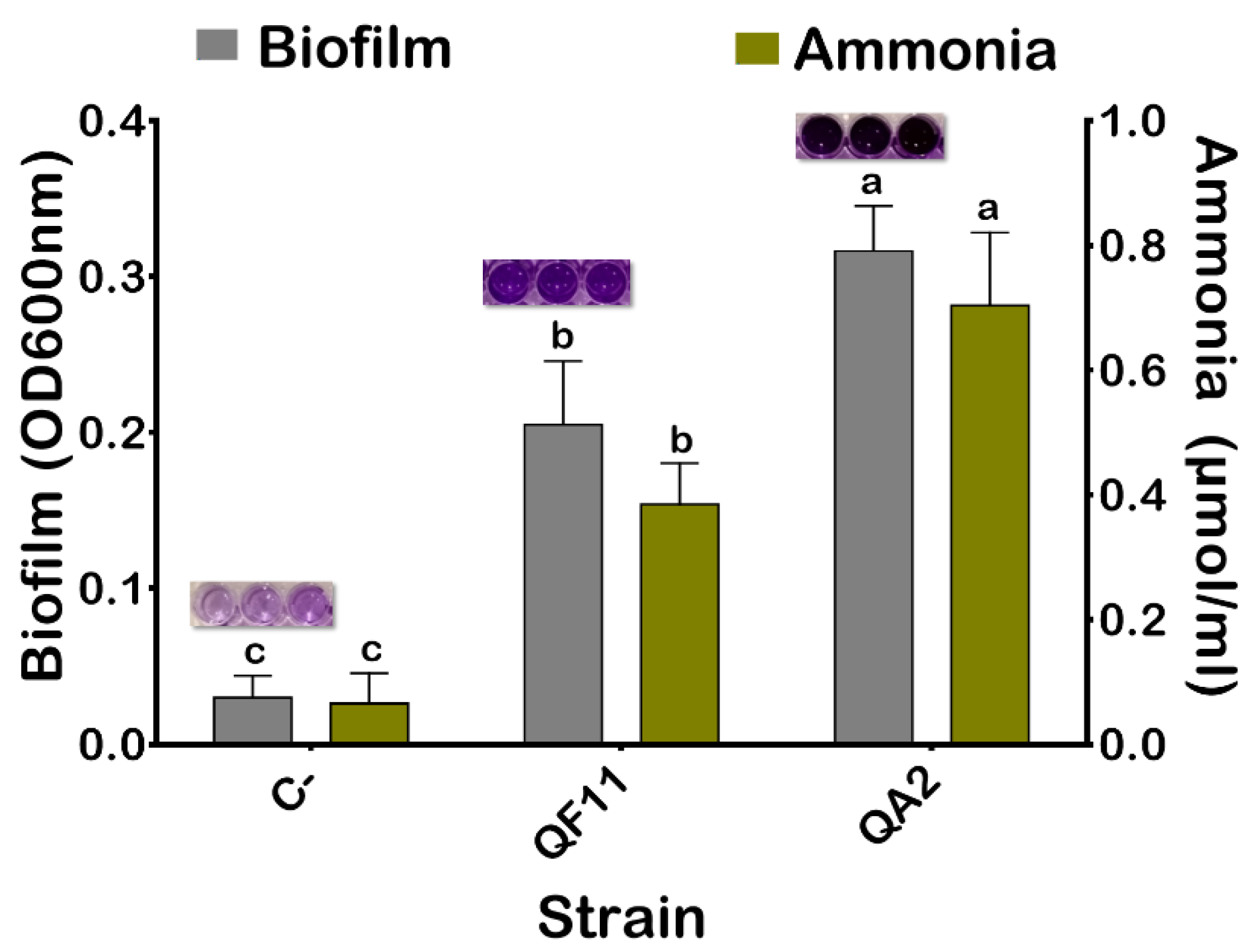
3.5. The QA2 Isolate Is a Strong Biofilm Producer
3.6. The QA2 Isolate Produces a High amount of Ammonia and HCN and Elicited Anti-Pathogenic Bacterial Activity
3.7. The QA2 Isolate Inhibits Fungal Growth and Overproduces Extracellular Enzymes
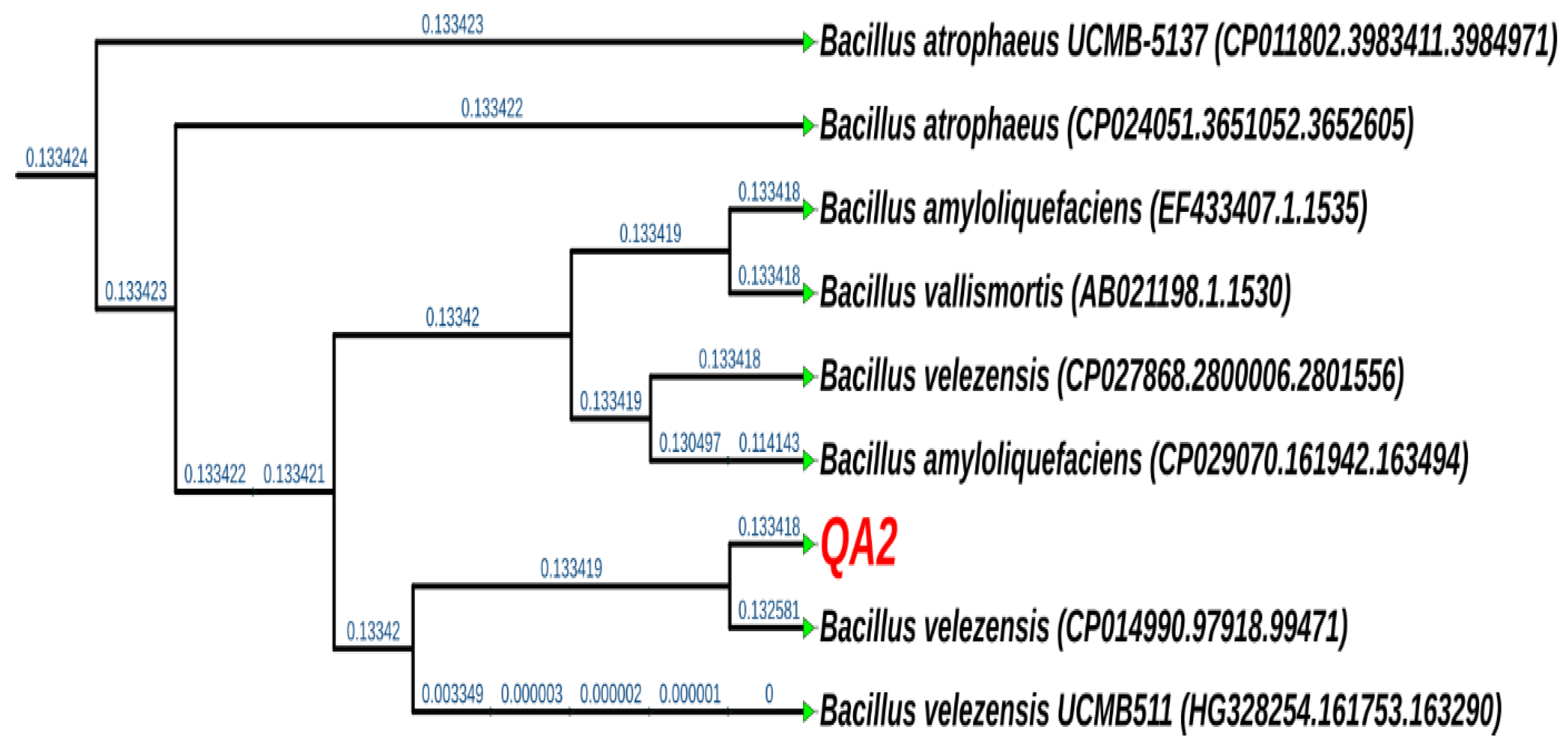
3.8. The QA2 Isolate Belongs to the Genus of Bacillus velezensis
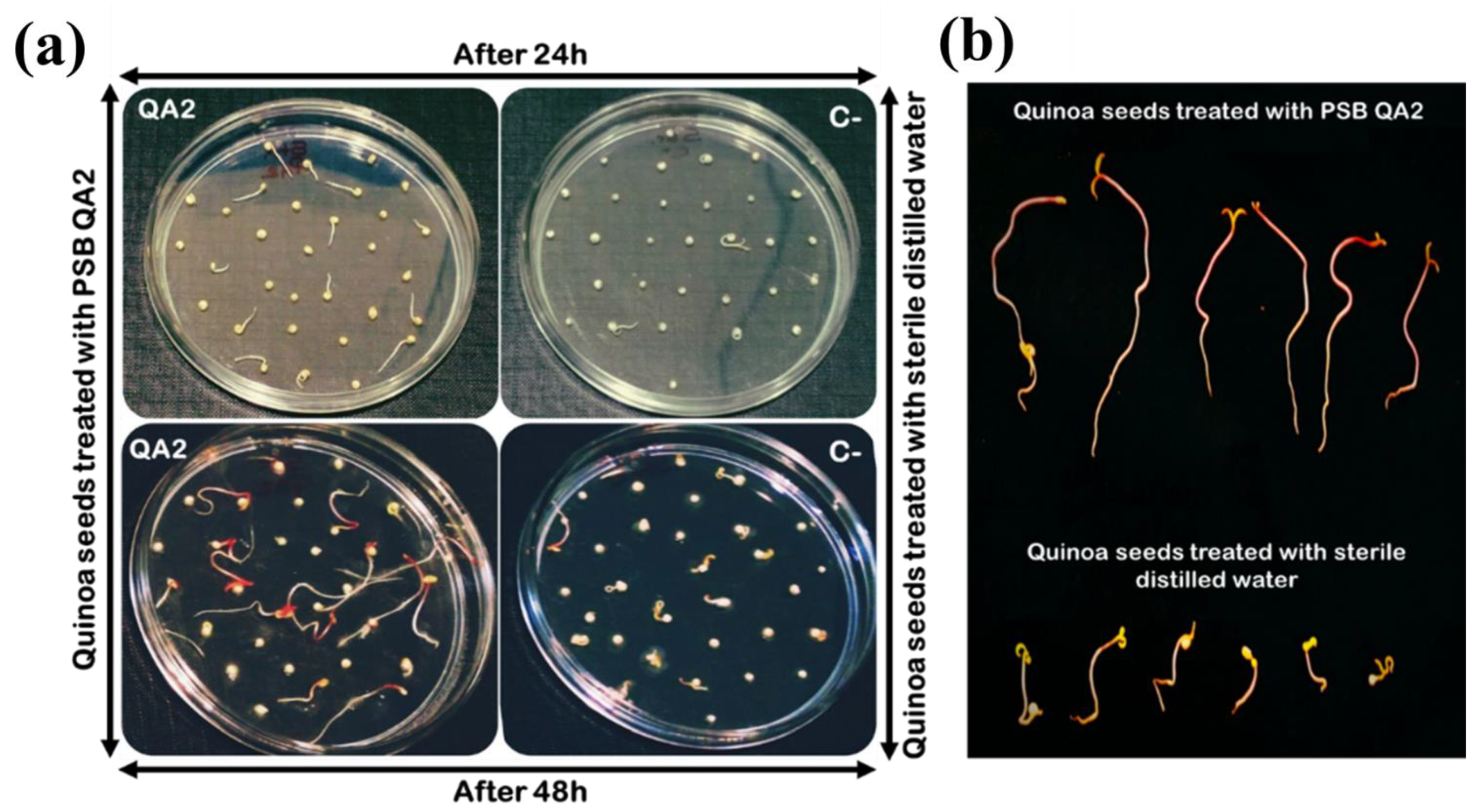
3.9. Strain B. velezensis QA2 Stimulates Quinoa Seed Germination In Vitro
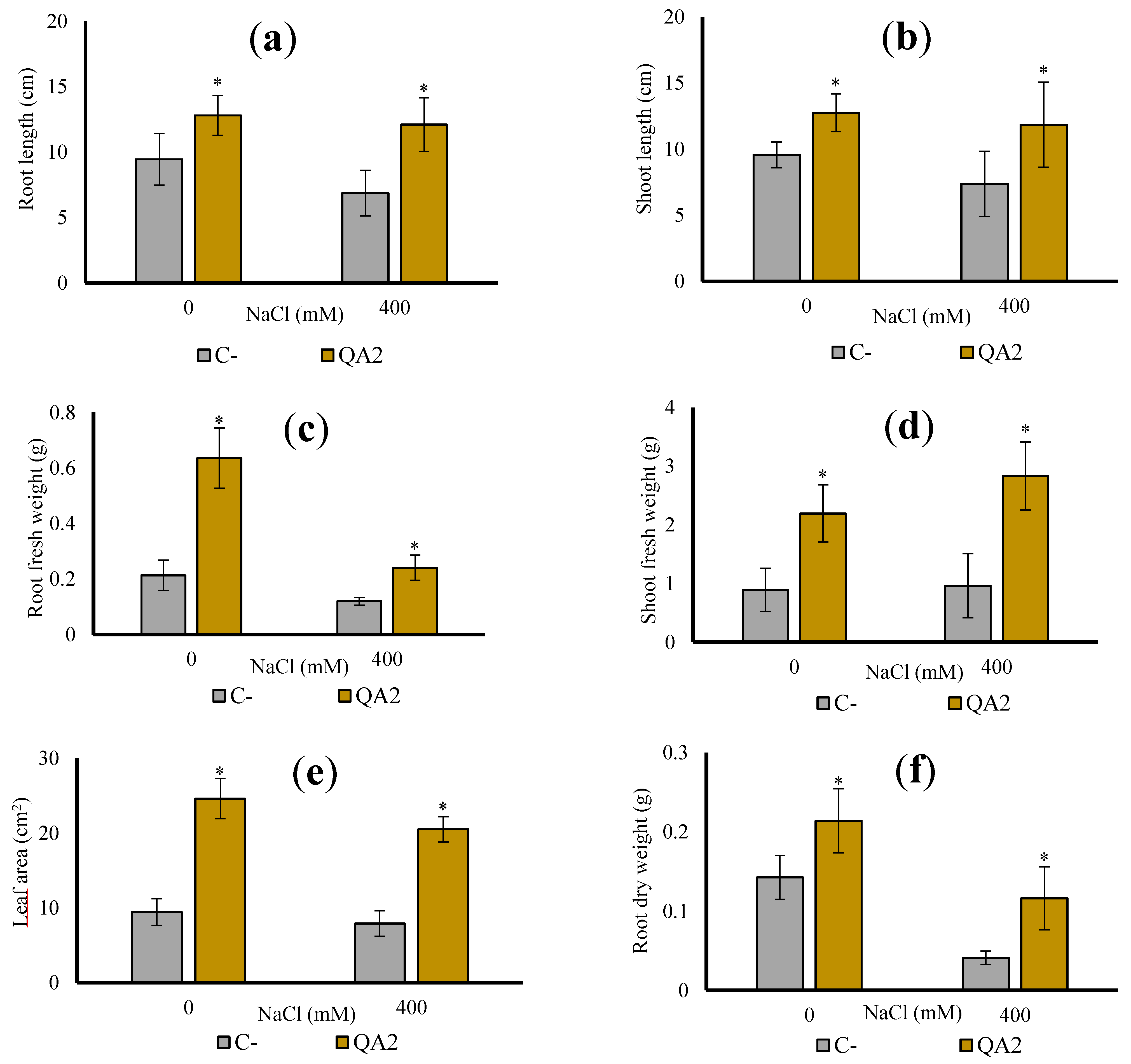
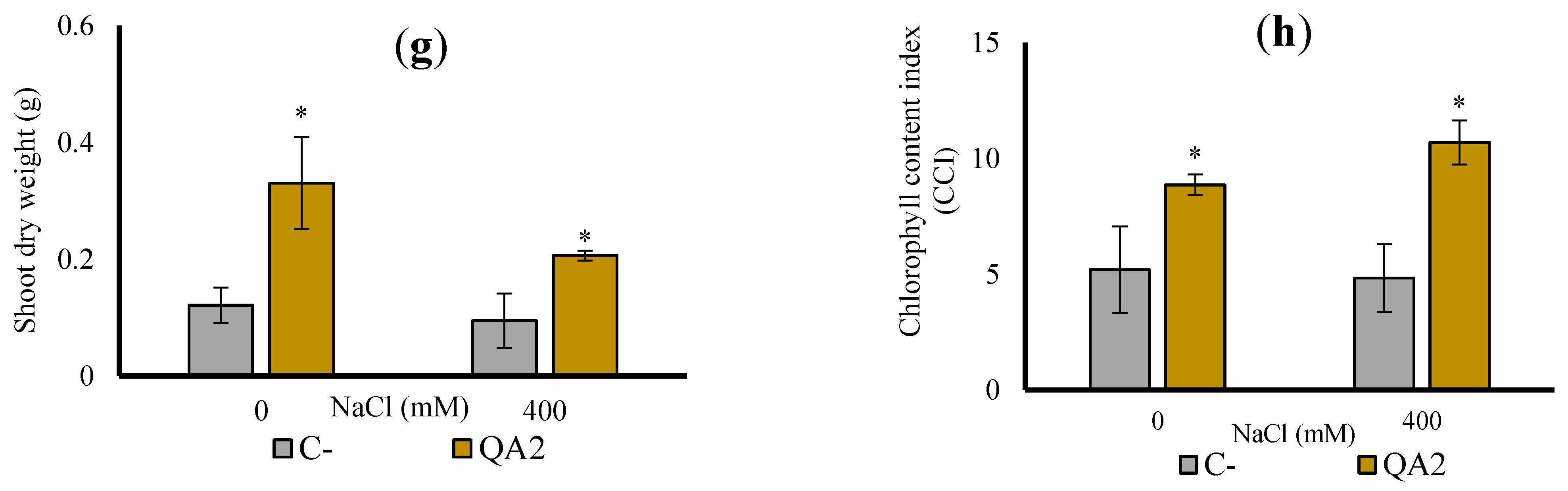
3.10. B. velezensis QA2 Strain Improves Shoot Biomass of Quinoa Plant Cultivated In Vivo under Salt Stress
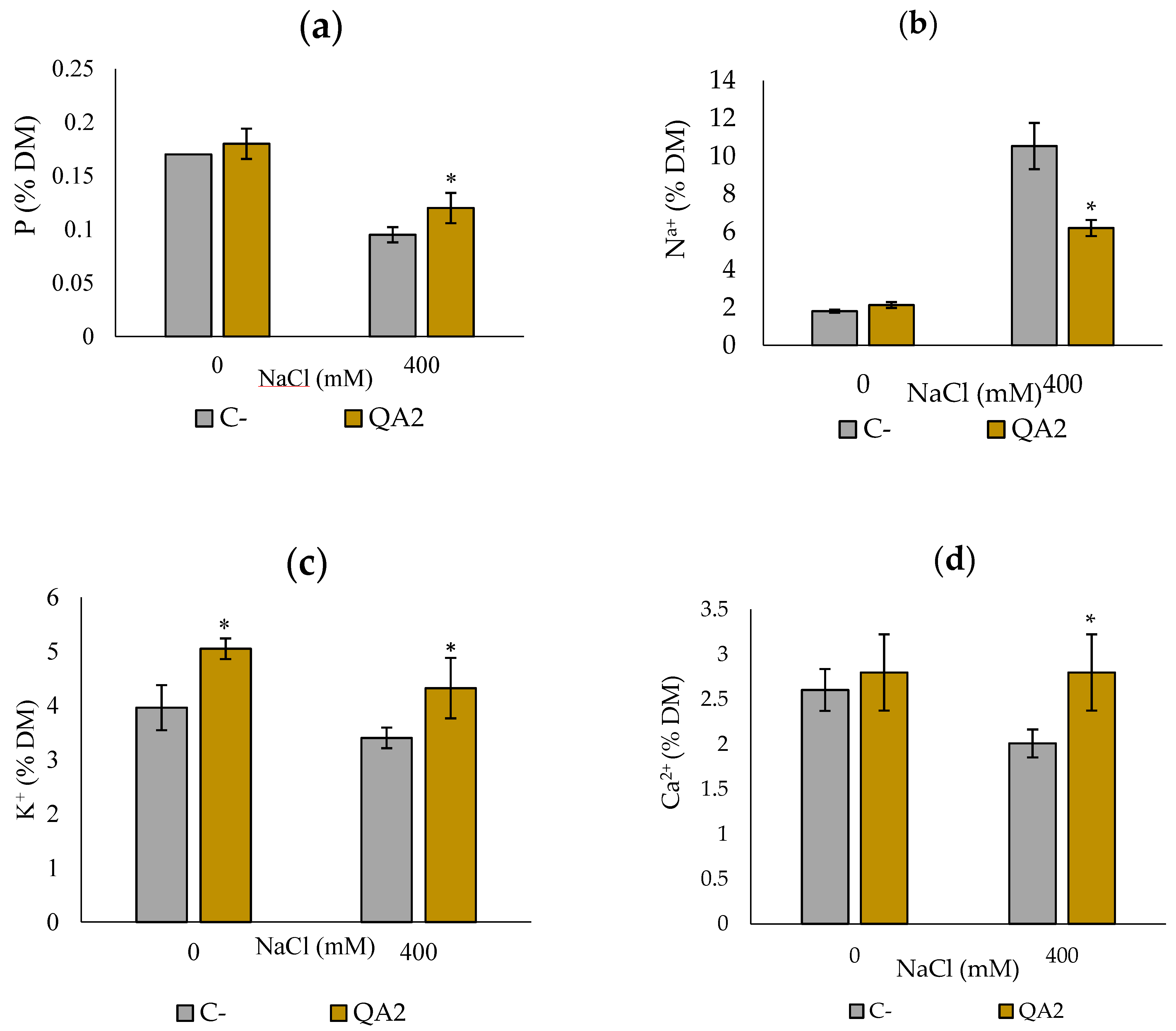
3.11. Analysis of Chlorophyll and P, Na+, K+ and Ca2+ Contents upon Plant Inoculation with B. velezensis QA2 Strain
4. Discussion
4.1. In Vitro and In Vivo Studies of the Bacillus velezensis QA2 Strain
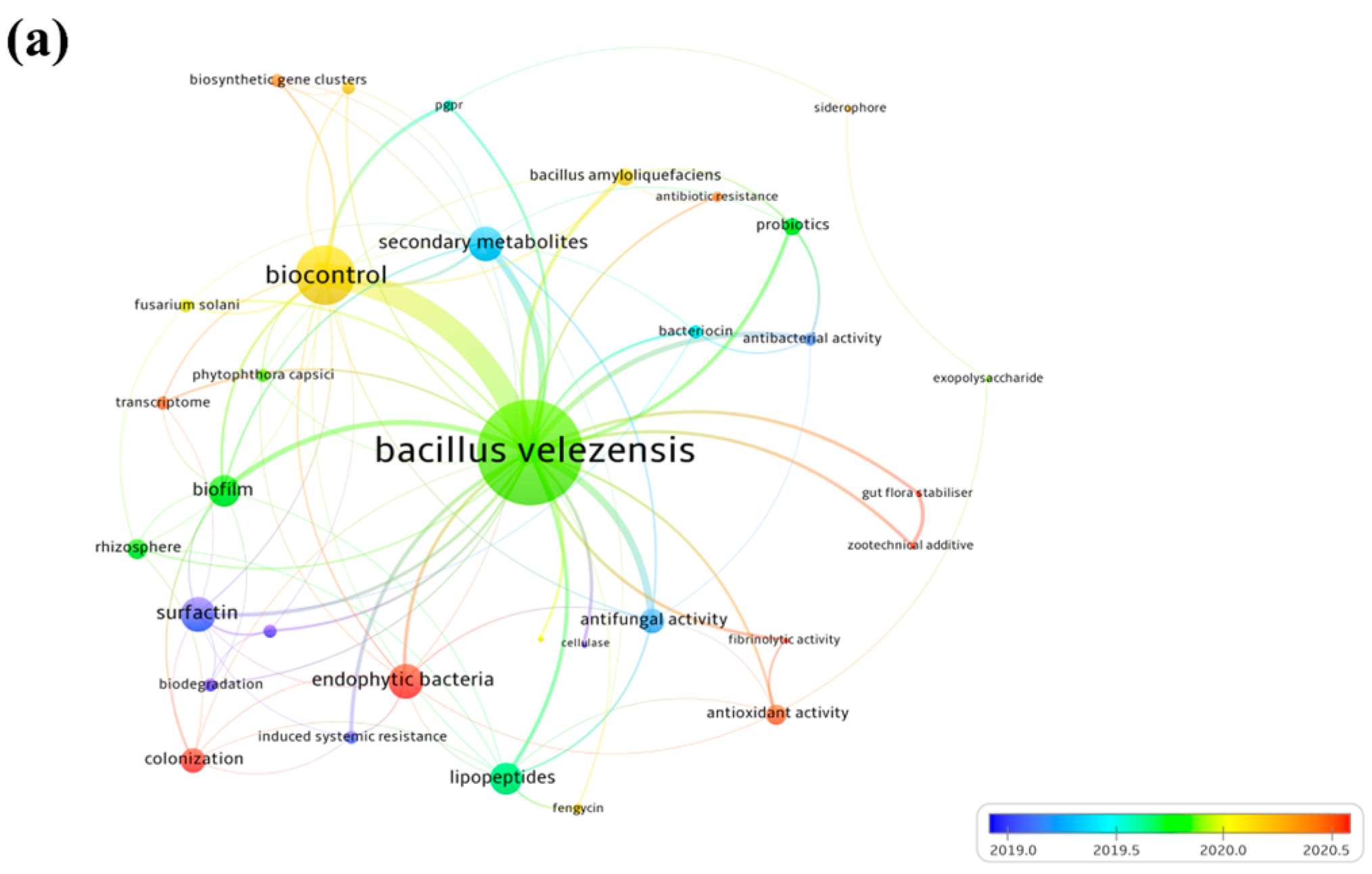
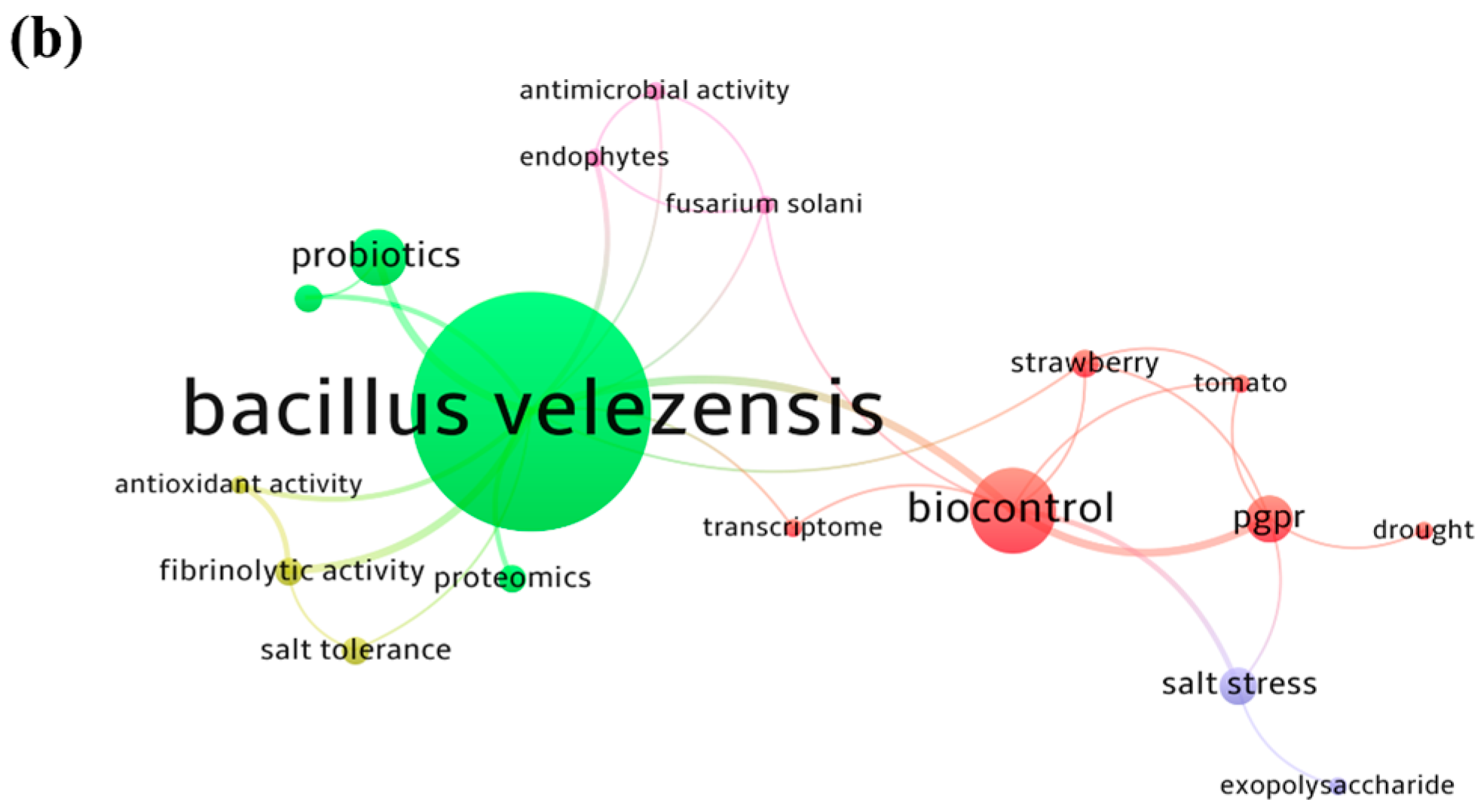
4.2. Hot Topics Analysis of Bacillus velezensis Strains
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 156, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, S.M.; Ahmad, M.; Zahir, Z.A.; Javaid, A.; Ashraf, M. The role of mycorrhizae and plant growth promoting rhizobacteria (PGPR) in improving crop productivity under stressful environments. Biotechnol. Adv. 2014, 32, 429–448. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, S.K.; Maurya, S.K.; Singh, D.P. Salinity tolerance in free living plant growth promoting rhizobacteria. Indian J. Sci. Res. 2012, 3, 73. [Google Scholar]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307. [Google Scholar] [CrossRef]
- Kilian, M.; Steiner, U.; Krebs, B.; Junge, H.; Schmiedeknecht, G.; Hain, R. FZB24® Bacillus subtilis–mode of action of a microbial agent enhancing plant vitality. Pflanzenschutz-Nachr. Bayer 2000, 1, 1. [Google Scholar]
- Brannen, P.; Kenney, D. Kodiak®—A successful biological-control product for suppression of soil-borne plant pathogens of cotton. J. Ind. Microbiol. Biotechnol. 1997, 19, 169–171. [Google Scholar] [CrossRef]
- Bisutti, I.; Pelz, J.; Büttner, C.; Stephan, D. Field assessment on the influence of RhizoVital® 42 fl. and Trichostar® on strawberries in the presence of soil-borne diseases. Crop Prot. 2017, 96, 195–203. [Google Scholar] [CrossRef]
- Ruan, C.-J.; da Silva, J.A.T.; Mopper, S.; Qin, P.; Lutts, S. Halophyte improvement for a salinized world. Crit. Rev. Plant Sci. 2010, 29, 329–359. [Google Scholar] [CrossRef]
- Flowers, T.J.; Galal, H.K.; Bromham, L. Evolution of halophytes: Multiple origins of salt tolerance in land plants. Funct. Plant Biol. 2010, 37, 604–612. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Gaurav, A.K.; Srivastava, S.; Verma, J.P. Plant Growth-Promoting Bacteria: Biological Tools for the Mitigation of Salinity Stress in Plants. Front. Microbiol. 2020, 11, 1216. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S. Learning from halophytes: Physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 2013, 112, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Numan, M.; Bashir, S.; Khan, Y.; Mumtaz, R.; Shinwari, Z.K.; Khan, A.L.; Khan, A.; Al-Harrasi, A. Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: A review. Microbiol. Res. 2018, 209, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Ruales, J.; Nair, B.M. Nutritional quality of the protein in quinoa (Chenopodium quinoa, Willd) seeds. Plant Foods Hum. Nutr. 1992, 42, 1–11. [Google Scholar] [CrossRef]
- Vega-Gálvez, A.; Miranda, M.; Vergara, J.; Uribe, E.; Puente, L.; Martínez, E.A. Nutrition facts and functional potential of quinoa (Chenopodium quinoa willd.), an ancient Andean grain: A review. J. Sci. Food Agric. 2010, 90, 2541–2547. [Google Scholar] [CrossRef]
- Adolf, V.I.; Jacobsen, S.-E.; Shabala, S. Salt tolerance mechanisms in quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Liu, F.-P.; Liu, H.-Q.; Zhou, H.-L.; Dong, Z.-G.; Bai, X.-H.; Bai, P.; Qiao, J.-J. Isolation and characterization of phosphate-solubilizing bacteria from betel nut (Areca catechu) and their effects on plant growth and phosphorus mobilization in tropical soils. Biol. Fertil. Soils 2014, 50, 927–937. [Google Scholar] [CrossRef]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2003, 118, 10–15. [Google Scholar] [CrossRef]
- Oteino, N.; Lally, R.D.; Kiwanuka, S.; Lloyd, A.; Ryan, D.; Germaine, K.J.; Dowling, D.N. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar] [CrossRef]
- Biswas, J.; Banerjee, A.; Rai, M.; Naidu, R.; Biswas, B.; Vithanage, M.; Chandra Dash, M.; Sarkar, S.; Meers, E. Potential application of selected metal resistant phosphate solubilizing bacteria isolated from the gut of earthworm (Metaphire posthuma) in plant growth promotion. Geoderma 2018, 330, 117–124. [Google Scholar] [CrossRef]
- Sarkar, A.; Ghosh, P.; Pramanik, K.; Mitra, S.; Soren, T.; Pandey, S.; Mondal, M.; Maiti, T. A halotolerant Enterobacter sp. displaying ACC deaminase activity promotes rice seedling growth under salt stress. Res. Microbiol. 2017, 169, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Mahdi, I.; Fahsi, N.; Hafidi, M.; Allaoui, A.; Biskri, L. Plant Growth Enhancement using Rhizospheric Halotolerant Phosphate Solubilizing Bacterium Bacillus licheniformis QA1 and Enterobacter asburiae QF11 Isolated from Chenopodium quinoa Willd. Microorganisms 2020, 8, 948. [Google Scholar] [CrossRef] [PubMed]
- Jetiyanon, K. Multiple mechanisms of Enterobacter asburiae strain RS83 for plant growth enhancement. Songklanakarin J. Sci. Technol. 2015, 37, 29–36. [Google Scholar]
- Habibi, S.; Djedidi, S.; Prongjunthuek, K.; Mortuza, M.F.; Ohkama-Ohtsu, N.; Sekimoto, H.; Yokoyoma, T. Physiological and genetic characterization of rice nitrogen fixer PGPR isolated from rhizosphere soils of different crops. Plant Soil 2014, 379, 51–66. [Google Scholar] [CrossRef]
- Li, H.; Ding, X.; Wang, C.; Ke, H.; Wu, Z.; WANG, Y.; Liu, H.; Guo, J. Control of tomato yellow leaf curl virus disease by Enterobacter asburiaeBQ9 as a result of priming plant resistance in tomatoes. Turk. J. Biol. 2016, 40, 150–159. [Google Scholar] [CrossRef]
- Patel, K.S.; Naik, J.H.; Chaudhari, S.; Amaresan, N. Characterization of culturable bacteria isolated from hot springs for plant growth promoting traits and effect on tomato (Lycopersicon esculentum) seedling. Comptes Rendus Biol. 2017, 340, 244–249. [Google Scholar] [CrossRef]
- Kumar Johri, J.; Surange, S.; Shekhar Nautiyal, C. Occurrence of Salt, pH, and Temperature-tolerant, Phosphate-solubilizing Bacteria in Alkaline Soils. Curr. Microbiol. 1999, 39, 89–93. [Google Scholar] [CrossRef]
- Nautiyal, C.S. An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef]
- Mukhtar, S.; Shahid, I.; Mehnaz, S.; Malik, K. Assessment of two carrier materials for phosphate solubilizing biofertilizers and their effect on growth of wheat (Triticum aestivum L.). Microbiol. Res. 2017, 205, 107–117. [Google Scholar] [CrossRef]
- Kryachko, Y.; Semler, D.; Vogrinetz, J.; Lemke, M.; Irvine, R.; Davidson, J.; Links, M.G.; McCarthy, E.L.; Haug, B.; Hemmingsen, S.M. Analyses of 16S rRNA and cpn60 gene sequences provide complementary information about potentially useful and harmful oil field microbiota. Int. Biodeterior. Biodegrad. 2017, 123, 320–327. [Google Scholar] [CrossRef]
- Hassan, A.K.; Shahata, M.A.; Refaie, E.M.; Ibrahim, R.S. Pathogenicity testing and antimicrobial susceptibility of Helicobacter pullorum isolates from chicken origin. Int. J. Vet. Sci. Med. 2014, 2, 72–77. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leveau, J.H.; Lindow, S.E. Utilization of the plant hormone indole-3-acetic acid for growth by Pseudomonas putida strain 1290. Appl. Environ. Microbiol. 2005, 71, 2365–2371. [Google Scholar] [CrossRef] [PubMed]
- Latif Khan, A.; Ahmed Halo, B.; Elyassi, A.; Ali, S.; Al-Hosni, K.; Hussain, J.; Al-Harrasi, A.; Lee, I.-J. Indole acetic acid and ACC deaminase from endophytic bacteria improves the growth of Solarium lycopersicum. Electron. J. Biotechnol. 2016, 21, 58–64. [Google Scholar] [CrossRef]
- Christensen, G.D.; Simpson, W.A.; Younger, J.; Baddour, L.; Barrett, F.; Melton, D.; Beachey, E. Adherence of coagulase-negative staphylococci to plastic tissue culture plates: A quantitative model for the adherence of staphylococci to medical devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Milagres, A.; Machuca, A.; Napoleão, D. Detection of siderophore production from several fungi and bacteria by a modification of chrome Azurol S (CAS) agar plate assay. J. Microbiol. Methods 1999, 37, 111–122. [Google Scholar] [CrossRef]
- Cappuccino, J.C.; Natalie, S. Microbiology: A Laboratory Manual, 3rd ed.; Co, B.C.P., Ed.; Benjamin/Cummings Publishing Company: Redwood City, CA, USA, 1992. [Google Scholar]
- Chrouqi, L.; Lahcen, O.; Jadrane, I.; Koussa, T.; Alfeddy, M.N. Screening of soil rhizobacteria isolated from wheat plants grown in the Marrakech region (Morocco, North Africa) for plant growth promoting activities. JMES 2017, 8, 3382–3390. [Google Scholar]
- Khanghahi, M.Y.; Ricciuti, P.; Allegretta, I.; Terzano, R.; Crecchio, C. Solubilization of insoluble zinc compounds by zinc solubilizing bacteria (ZSB) and optimization of their growth conditions. Environ. Sci. Pollut. Res. 2018, 25, 25862–25868. [Google Scholar] [CrossRef]
- Lorck, H. Production of hydrocyanic acid by bacteria. Physiol. Plant. 1948, 1, 142–146. [Google Scholar] [CrossRef]
- Smibert, R. Phenotypic characterization. In Methods for General and Molecular Microbiology; American Society for Microbiology: Washington, DC, USA, 1994; pp. 607–654. [Google Scholar]
- Kasana, R.C.; Salwan, R.; Dhar, H.; Dutt, S.; Gulati, A. A rapid and easy method for the detection of microbial cellulases on agar plates using gram’s iodine. Curr. Microbiol. 2008, 57, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Navarro, V.; Villarreal, M.L.; Rojas, G.; Lozoya, X. Antimicrobial evaluation of some plants used in Mexican traditional medicine for the treatment of infectious diseases. J. Ethnopharmacol. 1996, 53, 143–147. [Google Scholar] [CrossRef]
- Costa, A.C.B.P.; de Campos Rasteiro, V.M.; Pereira, C.A.; da Silva Hashimoto, E.S.H.; Beltrame, M.; Junqueira, J.C.; Jorge, A.O.C. Susceptibility of Candida albicans and Candida dubliniensis to erythrosine- and LED-mediated photodynamic therapy. Arch. Oral Biol. 2011, 56, 1299–1305. [Google Scholar] [CrossRef] [Green Version]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Ji, S.H.; Gururani, M.A.; Chun, S.C. Isolation and characterization of plant growth promoting endophytic diazotrophic bacteria from Korean rice cultivars. Microbiol. Res. 2014, 169, 83–98. [Google Scholar] [CrossRef]
- Suleman, M.; Yasmin, S.; Rasul, M.; Yahya, M.; Atta, B.M.; Mirza, M.S. Phosphate solubilizing bacteria with glucose dehydrogenase gene for phosphorus uptake and beneficial effects on wheat. PLoS ONE 2018, 13, e0204408. [Google Scholar] [CrossRef]
- Pitzschke, A. Developmental peculiarities and seed-borne endophytes in quinoa: Omnipresent, robust bacilli contribute to plant fitness. Front. Microbiol. 2016, 7, 2. [Google Scholar] [CrossRef]
- Islam, S.; Mannan Akanda, A.; Prova, A.; Islam, T.; Hossain, M. Isolation and Identification of Plant Growth Promoting Rhizobacteria from Cucumber Rhizosphere and Their Effect on Plant Growth Promotion and Disease Suppression. Front. Microbiol. 2016, 6, 1360. [Google Scholar] [CrossRef]
- Khan, N.; Zandi, P.; Ali, S.; Mehmood, A.; Shahid, M.A.; Yang, J. Corrigendum: Impact of Salicylic acid and PGPR on the Drought Tolerance and Phytoremediation potential of Helianthus annus. Front. Microbiol. 2019, 10, 2222. [Google Scholar] [CrossRef]
- Shah, S.H.H.; Lei, S.; Ali, M.; Doronin, D.; Hussain, S.T. Prosumption: Bibliometric analysis using HistCite and VOSviewer. Kybernetes 2020, 49, 1020–1045. [Google Scholar] [CrossRef]
- Mann, E.E.; Wozniak, D.J. Pseudomonas biofilm matrix composition and niche biology. FEMS Microbiol. Rev. 2012, 36, 893–916. [Google Scholar] [CrossRef]
- Rijavec, T.; Lapanje, A. Hydrogen Cyanide in the Rhizosphere: Not Suppressing Plant Pathogens, but Rather Regulating Availability of Phosphate. Front. Microbiol. 2016, 7, 1785. [Google Scholar] [CrossRef]
- Mitchell, R.; Alexander, M. Lysis of soil fungi by bacteria. Can. J. Microbiol. 2011, 9, 169–177. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial Phosphorus Solubilization and Its Potential for Use in Sustainable Agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.R.; Reva, O.; et al. Comparative analysis of the complete genome sequence of the plant growth–promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, C.A.; Kim, S.-J.; Kwon, S.-W.; Rooney, A.P. Bacillus velezensis is not a later heterotypic synonym of Bacillus amyloliquefaciens; Bacillus methylotrophicus, Bacillus amyloliquefaciens subsp. plantarum and ‘Bacillus oryzicola’ are later heterotypic synonyms of Bacillus velezensis based on phylogenomics. Int. J. Syst. Evol. Microbiol. 2016, 66, 1212–1217. [Google Scholar] [CrossRef]
- Dinesh, R.; Anandaraj, M.; Kumar, A.; Bini, Y.K.; Subila, K.P.; Aravind, R. Isolation, characterization, and evaluation of multi-trait plant growth promoting rhizobacteria for their growth promoting and disease suppressing effects on ginger. Microbiol. Res. 2015, 173, 34–43. [Google Scholar] [CrossRef]
- Went, F.W.; Thimann, K.V. Phytohormones; The Macmillan Company: New York, NY, USA, 1937. [Google Scholar]
- Chaiharn, M.; Lumyong, S. Screening and optimization of indole-3-acetic acid production and phosphate solubilization from rhizobacteria aimed at improving plant growth. Curr. Microbiol. 2011, 62, 173–181. [Google Scholar] [CrossRef]
- Arshad, M.; Frankenberger, W. Microbial production of plant grow the regulators. Blaine Metting 1992, 307–347. [Google Scholar]
- Khan, M.S.; Gao, J.; Chen, X.; Zhang, M.; Yang, F.; Du, Y.; Zhang, X. The Endophytic Bacteria Bacillus velezensisLle-9, Isolated from Lilium leucanthum, Harbors Antifungal Activity and Plant Growth-Promoting Effects. J. Microbiol. Biotechnol. 2020, 30, 668–680. [Google Scholar] [CrossRef]
- Tailor, A.J.; Joshi, B.H. Characterization and optimization of siderophore production from Pseudomonas fluorescens strain isolated from sugarcane rhizosphere. J. Environ. Res. Dev. 2012, 6, 688–694. [Google Scholar]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef]
- Gontia-Mishra, I.; Sapre, S.; Sharma, A.; Tiwari, S. Alleviation of mercury toxicity in wheat by the interaction of mercury-tolerant plant growth-promoting rhizobacteria. J. Plant Growth Regul. 2016, 35, 1000–1012. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springerplus 2013, 2, 587. [Google Scholar] [CrossRef]
- Bensidhoum, L.; Nabti, E.; Tabli, N.; Kupferschmied, P.; Weiss, A.; Rothballer, M.; Schmid, M.; Keel, C.; Hartmann, A. Heavy metal tolerant Pseudomonas protegens isolates from agricultural well water in northeastern Algeria with plant growth promoting, insecticidal and antifungal activities. Eur. J. Soil Biol. 2016, 75, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Walsh, U.F.; Morrissey, J.P.; O’Gara, F. Pseudomonas for biocontrol of phytopathogens: From functional genomics to commercial exploitation. Curr. Opin. Biotechnol. 2001, 12, 289–295. [Google Scholar] [CrossRef]
- Agbodjato, N.A.; Noumavo, P.A.; Baba-Moussa, F.; Salami, H.A.; Sina, H.; Sèzan, A.; Bankolé, H.; Adjanohoun, A.; Baba-Moussa, L. Characterization of potential plant growth promoting rhizobacteria isolated from Maize (Zea mays L.) in central and Northern Benin (West Africa). Appl. Environ. Soil Sci. 2015, 2015, 901656. [Google Scholar] [CrossRef]
- Orhan, F. Alleviation of salt stress by halotolerant and halophilic plant growth-promoting bacteria in wheat (Triticum aestivum). Braz. J. Microbiol. 2016, 47, 621–627. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Khatoon, Z.; McTiernan, C.; Suuronen, E.; Mah, T.-F.; Alarcon, E. Bacterial biofilm formation on implantable devices and approaches to its treatment and prevention. Heliyon 2018, 4, e01067. [Google Scholar] [CrossRef]
- Bleich, R.; Watrous, J.D.; Dorrestein, P.C.; Bowers, A.A.; Shank, E.A. Thiopeptide antibiotics stimulate biofilm formation in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2015, 112, 3086–3091. [Google Scholar] [CrossRef]
- Zhao, L.; Xu, Y.; Lai, X.H.; Shan, C.; Deng, Z.; Ji, Y. Screening and characterization of endophytic Bacillus and Paenibacillus strains from medicinal plant Lonicera japonica for use as potential plant growth promoters. Braz. J. Microbiol. 2015, 46, 977–989. [Google Scholar] [CrossRef]
- Amaresan, N.; Kumar, K.; Madhuri, K.; Usharani, G.K. Isolation and characterization of salt tolerant plant growth promoting rhizobacteria from plants grown in tsunami affected regions of Andaman and Nicobar Islands. Geomicrobiol. J. 2016, 33, 942–947. [Google Scholar] [CrossRef]
- Xun, F.; Xie, B.; Liu, S.; Guo, C. Effect of plant growth-promoting bacteria (PGPR) and arbuscular mycorrhizal fungi (AMF) inoculation on oats in saline-alkali soil contaminated by petroleum to enhance phytoremediation. Environ. Sci. Pollut. Res. Int. 2015, 22, 598–608. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Uhl, J.; Grosch, R.; Alquéres, S.; Pittroff, S.; Dietel, K.; Schmitt-Kopplin, P.; Borriss, R.; Hartmann, A. Cyclic Lipopeptides of Bacillus amyloliquefaciens subsp. plantarum Colonizing the Lettuce Rhizosphere Enhance Plant Defense Responses Toward the Bottom Rot Pathogen Rhizoctonia solani. Mol. Plant-Microbe Interact. 2015, 28, 984–995. [Google Scholar] [CrossRef] [Green Version]
- Burkett-Cadena, M.; Kokalis-Burelle, N.; Lawrence, K.S.; van Santen, E.; Kloepper, J.W. Suppressiveness of root-knot nematodes mediated by rhizobacteria. Biol. Control 2008, 47, 55–59. [Google Scholar] [CrossRef]
- Cao, Y.; Pi, H.; Chandrangsu, P.; Li, Y.; Wang, Y.; Zhou, H.; Xiong, H.; Helmann, J.D.; Cai, Y. Antagonism of Two Plant-Growth Promoting Bacillus velezensis Isolates Against Ralstonia solanacearum and Fusarium oxysporum. Sci. Rep. 2018, 8, 4360. [Google Scholar] [CrossRef]
- Borriss, R. Use of Plant-Associated Bacillus Strains as Biofertilizers and Biocontrol Agents in Agriculture. In Bacteria in Agrobiology: Plant Growth Responses; Springer: Berlin/Heidelberg, Germany, 2011; pp. 41–76. [Google Scholar]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef]
- Alvarez, F.; Castro, M.; Principe, A.; Borioli, G.; Fischer, S.; Mori, G.; Jofre, E. The plant-associated Bacillus amyloliquefaciens strains MEP2 18 and ARP2 3 capable of producing the cyclic lipopeptides iturin or surfactin and fengycin are effective in biocontrol of sclerotinia stem rot disease. J. Appl. Microbiol. 2012, 112, 159–174. [Google Scholar] [CrossRef]
- Rabbee, M.F.; Ali, M.S.; Choi, J.; Hwang, B.S.; Jeong, S.C.; Baek, K.-H. Bacillus velezensis: A Valuable Member of Bioactive Molecules within Plant Microbiomes. Molecules 2019, 24, 1046. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalek, A.; Behiry, S.I.; Al-Askar, A.A. Bacillus velezensis PEA1 Inhibits Fusarium oxysporum Growth and Induces Systemic Resistance to Cucumber Mosaic Virus. Agronomy 2020, 10, 1312. [Google Scholar] [CrossRef]
- Bakhshandeh, E.; Rahimian, H.; Pirdashti, H.; Nematzadeh, G.A. Phosphate solubilization potential and modeling of stress tolerance of rhizobacteria from rice paddy soil in northern Iran. World J. Microbiol. Biotechnol. 2014, 30, 2437–2447. [Google Scholar] [CrossRef]
- Mansour, M. Nitrogen containing compounds and adaptation of plants to salinity stress. Biol. Plant. 2000, 43, 491–500. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Milošević, N.; Marinković, J.; Tintor, B. Mitigating abiotic stress in crop plants by microorganisms. Matica Srp. Proc. Nat. Sci. 2012, 123, 17–26. [Google Scholar] [CrossRef]
- Lai, W.-A.; Rekha, P.; Arun, A.; Young, C.-C. Effect of mineral fertilizer, pig manure, and Azospirillum rugosum on growth and nutrient contents of Lactuca sativa L. Biol. Fertil. Soils 2008, 45, 155–164. [Google Scholar] [CrossRef]
- Tahir, M.; Mirza, M.S.; Zaheer, A.; Dimitrov, M.R.; Smidt, H.; Hameed, S. Isolation and identification of phosphate solubilizer Azospirillum, Bacillus and Enterobacter strains by 16SrRNA sequence analysis and their effect on growth of wheat (Triticum aestivum L.). Aust. J. Crop Sci. 2013, 7, 1284–1292. [Google Scholar]
- Masmoudi, F.; Tounsi, S.; Dunlap, C.A.; Trigui, M. Endophytic halotolerant Bacillus velezensis FMH2 alleviates salt stress on tomato plants by improving plant growth and altering physiological and antioxidant responses. Plant Physiol. Biochem. 2021, 165, 217–227. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Wu, G.; Veronican Njeri, K.; Shen, Q.; Zhang, N.; Zhang, R. Induced maize salt tolerance by rhizosphere inoculation of Bacillus amyloliquefaciens SQR9. Physiol. Plant. 2016, 158, 34–44. [Google Scholar] [CrossRef]
- Hegazi, A.M.; El-Shraiy, A.M.; Ghoname, A.A. Mitigation of Salt Stress Negative Effects on Sweet Pepper Using Arbuscular Mycorrhizal Fungi (AMF), Bacillus megaterium and Brassinosteroids (BRs). Gesunde Pflanz. 2017, 69, 91–102. [Google Scholar] [CrossRef]
- Santos, A.D.A.; Silveira, J.A.G.D.; Bonifacio, A.; Rodrigues, A.C.; Figueiredo, M.D.V.B. Antioxidant response of cowpea co-inoculated with plant growth-promoting bacteria under salt stress. Braz. J. Microbiol. 2018, 49, 513–521. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 0 mM NaCl | 400 mM NaCl | ||
|---|---|---|---|
| Symbol | Treatment | Symbol | Treatment |
| C− | Seeds treated with sterilized distilled water (Negative control) | C− | Seeds treated with sterilized distilled water (Saline control) |
| QA2 | Seeds treated with QA2 isolate | QA2 | Seeds treated with QA2 isolate |
| Assay | Method | Result |
|---|---|---|
| Bacterial isolate | QA2 | |
| Morphology | Culture on TSA plate at 30 °C | Motile |
| Velvety | ||
| Rod-shaped | ||
| Extreme growth properties | NaCl tolerance (%) | 11% |
| Maximum tolerable temperature | 45 °C | |
| Siderophore production | CAS medium | +++ |
| HCN production | TSA medium amended with 0.44% glycine | +++ |
| Extracellular enzymes (Halo diameter/colony diameter) | Protease | 1.15 |
| Cellulase | 5.41 | |
| Zinc solubilization | ZnO | ++ |
| Zn3(PO4)2 | − | |
| ZnCO3 | − | |
| Antibiotic resistance using antibiotic-amended agar | Ampicillin (100 µg/mL) | − |
| Chloramphenicol (20 µg/mL) | − | |
| Streptomycin (100 µg/mL) | − | |
| Spectinomycin (60 µg/mL) | − | |
| Kanamycin (50 µg/mL) | − | |
| Tetracycline (10 µg/mL) | − | |
| Antibacterial activity using disk diffusion method | Escherichia coli | +++ |
| Salmonella typhi | +++ | |
| Enterococcus faecalis | ++ | |
| Staphylococcus aureus | +++ | |
| Antifungal activity using bacteria—fungi confrontation assay | Fusarium oxysporum f.sp. albedinisCandida albicans | +++++ |
| Germination Parameters | C− | B. velezensis QA2 | |
|---|---|---|---|
| Germination rate (%) | 24 h | 16.6 ± 6.65 | 57.3 ± 10.26 * |
| 48 h | 58.6 ± 5.13 | 89.66 ± 6.5 * | |
| Total Length (cm) | 1.7 ± 0.55 | 6.48 ± 0.87 * | |
| Fresh Weight (mg) | 36 ± 10.5 | 71.6 ± 6.11 * | |
| Dry Weight (mg) | 6.6 ± 1.52 | 12 ± 2.64 * | |
| Seedling Vigor Index | 99.46 ± 56.95 | 580.60 ± 79.06 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahdi, I.; Allaoui, A.; Fahsi, N.; Biskri, L. Bacillus velezensis QA2 Potentially Induced Salt Stress Tolerance and Enhanced Phosphate Uptake in Quinoa Plants. Microorganisms 2022, 10, 1836. https://doi.org/10.3390/microorganisms10091836
Mahdi I, Allaoui A, Fahsi N, Biskri L. Bacillus velezensis QA2 Potentially Induced Salt Stress Tolerance and Enhanced Phosphate Uptake in Quinoa Plants. Microorganisms. 2022; 10(9):1836. https://doi.org/10.3390/microorganisms10091836
Chicago/Turabian StyleMahdi, Ismail, Abdelmounaaim Allaoui, Nidal Fahsi, and Latefa Biskri. 2022. "Bacillus velezensis QA2 Potentially Induced Salt Stress Tolerance and Enhanced Phosphate Uptake in Quinoa Plants" Microorganisms 10, no. 9: 1836. https://doi.org/10.3390/microorganisms10091836
APA StyleMahdi, I., Allaoui, A., Fahsi, N., & Biskri, L. (2022). Bacillus velezensis QA2 Potentially Induced Salt Stress Tolerance and Enhanced Phosphate Uptake in Quinoa Plants. Microorganisms, 10(9), 1836. https://doi.org/10.3390/microorganisms10091836