

Enterococcus faecalis NADH Peroxidase-Defective Mutants Stain Falsely in Colony Zymogram Assay for Extracellular Electron Transfer to Ferric Ions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Media
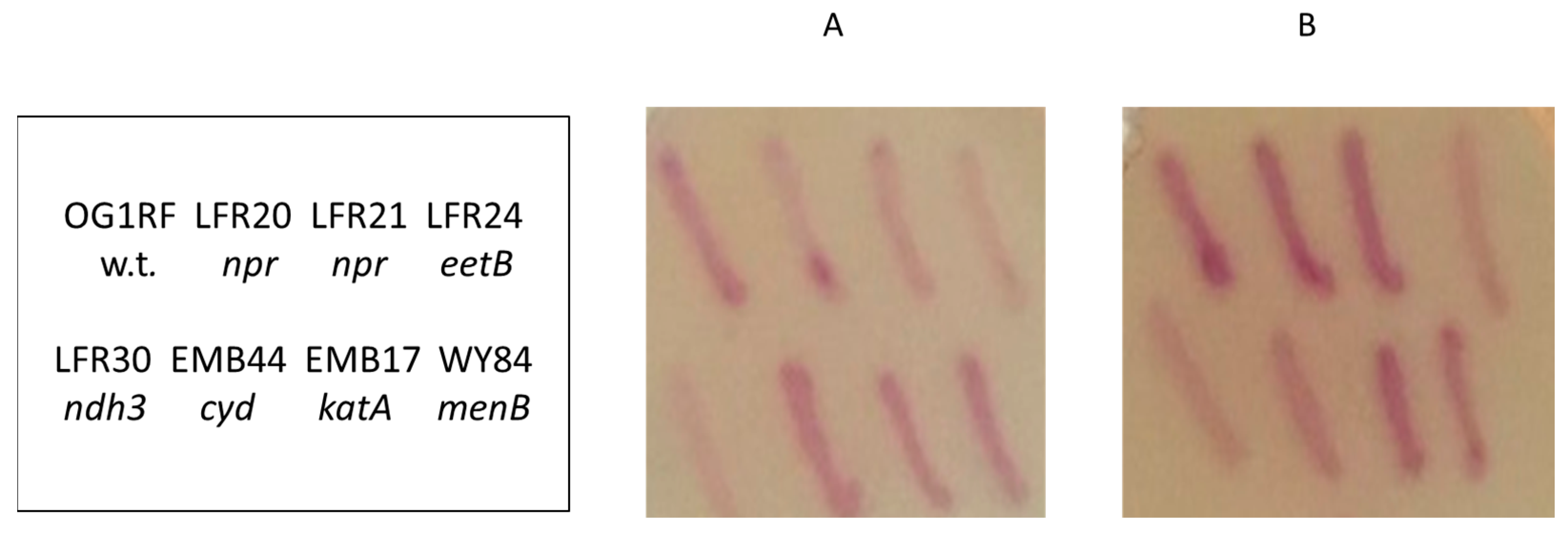
2.2. Colony Zymogram Staining for Ferric Reductase Activity
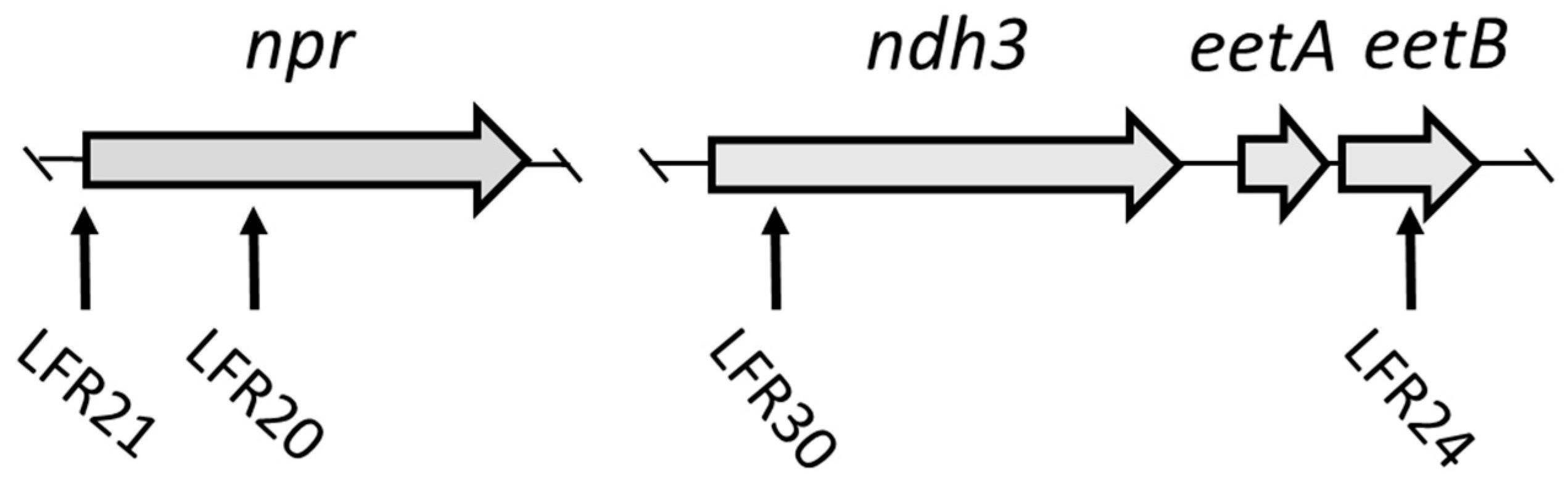
2.3. Genetic Characterization of Mutants
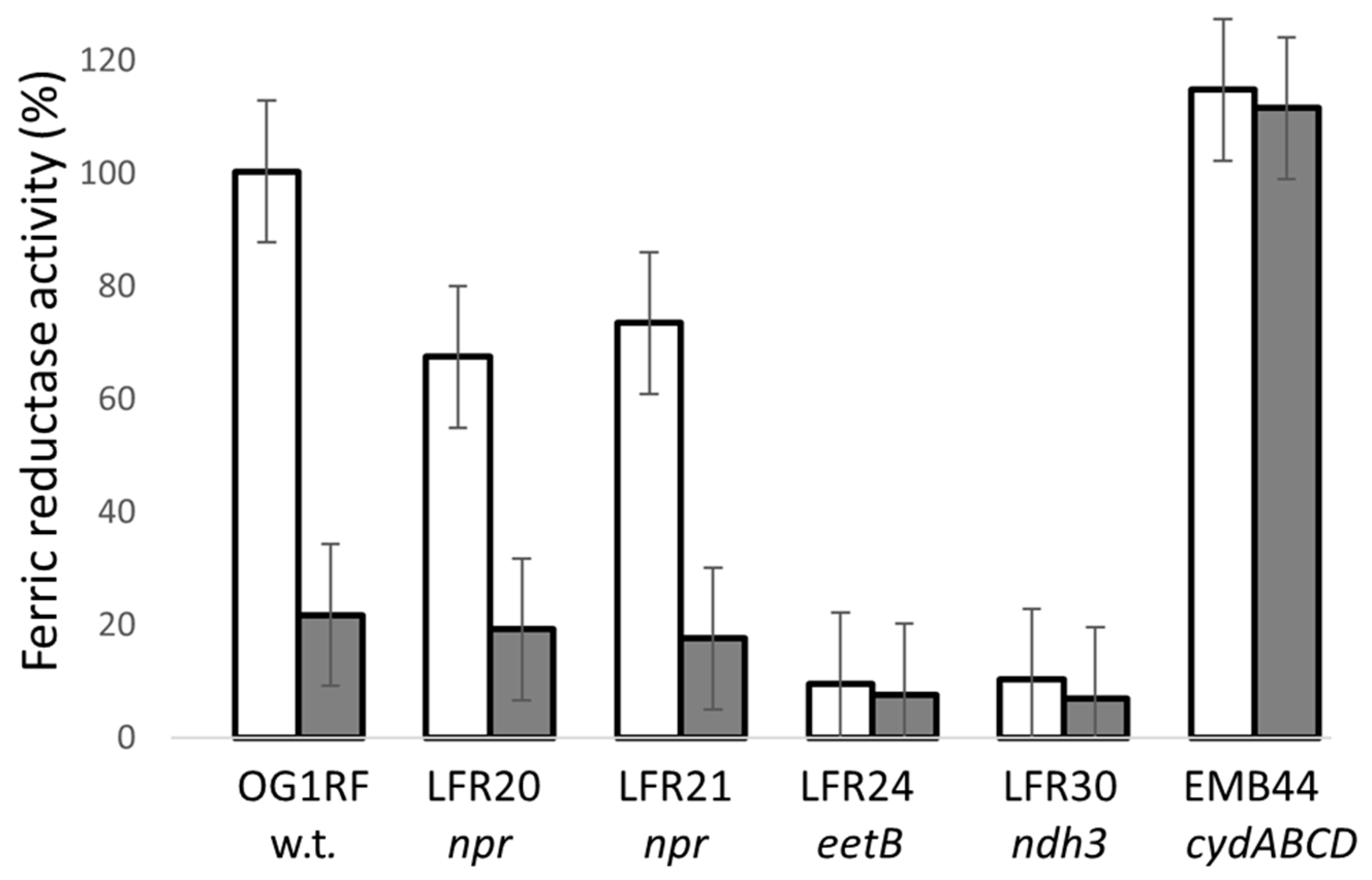
2.4. Determination of Ferric Reductase Activity of Washed Cells
3. Results
3.1. Screen for Mutants
3.2. npr-Defective Mutants
3.3. LFR24 and LFR30 Are Defective in EET to Ferric Ions
3.4. NADH Peroxidase Mutants Have Ferric Reductase Activity
3.5. Ferrozine Zymogram-Staining Activity of Colonies Depends on Multiple Factors
3.6. Hydrogen Peroxide Quenches in Ferric Reductase Assays
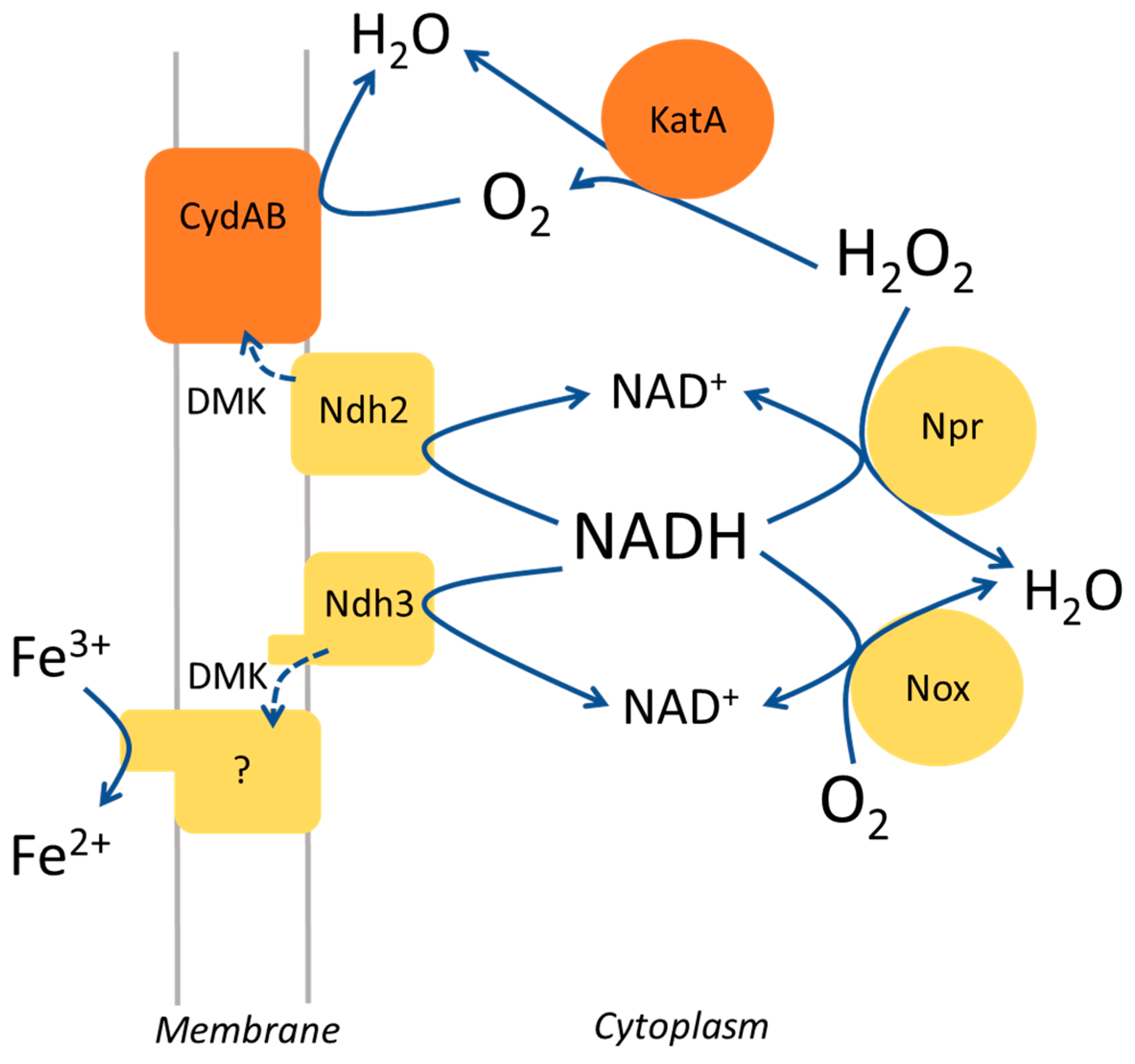
4. Discussion
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Selleck, E.M.; Van Tyne, D.; Gilmore, M.S. Pathogenicity of Enterococci. Microbiol. Spectr. 2019, 7, 1–38. [Google Scholar] [CrossRef]
- Ramsey, M.; Hartke, A.; Huycke, M. The physiology and metabolism of Enterococci. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, C.D., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Winstedt, L.; Frankenberg, L.; Hederstedt, L.; von Wachenfeldt, C. Enterococcus faecalis V583 contains a cytochrome bd-type respiratory oxidase. J. Bacteriol. 2000, 182, 3863–3866. [Google Scholar] [CrossRef] [Green Version]
- Baureder, M.; Hederstedt, L. Heme proteins in lactic acid bacteria. Adv. Microb. Physiol. 2013, 62, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.T.; Hyun, M.S.; Chang, I.S.; Kim, H.J.; Park, H.S.; Kim, B.H.; Kim, S.D.; Wimpenny, J.W.T.; Weightman, A.J. Dissimilatory Fe(III) reduction by an electrochmeically active lactic acid bacterium phylogentically related to Enterococcus gallinarium isolated from submerged soil. J. Appl. Microbiol. 2005, 99, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Pankratova, G.; Hasan, K.; Leech, D.; Hederstedt, L.; Gorton, L. Electrochemical wiring of the gram-positive bacterium Enterococcus faecalis with osmium redox polymer modified electrodes. Electrochem. Commun. 2017, 75, 56–59. [Google Scholar] [CrossRef]
- Pankratova, G.; Leech, D.; Gorton, L.; Hederstedt, L. Extracellular electron transfer by the gram-positive bacterium Enterococcus faecalis. Biochemistry 2018, 57, 4597–4603. [Google Scholar] [CrossRef] [Green Version]
- Keogh, D.; Lam, L.N.; Doyle, L.E.; Matysik, A.; Pavagadhi, S.; Umashankar, S.; Low, P.M.; Dale, J.L.; Song, Y.; Ng, S.P.; et al. Extracellular electron transfer powers Enterococcus faecalis biofilm metabolism. mBio 2018, 9, e00626-17. [Google Scholar] [CrossRef] [Green Version]
- Lovley, D. Electrotrophy: Other microbial species, iron, and electrodes as electron donors for microbial respirations. Bioresour. Technol. 2022, 345, 126553. [Google Scholar] [CrossRef]
- Lovley, D.; Holmes, D.E. Electromicrobiology: The ecophysiology of phylogenetically diverse electroactive microorganisms. Nat. Rev. Microbiol. 2022, 20, 5–19. [Google Scholar] [CrossRef]
- Aiyer, K.; Doyle, L.E. Capturing the signal of weak electricigens: A worthy endeavour. Trends Biotechnol. 2022, 40, 564–575. [Google Scholar] [CrossRef]
- Gomaa, O.M.; Costa, N.L.; Paquete, C.M. Electron transfer in Gram-positive bacteria: Enhancement strategies for bioelectrochemical applications. World J. Microbiol. Biotechnol. 2022, 38, 83. [Google Scholar] [CrossRef] [PubMed]
- Pankratova, G.; Hederstedt, L.; Gorton, L. Extracellular electron transfer features of Gram-positive bacteria. Anal. Chim. Acta 2019, 1076, 32–47. [Google Scholar] [CrossRef] [PubMed]
- Hederstedt, L.; Gorton, L.; Pankratova, G. Two routes for extracellular electron transfer in Enterococcus faecalis. J. Bacteriol. 2020, 202, e00725-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stookey, L.L. Ferrozine: A new spectrophotometric reagent for iron. Anal. Chem. 1970, 42, 779–781. [Google Scholar] [CrossRef] [Green Version]
- Baureder, M.; Hederstedt, L. Genes important for catalase activity in Enterococcus faecalis. PLoS ONE 2012, 7, e36725. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Yang, Y.; Moore, D.; Nimmo, S.; Lightfoot, S.; Huycke, M. 4-Hydroxy-2-nonenal mediates genotoxicity and bystander effects caused by Enterococcus faecalis infected macrophages. Gastroenterology 2012, 142, 543–551. [Google Scholar] [CrossRef] [Green Version]
- Dunny, G.; Brown, B.; Clewell, D. Induced cell aggregation and mating in Streptococcus faecalis: Evidence for a bacterial sex pheromone. Proc. Natl. Avad. Sci. USA 1978, 75, 3479–3483. [Google Scholar] [CrossRef] [Green Version]
- Yagi, Y.; Clewell, D.B. Recombinant-deficient mutants of Streptococcus faecalis. J. Bacteriol. 1980, 143, 966–970. [Google Scholar] [CrossRef] [Green Version]
- Frankenberg, L.; Brugna, M.; Hederstedt, L. Enterococcus faecalis heme-dependent catalase. J. Bacteriol. 2002, 184, 6351–6356. [Google Scholar] [CrossRef] [Green Version]
- Wolff, S.P. Ferrous ion oxidation in presence of ferric ion indicator xylenol organge for measurement of hydroperoxides. Methods Enzymol. 1994, 233, 182–189. [Google Scholar]
- Deneer, H.G.; Boychuk, I. Reduction of ferric iron by Listeria monocytogenes and other species of Listeria. Can. J. Microbiol. 1993, 39, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Baureder, M.; Barane, E.; Hederstedt, L. In vitro assembly of catalase. J. Biol. Chem. 2014, 289, 28411–28420. [Google Scholar] [CrossRef] [Green Version]
- La Carbona, S.; Sauvageot, N.; Giard, J.-C.; Benachour, A.; Posteraro, B.; Auffray, Y.; Sanguinetti, M.; Hartke, A. Comparative study of the physiological roles of three peroxidases (NADH peroxidase, alkyl hydroperoxidase reductase and thiol peroxidase) in oxidative stress response, survival inside macrophages and virulence of Enteroccus faecalis. Mol. Microbiol. 2007, 66, 1148–1163. [Google Scholar] [CrossRef] [PubMed]
- Wasselin, V.; Budin-Verneuil, A.; Rincé, I.; Léger, L.; Boukerb, A.; Hartke, A.; Benachour, A.; Riboulet-Bisson, E. The enigmatic physiological roles of AhpCF, Gpx, Npr and Kat in peroxide stress response of Enterococcus faecalis. Res. Microbiol. 2022, 173, 103982. [Google Scholar] [CrossRef] [PubMed]
- Lam, L.N.; Wong, J.J.; Matysik, A.; Paxman, J.J.; Chong, L.K.K.; Low, P.M.; Chua, Z.S.; Heras, B.; Marsili, E.; Kline, K.A. Sortase-assembled pili promote extracellular electron transfer and iron acquisition in Enterococcus faecalis biofilm. bioRxiv 2019, 601666. [Google Scholar] [CrossRef] [Green Version]
- Light, S.H.; Su, L.; Rivera-Lugo, R.; Cornejo, J.A.; Louie, A.; Iavarone, A.T.; Ajo-Franklin, C.M.; Portnoy, D.A. A flavin-based extracellular electron transfer mechanism in diverse Gram-positive bacteria. Nature 2018, 562, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Bizzini, A.; Zhao, C.; Budin-Verneuil, A.; Sauvageot, N.; Giard, J.-C.; Auffray, Y.; Hartke, A. Glycerol is metabolized in a complex and strain-dependent manner in Enterococcus faecalis. J. Bacteriol. 2010, 192, 779–785. [Google Scholar] [CrossRef] [Green Version]
- Naraki, S.; Igimi, S.; Sasaki, Y. NADH peroxidase plays a crucial role in consuming H2O2 in Lactobacillus casei IGM394. Biosci. Microbiota Food Health 2020, 39, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Borisov, V.B.; Siletsky, S.A.; Nastasi, M.R.; Forte, E. ROS defense systems and terminal oxidases in bacteria. Antioxidants 2021, 10, 839. [Google Scholar] [CrossRef]
- Huycke, M.M.; Moore, D.; Joyce, W.; Wise, P.; Shepard, L.; Kotake, Y.; Gilmore, M.S. Extracellular superoxide production by Enterococcus faecalis requires demethylmenaquinone and is attenuated by functional terminal quinol oxidases. Mol. Microbiol. 2008, 42, 729–740. [Google Scholar] [CrossRef]
- Meyerstein, D. What are the oxidizing intermediates in the Fenton and Fenton-like reactions? A perspective. Antioxidants 2022, 11, 1368. [Google Scholar] [CrossRef] [PubMed]
- Baureder, M.; Reimann, R.; Hederstedt, L. Contribution of catalase to hydrogen peroxide resistance in Enterococcus faecalis. FEMS Microbiol. Lett. 2012, 331, 160–164. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, A.; Adi, P.; Thi, T.; Yang, J.-H.; Labibah, A.; Huang, C.-M. Skin bacteria mediate glycerol fermentation to produce electricity and resist UV-B. Microorganisms 2020, 8, 1092. [Google Scholar] [CrossRef]
- Riboulet, E.; Verneuil, N.; La Carbona, S.; Sauvageot, N.; Auffray, Y.; Hartke, A.; Giard, J.-C. Relationships between oxidative stress response and virulence in Enterococcus faecalis. J. Mol. Biol. Biotech. 2007, 13, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.L.; Schmidt, T.M. Shallow breathing: Bacterial life at low O2. Nat. Rev. Microbiol. 2013, 11, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Lugo, R.; Huang, S.; Lee, F.; Méheust, R.; Iavarone, A.T.; Sidebottom, A.M.; Oldfield, E.; Portnoy, D.A.; Light, S.H. Distinct energy-coupling factor transporter subunits enable flavin acquisition and extracytoplasmic trafficking for extracellular electron transfer in Listeria monocytogenes. bioRxiv 2022. [Google Scholar] [CrossRef]
- Méheust, R.; Huang, S.; Rivera-Lugo, R.; Banfield, J.F.; Light, S.H. Post-translational flavinylation is associated with diverse extracytosolic redox functionalities throughout bacterial life. eLife 2021, 10, e66878. [Google Scholar] [CrossRef]
- Tolar, J.G.; Li, S.; Ajo-Franklin, C.M. The differing roles of flavins and quinones in extracellular electron transfer in Lactiplantibacillus plantarum. Appl. Environ. Microbiol. 2022, e01313-22, in press. [Google Scholar] [CrossRef]
- Ch’ng, J.H.; Muthu, M.; Chong, K.K.L.; Wong, J.J.; Tan, C.A.Z.; Koh, Z.J.S.; Lopez, D.; Matysik, A.; Nair, Z.J.; Barkham, T.; et al. Heme cross-feeding can augment Staphylococcus aureus and Enterococcus faecalis dual species biofilms. ISME J. 2022, 16, 2015–2016. [Google Scholar] [CrossRef]
- Franza, T.; Delavenne, E.; Derré-Bobillot, A.; Juillard, V.; Boulay, M.; Demey, E.; Vinh, J.; Lamberet, G.; Gaudu, P. A partial metabolic pathway enables group b streptococcus to overcome quinone deficiency in a host bacterial community. Mol. Microbiol. 2016, 102, 81–91. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain a | Gene Defect | TSBG + Fe | TSBG + Fe + Hemin |
|---|---|---|---|
| OG1RF | (wild-type) | + | ++ |
| LFR20 | npr | − | ++(+) |
| LFR21 | npr | − | ++(+) |
| LFR24 | eetB | − | − |
| LFR30 | ndh3 | − | − |
| EMB1 | katA | + | ++ |
| EMB4 | cydC katA | + | +(−) |
| EMB7 | npr | − | ++(+) |
| EMB8 | npr | − | ++(+) |
| EMB15 | npr | − | ++(+) |
| EMB17 | katA | + | ++ |
| EMB20 | npr | − | ++(+) |
| EMB37 | npr | − | ++(+) |
| EMB44 | cydABCD | + | +(−) |
| JH2-2 | (wild-type) | + | ++ |
| JH2-2 Δnpr | npr | − | ++(+) |
| WY84 | menB | +(−) | +(−) |
| Strain | Gene | EfaMarTn Insertion Position a | Locus Tag |
|---|---|---|---|
| LFR20 | npr | 1.177.185 | OG1RF_10983 |
| LFR21 | npr | 1.176.588 | OG1RF_10983 |
| LFR24 | eetB | 3.137.739 | OG1RF_12512 |
| LFR30 | ndh3 | 3.135.429 | OG1RF_12510 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hederstedt, L. Enterococcus faecalis NADH Peroxidase-Defective Mutants Stain Falsely in Colony Zymogram Assay for Extracellular Electron Transfer to Ferric Ions. Microorganisms 2023, 11, 106. https://doi.org/10.3390/microorganisms11010106
Hederstedt L. Enterococcus faecalis NADH Peroxidase-Defective Mutants Stain Falsely in Colony Zymogram Assay for Extracellular Electron Transfer to Ferric Ions. Microorganisms. 2023; 11(1):106. https://doi.org/10.3390/microorganisms11010106
Chicago/Turabian StyleHederstedt, Lars. 2023. "Enterococcus faecalis NADH Peroxidase-Defective Mutants Stain Falsely in Colony Zymogram Assay for Extracellular Electron Transfer to Ferric Ions" Microorganisms 11, no. 1: 106. https://doi.org/10.3390/microorganisms11010106
APA StyleHederstedt, L. (2023). Enterococcus faecalis NADH Peroxidase-Defective Mutants Stain Falsely in Colony Zymogram Assay for Extracellular Electron Transfer to Ferric Ions. Microorganisms, 11(1), 106. https://doi.org/10.3390/microorganisms11010106