

Efficacy of Different Encapsulation Techniques on the Viability and Stability of Diverse Phage under Simulated Gastric Conditions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria and Bacteriophage
2.2. Phage Propagation and Concentration
2.3. Bacteriophage DNA Genome Extraction
2.4. DNA Sequencing & Assembly
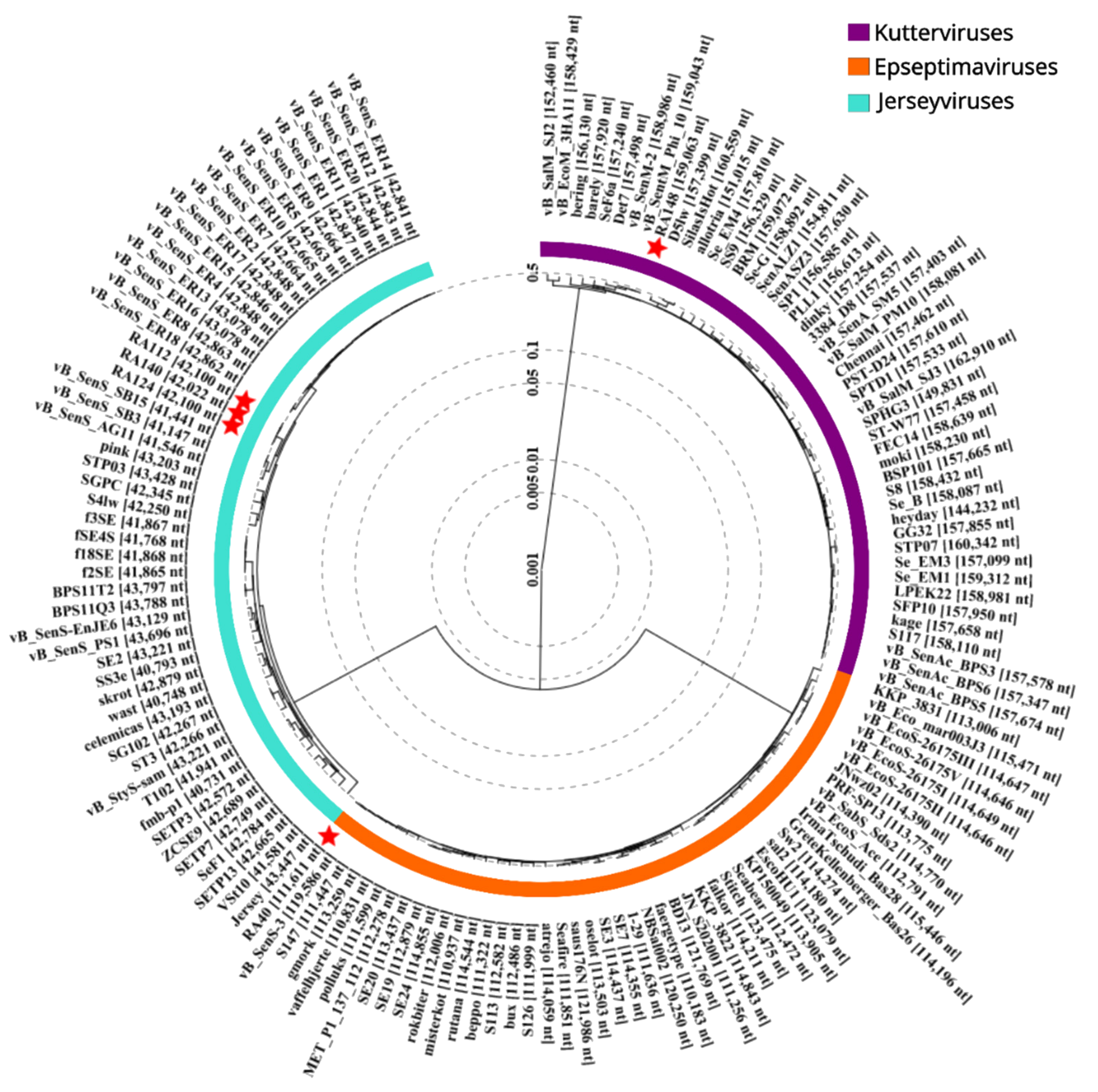
2.5. Phage Phylogenetic Analysis
2.6. Encapsulation of Phage in Alginate–Carrageenan (ALG-CG)
2.7. Encapsulation Efficiency and Particle Size Measurement
2.8. Susceptibility of Free and Microencapsulated Phage to Simulated Gastric and Duodenum Conditions
2.9. Statistical Analysis
3. Results
3.1. Phage Genome Characteristics
3.2. Encapsulation Efficiency
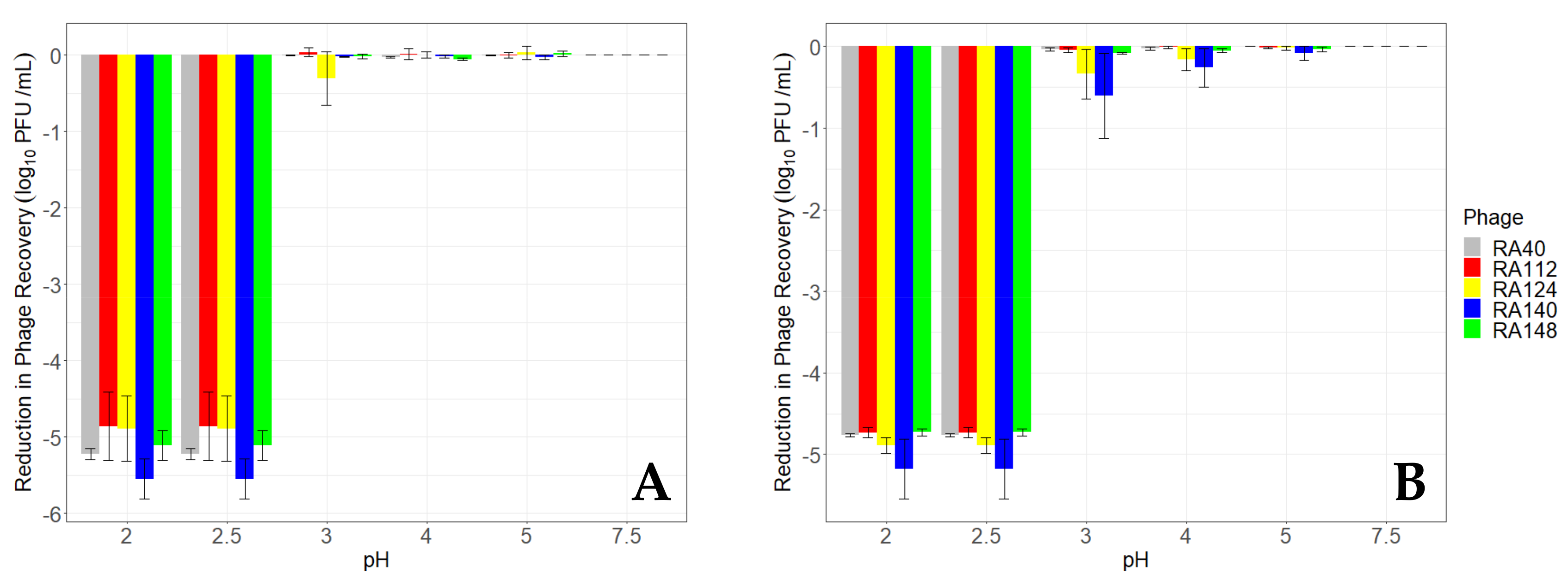
3.3. Viability of Unencapsulated Phage at Different pHs
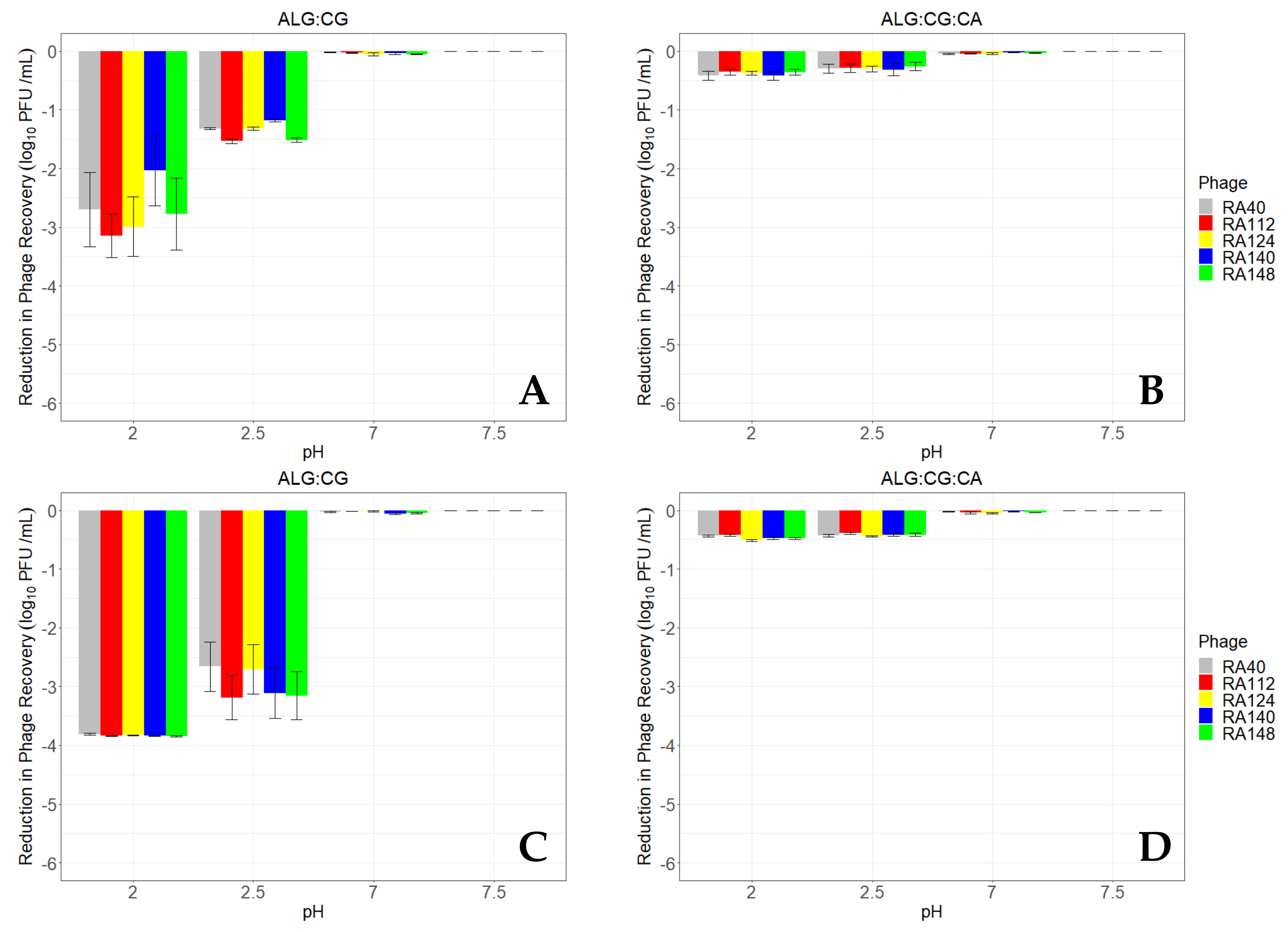
3.4. ALG-CG Microcapsules Can Protect Phage from a Simulated Gastric Environment
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Saha, M.; Sarkar, A. Review on Multiple Facets of Drug Resistance: A Rising Challenge in the 21st Century. J. Xenobiot. 2021, 11, 197–214. [Google Scholar] [CrossRef]
- Van, T.T.H.; Yidana, Z.; Smooker, P.M.; Coloe, P.J. Antibiotic Use in Food Animals Worldwide, with a Focus on Africa: Pluses and Minuses. J. Glob. Antimicrob. Resist. 2020, 20, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.G.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global Trends in Antimicrobial Resistance in Animals in Low- And Middle-Income Countries. Science 2019, 365, eaaw1944. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global Trends in Antimicrobial Use in Food Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Juniora, C.A. Worldwide Epidemiology of Salmonella Serovars in Animal-Based Foods: A Meta-Analysis. Appl. Environ. Microbiol. 2019, 85, e00591-19. [Google Scholar] [CrossRef] [PubMed]
- Machado Junior, P.C.; Chung, C.; Hagerman, A. Modeling Salmonella Spread in Broiler Production: Identifying Determinants and Control Strategies. Front. Vet. Sci. 2020, 7, 564. [Google Scholar] [CrossRef] [PubMed]
- McDermott, P.F.; Zhao, S.; Tate, H. Antimicrobial Resistance in Nontyphoidal Salmonella. Microbiol. Spectr. 2018, 6, ARBA-0014-2017. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2020/2021. EFSA J. 2023, 21, e07867. [Google Scholar] [CrossRef]
- Alba, P.; Leekitcharoenphon, P.; Carfora, V.; Amoruso, R.; Cordaro, G.; Di Matteo, P.; Ianzano, A.; Iurescia, M.; Diaconu, E.L.; Pedersen, S.K.; et al. Molecular Epidemiology of Salmonella Infantis in Europe: Insights into the Success of the Bacterial Host and Its Parasitic pESI-like Megaplasmid. Microb. Genomics 2020, 6, e000365. [Google Scholar] [CrossRef]
- Proietti, P.C.; Musa, L.; Stefanetti, V.; Orsini, M.; Toppi, V.; Branciari, R.; Blasi, F.; Magistrali, C.F.; Capomaccio, S.; Kika, T.S.; et al. Mcr-1-Mediated Colistin Resistance and Genomic Characterization of Antimicrobial Resistance in ESBL-Producing Salmonella Infantis Strains from a Broiler Meat Production Chain in Italy. Antibiotics 2022, 11, 728. [Google Scholar] [CrossRef]
- Gehring, R.; Mochel, J.P.; Schmerold, I. Understanding the Background and Clinical Significance of the WHO, WOAH, and EMA Classifications of Antimicrobials to Mitigate Antimicrobial Resistance. Front. Vet. Sci. 2023, 10, 1153048. [Google Scholar] [CrossRef] [PubMed]
- Czaplewski, L.; Bax, R.; Clokie, M.; Dawson, M.; Fairhead, H.; Fischetti, V.A.; Foster, S.; Gilmore, B.F.; Hancock, R.E.; Harper, D.; et al. Alternatives to Antibiotics-a Pipeline Portfolio Review. Lancet Infect. Dis. 2016, 16, 239–251. [Google Scholar] [CrossRef]
- Shaufi, M.A.M.; Sieo, C.C.; Chong, C.W.; Geok Hun, T.; Omar, A.R.; Han Ming, G.; Wan Ho, Y. Effects of Phage Cocktail, Probiotics, and Their Combination on Growth Performance and Gut Microbiota of Broiler Chickens. Animals 2023, 13, 1328. [Google Scholar] [CrossRef] [PubMed]
- Abd-El Wahab, A.; Basiouni, S.; El-Seedi, H.R.; Ahmed, M.F.E.; Bielke, L.R.; Hargis, B.; Tellez-Isaias, G.; Eisenreich, W.; Lehnherr, H.; Kittler, S.; et al. An Overview of the Use of Bacteriophages in the Poultry Industry: Successes, Challenges, and Possibilities for Overcoming Breakdowns. Front. Microbiol. 2023, 14, 1136638. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, V.; Baquero, D.; Hernandez, S.; Farfan, J.C.; Arias, J.; Arévalo, A.; Donado-Godoy, P.; Vives-Flores, M. Phage Cocktail SalmoFREE® Reduces Salmonella on a Commercial Broiler Farm. Poult. Sci. 2019, 98, 5054–5063. [Google Scholar] [CrossRef]
- Ackermann, H.-W.; DuBow, M.S. Viruses of Prokayotes: Natural Groups of Bacteriophages; CRC Press: Boca Raton, FL, USA, 1987; Volume 2. [Google Scholar]
- Malik, D.J.; Sokolov, I.J.; Vinner, G.K.; Mancuso, F.; Cinquerrui, S.; Vladisavljevic, G.T.; Clokie, M.R.J.; Garton, N.J.; Stapley, A.G.F.; Kirpichnikova, A. Formulation, Stabilisation and Encapsulation of Bacteriophage for Phage Therapy. Adv. Colloid Interface Sci. 2017, 249, 100–133. [Google Scholar] [CrossRef]
- Martinsen, T.C.; Bergh, K.; Waldum, H.L. Gastric Juice: A Barrier against Infectious Diseases. Basic Clin. Pharmacol. Toxicol. 2005, 96, 94–102. [Google Scholar] [CrossRef]
- Heo, J.M.; Opapeju, F.O.; Pluske, J.R.; Kim, J.C.; Hampson, D.J.; Nyachoti, C.M. Gastrointestinal Health and Function in Weaned Pigs: A Review of Feeding Strategies to Control Post-Weaning Diarrhoea without Using in-Feed Antimicrobial Compounds. J. Anim. Physiol. Anim. Nutr. 2013, 97, 207–237. [Google Scholar] [CrossRef]
- Yin, H.; Li, J.; Huang, H.; Wang, Y.; Qian, X.; Ren, J.; Xue, F.; Dai, J.; Tang, F. Microencapsulated Phages Show Prolonged Stability in Gastrointestinal Environments and High Therapeutic Efficiency to Treat Escherichia coli O157:H7 Infection. Vet. Res. 2021, 52, 118. [Google Scholar] [CrossRef]
- Lu, T.K.; Koeris, M.S. The next Generation of Bacteriophage Therapy. Curr. Opin. Microbiol. 2011, 14, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Petsong, K.; Benjakul, S.; Vongkamjan, K. Evaluation of Storage Conditions and Efficiency of a Novel Microencapsulated Salmonella Phage Cocktail for Controlling S. Enteritidis and S. Typhimurium in vitro and in Fresh Foods. Food Microbiol. 2019, 83, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Javaudin, F.; Bémer, P.; Batard, E.; Montassier, E. Impact of Phage Therapy on Multidrug-resistant Escherichia coli Intestinal Carriage in a Murine Model. Microorganisms 2021, 9, 2580. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.A.; Liu, H.; Wang, Q.; Zhong, F.; Guo, Q.; Balamurugan, S. Use of Encapsulated Bacteriophages to Enhance Farm to Fork Food Safety. Crit. Rev. Food Sci. Nutr. 2017, 57, 2801–2810. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Rebenaque, L.; Malik, D.J.; Catalá-Gregori, P.; Marin, C.; Sevilla-Navarro, S. Gastrointestinal Dynamics of Non-Encapsulated and Microencapsulated Salmonella Bacteriophages in Broiler Production. Animals 2022, 12, 144. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, D.; Yu, H.; Han, J.; Liu, W.; Qu, D. Encapsulation of Salmonella Phage SL01 in Alginate/Carrageenan Microcapsules as a Delivery System and Its Application in Vitro. Front. Microbiol. 2022, 13, 906103. [Google Scholar] [CrossRef] [PubMed]
- Raghav, N.; Vashisth, C.; Mor, N.; Arya, P.; Sharma, M.R.; Kaur, R.; Bhatti, S.P.; Kennedy, J.F. Recent Advances in Cellulose, Pectin, Carrageenan and Alginate-Based Oral Drug Delivery Systems. Int. J. Biol. Macromol. 2023, 244, 125357. [Google Scholar] [CrossRef] [PubMed]
- Naghizadeh, M.; Torshizi, M.A.K.; Rahimi, S.; Dalgaard, T.S. Synergistic Effect of Phage Therapy Using a Cocktail Rather than a Single Phage in the Control of Severe Colibacillosis in Quails. Poult. Sci. 2019, 98, 653–663. [Google Scholar] [CrossRef]
- Nale, J.Y.; Vinner, G.K.; Lopez, V.C.; Thanki, A.M.; Phothaworn, P.; Thiennimitr, P.; Garcia, A.; AbuOun, M.; Anjum, M.F.; Korbsrisate, S.; et al. An Optimized Bacteriophage Cocktail Can Effectively Control Salmonella in Vitro and in Galleria mellonella. Front. Microbiol. 2021, 11, 609955. [Google Scholar] [CrossRef]
- Chen, L.; Yuan, S.; Liu, Q.; Mai, G.; Yang, J.; Deng, D.; Zhang, B.; Liu, C.; Ma, Y. In Vitro Design and Evaluation of Phage Cocktails against Aeromonas salmonicida. Front. Microbiol. 2018, 9, 1476. [Google Scholar] [CrossRef]
- Richardson, E.J.; Limaye, B.; Inamdar, H.; Datta, A.; Manjari, K.S.; Pullinger, G.D.; Thomson, N.R.; Joshi, R.R.; Watson, M.; Stevens, M.P. Genome Sequences of Salmonella enterica Serovar Typhimurium, Choleraesuis, Dublin, and Gallinarum Strains of Well-Defined Virulence in Food-Producing Animals. J. Bacteriol. 2011, 193, 3162–3163. [Google Scholar] [CrossRef] [PubMed]
- Van Twest, R.; Kropinski, A.M. Bacteriophage Enrichment from Water and Soil. Methods Mol. Biol. 2009, 501, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D.W. Precipitation of Bacteriophage λ Particles from Large-Scale Lysates. Cold Spring Harb. Protoc. 2006, 2006, pdb.prot3966. [Google Scholar] [CrossRef]
- Hooton, S.; D’Angelantonio, D.; Hu, Y.; Connerton, P.L.; Aprea, G.; Connerton, I.F. Campylobacter Bacteriophage DA10: An Excised Temperate Bacteriophage Targeted by CRISPR-Cas. BMC Genomics 2020, 21, 400. [Google Scholar] [CrossRef] [PubMed]
- Arkin, A.P.; Cottingham, R.W.; Henry, C.S.; Harris, N.L.; Stevens, R.L.; Maslov, S.; Dehal, P.; Ware, D.; Perez, F.; Canon, S.; et al. KBase: The United States Department of Energy Systems Biology Knowledgebase. Nat. Biotechnol. 2018, 36, 566–569. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A Comparative Pathogenomic Platform with an Interactive Web Interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef] [PubMed]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2019, 48, D517–D525. [Google Scholar] [CrossRef]
- Shang, J.; Peng, C.; Liao, H.; Tang, X.; Sun, Y. PhaBOX: A Web Server for Identifying and Characterizing Phage Contigs in Metagenomic Data. arXiv 2023, arXiv:2303.15707. [Google Scholar] [CrossRef]
- Garneau, J.R.; Depardieu, F.; Fortier, L.C.; Bikard, D.; Monot, M. PhageTerm: A Tool for Fast and Accurate Determination of Phage Termini and Packaging Mechanism Using next-Generation Sequencing Data. Sci. Rep. 2017, 7, 8292. [Google Scholar] [CrossRef]
- Nishimura, Y.; Yoshida, T.; Kuronishi, M.; Uehara, H.; Ogata, H.; Goto, S. ViPTree: The Viral Proteomic Tree Server. Bioinformatics 2017, 33, 2379–2380. [Google Scholar] [CrossRef] [PubMed]
- Samtlebe, M.; Ergin, F.; Wagner, N.; Neve, H.; Küçükçetin, A.; Franz, C.M.A.P.; Heller, K.J.; Hinrichs, J.; Atamer, Z. Carrier Systems for Bacteriophages to Supplement Food Systems: Encapsulation and Controlled Release to Modulate the Human Gut Microbiota. LWT 2016, 68, 334–340. [Google Scholar] [CrossRef]
- Kovacs-Nolan, J.; Mine, Y. Microencapsulation for the Gastric Passage and Controlled Intestinal Release of Immunoglobulin Y. J. Immunol. Methods 2005, 296, 199–209. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 16 August 2023).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Gendre, J.; Ansaldi, M.; Olivenza, D.R.; Denis, Y.; Casadesús, J.; Ginet, N. Genetic Mining of Newly Isolated Salmophages for Phage Therapy. Int. J. Mol. Sci. 2022, 23, 8917. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Morello, S.; Pinto, A.; Del Gaudio, P.; Auriemma, G.; Aquino, R.P. Zinc and Calcium Cations Combination in the Production of Floating Alginate Beads as Prednisolone Delivery Systems. Molecules 2020, 25, 1140. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Pacan, J.C.; Wang, Q.; Sabour, P.M.; Huang, X.; Xu, Y. Enhanced Alginate Microspheres as Means of Oral Delivery of Bacteriophage for Reducing Staphylococcus aureus Intestinal Carriage. Food Hydrocoll. 2012, 26, 434–440. [Google Scholar] [CrossRef]
- Gencay, Y.E.; Gambino, M.; Prüssing, T.F.; Brøndsted, L. The Genera of Bacteriophages and Their Receptors Are the Major Determinants of Host Range. Environ. Microbiol. 2019, 21, 2095–2111. [Google Scholar] [CrossRef]
- Tao, C.; Yi, Z.; Zhang, Y.; Wang, Y.; Zhu, H.; Afayibo, D.J.A.; Li, T.; Tian, M.; Qi, J.; Ding, C.; et al. Characterization of a Broad-Host-Range Lytic Phage SHWT1 Against Multidrug-Resistant Salmonella and Evaluation of Its Therapeutic Efficacy in vitro and in vivo. Front. Vet. Sci. 2021, 8, 683853. [Google Scholar] [CrossRef]
- Li, Y.; Lv, P.; Shi, D.; Zhao, H.; Yuan, X.; Jin, X.; Wang, X. A Cocktail of Three Virulent Phages Controls Multidrug-Resistant Salmonella Enteritidis Infection in Poultry. Front. Microbiol. 2022, 13, 940525. [Google Scholar] [CrossRef]
- Ding, Y.; Huang, C.; Zhu, W.; Li, Z.; Zhang, Y.; Wang, J.; Pan, H.; Li, H.; Wang, X. Characterization of a Novel Jerseyvirus Phage T102 and Its Inhibition Effect on Biofilms of Multidrug-Resistant Salmonella. Virus Res. 2023, 326, 199054. [Google Scholar] [CrossRef]
- Duc, H.M.; Zhang, Y.; Son, H.M.; Huang, H.H.; Masuda, Y.; Honjoh, K.I.; Miyamoto, T. Genomic Characterization and Application of a Novel Bacteriophage STG2 Capable of Reducing Planktonic and Biofilm Cells of Salmonella. Int. J. Food Microbiol. 2023, 385, 109999. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Ji, H.; Li, X.; Ke, X.; Li, X.; Chen, P.; Qian, P. Host Receptor Identification of a Polyvalent Lytic Phage GSP044, and Preliminary Assessment of Its Efficacy in the Clearance of Salmonella. Microbiol. Res. 2023, 273, 127412. [Google Scholar] [CrossRef] [PubMed]
- Yamaki, S.; Yamazaki, K.; Kawai, Y. Broad Host Range Bacteriophage, EscoHU1, Infecting Escherichia coli O157:H7 and Salmonella enterica: Characterization, Comparative Genomics, and Applications in Food Safety. Int. J. Food Microbiol. 2022, 372, 109680. [Google Scholar] [CrossRef] [PubMed]
- Hooton, S.P.T.; Timms, A.R.; Rowsell, J.; Wilson, R.; Connerton, I.F. Salmonella Typhimurium-Specific Bacteriophage ΦSH19 and the Origins of Species Specificity in the Vi01-like Phage Family. Virol. J. 2011, 8, 498. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, A.N.; Woudstra, C.; Sørensen, M.C.H.; Brøndsted, L. Subtypes of Tail Spike Proteins Predicts the Host Range of Ackermannviridae Phages. Comput. Struct. Biotechnol. J. 2021, 19, 4854–4867. [Google Scholar] [CrossRef] [PubMed]
- Casjens, S.R.; Jacobs-Sera, D.; Hatfull, G.F.; Hendrix, R.W. Genome Sequence of Salmonella enterica Phage Det7. Genome Announc. 2015, 3, e00279-15. [Google Scholar] [CrossRef]
- Walter, M.; Fiedler, C.; Grassl, R.; Biebl, M.; Rachel, R.; Hermo-Parrado, X.L.; Llamas-Saiz, A.L.; Seckler, R.; Miller, S.; van Raaij, M.J. Structure of the Receptor-Binding Protein of Bacteriophage Det7: A Podoviral Tail Spike in a Myovirus. J. Virol. 2008, 82, 2265–2273. [Google Scholar] [CrossRef]
- Henze, L.J.; Koehl, N.J.; Bennett-Lenane, H.; Holm, R.; Grimm, M.; Schneider, F.; Weitschies, W.; Koziolek, M.; Griffin, B.T. Characterization of Gastrointestinal Transit and Luminal Conditions in Pigs Using a Telemetric Motility Capsule. Eur. J. Pharm. Sci. 2021, 156, 105627. [Google Scholar] [CrossRef]
- Lee, S.A.; Dunne, J.; Febery, E.; Wilcock, P.; Mottram, T.; Bedford, M.R. Superdosing Phytase Reduces Real-Time Gastric pH in Broilers and Weaned Piglets. Br. Poult. Sci. 2018, 59, 330–339. [Google Scholar] [CrossRef]
- Loveday, S.M. Protein Digestion and Absorption: The Influence of Food Processing. Nutr. Res. Rev. 2022, 1–16. [Google Scholar] [CrossRef]
- Jończyk, E.; Kłak, M.; Międzybrodzki, R.; Górski, A. The Influence of External Factors on Bacteriophages—Review. Folia Microbiol. 2011, 56, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Garcia, J.; Chavez-Carbajal, A.; Segundo-Arizmendi, N.; Baron-Pichardo, M.G.; Mendoza-Elvira, S.E.; Hernandez-Baltazar, E.; Hynes, A.P.; Torres-Angeles, O. Efficacy of Salmonella Bacteriophage S1 Delivered and Released by Alginate Beads in a Chicken Model of Infection. Viruses 2021, 13, 1932. [Google Scholar] [CrossRef] [PubMed]
- Colom, J.; Cano-Sarabia, M.; Otero, J.; Aríñez-Soriano, J.; Cortés, P.; Maspoch, D.; Llagostera, M. Microencapsulation with Alginate/CaCO3: A Strategy for Improved Phage Therapy. Sci. Rep. 2017, 7, 41441. [Google Scholar] [CrossRef] [PubMed]
- Svihus, B.; Hetland, H.; Choct, M.; Sundby, F. Passage Rate through the Anterior Digestive Tract of Broiler Chickens Fed on Diets with Ground and Whole Wheat. Br. Poult. Sci. 2002, 43, 662–668. [Google Scholar] [CrossRef] [PubMed]
- Mosimann, S.; Desiree, K.; Ebner, P. Efficacy of Phage Therapy in Poultry: A Systematic Review and Meta-Analysis. Poult. Sci. 2021, 100, 101472. [Google Scholar] [CrossRef] [PubMed]
- Tennant, S.M.; Hartland, E.L.; Phumoonna, T.; Lyras, D.; Rood, J.I.; Robins-Browne, R.M.; van Driel, I.R. Influence of Gastric Acid on Susceptibility to Infection with Ingested Bacterial Pathogens. Infect. Immun. 2008, 76, 639–645. [Google Scholar] [CrossRef] [PubMed]
- Yasutomi, E.; Hoshi, N.; Adachi, S.; Otsuka, T.; Kong, L.; Ku, Y.; Yamairi, H.; Inoue, J.; Ishida, T.; Watanabe, D.; et al. Proton Pump Inhibitors Increase the Susceptibility of Mice to Oral Infection with Enteropathogenic Bacteria. Dig. Dis. Sci. 2018, 63, 881–889. [Google Scholar] [CrossRef]
- Roque-Borda, C.A.; Souza Saraiva, M.D.M.; Monte, D.F.M.; Rodrigues Alves, L.B.; de Almeida, A.M.; Ferreira, T.S.; de Lima, T.S.; Benevides, V.P.; Cabrera, J.M.; Claire, S.; et al. HPMCAS-Coated Alginate Microparticles Loaded with Ctx(Ile21)-Ha as a Promising Antimicrobial Agent against Salmonella Enteritidis in a Chicken Infection Model. ACS Infect. Dis. 2022, 8, 472–481. [Google Scholar] [CrossRef]
- Karamanolis, G.; Theofanidou, I.; Yiasemidou, M.; Giannoulis, E.; Triantafyllou, K.; Ladas, S.D. A glass of water immediately increases gastric pH in healthy subjects. Dig. Dis. Sci. 2008, 53, 3128–3132. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Phage | No. of Reads | No. of Subsampled Reads | Genome Size (bp) | × Coverage | Accession Number | Phage Genus |
|---|---|---|---|---|---|---|
| RA40 | 3.6 M | 300 K | 111,611 | ×26.47 | OR242313 | Epseptimavirus |
| RA112 | 4 M | 20 K | 42,100 | ×30.02 | OR242314 | Jerseyvirus |
| RA124 | 4.9 M | 20 K | 42,100 | ×29.9 | OR242315 | Jerseyvirus |
| RA140 | 3.7 M | 20 K | 42,022 | ×32.72 | OR242316 | Jerseyvirus |
| RA148 | 3.3 M | 1 M | 159,063 | ×29.02 | OR242317 | Kuttervirus |
| EE % ± SE | MPS ± SE | |||
|---|---|---|---|---|
| Phage | ALG-CG | ALG-CG-CA | ALG-CG | ALG-CG-CA |
| RA40 | 97.2 ± 0.84 | 97.0 ± 0.63 | 1.97 ± 0.02 *** | 1.78 ± 0.02 |
| RA112 | 98.5 ± 1.14 | 97.1 ± 1.28 | 1.90 ± 0.02 * | 1.78 ± 0.03 |
| RA124 | 98.2 ± 1.66 | 96.3 ± 0.83 | 1.94 ± 0.02 *** | 1.81 ± 0.02 |
| RA140 | 96.3 ± 1.30 | 98.6 ± 0.78 | 1.94 ± 0.02 *** | 1.80 ± 0.03 |
| RA148 | 95.2 ± 0.62 | 98.2 ± 0.81 | 1.92 ± 0.02 *** | 1.80 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dlamini, S.B.; Gigante, A.M.; Hooton, S.P.T.; Atterbury, R.J. Efficacy of Different Encapsulation Techniques on the Viability and Stability of Diverse Phage under Simulated Gastric Conditions. Microorganisms 2023, 11, 2389. https://doi.org/10.3390/microorganisms11102389
Dlamini SB, Gigante AM, Hooton SPT, Atterbury RJ. Efficacy of Different Encapsulation Techniques on the Viability and Stability of Diverse Phage under Simulated Gastric Conditions. Microorganisms. 2023; 11(10):2389. https://doi.org/10.3390/microorganisms11102389
Chicago/Turabian StyleDlamini, Sicelo B., Adriano M. Gigante, Steven P. T. Hooton, and Robert J. Atterbury. 2023. "Efficacy of Different Encapsulation Techniques on the Viability and Stability of Diverse Phage under Simulated Gastric Conditions" Microorganisms 11, no. 10: 2389. https://doi.org/10.3390/microorganisms11102389
APA StyleDlamini, S. B., Gigante, A. M., Hooton, S. P. T., & Atterbury, R. J. (2023). Efficacy of Different Encapsulation Techniques on the Viability and Stability of Diverse Phage under Simulated Gastric Conditions. Microorganisms, 11(10), 2389. https://doi.org/10.3390/microorganisms11102389