
Transcriptome Analysis Reveals the Involvement of Mitophagy and Peroxisome in the Resistance to QoIs in Corynespora cassiicola
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates
2.2. Sensitivity Test to Fungicides
2.3. Total RNA Extraction and RNA-Seq
2.4. Identification of DEGs and Assessment of GO and KEGG Enrichment
2.5. Quantitative RT-PCR (qPCR) Analysis
2.6. Sensitivity to Oxidative Stress
3. Results
3.1. The Sensitivity of RI and RII Strains to Fungicides
3.2. Sequencing Data Quality Assessment and Sequence Comparison
3.3. Identification and Analysis of Differentially Expressed Genes (DEGs)
3.4. KEGG Enrichment Analysis
3.5. GO Functional Annotation
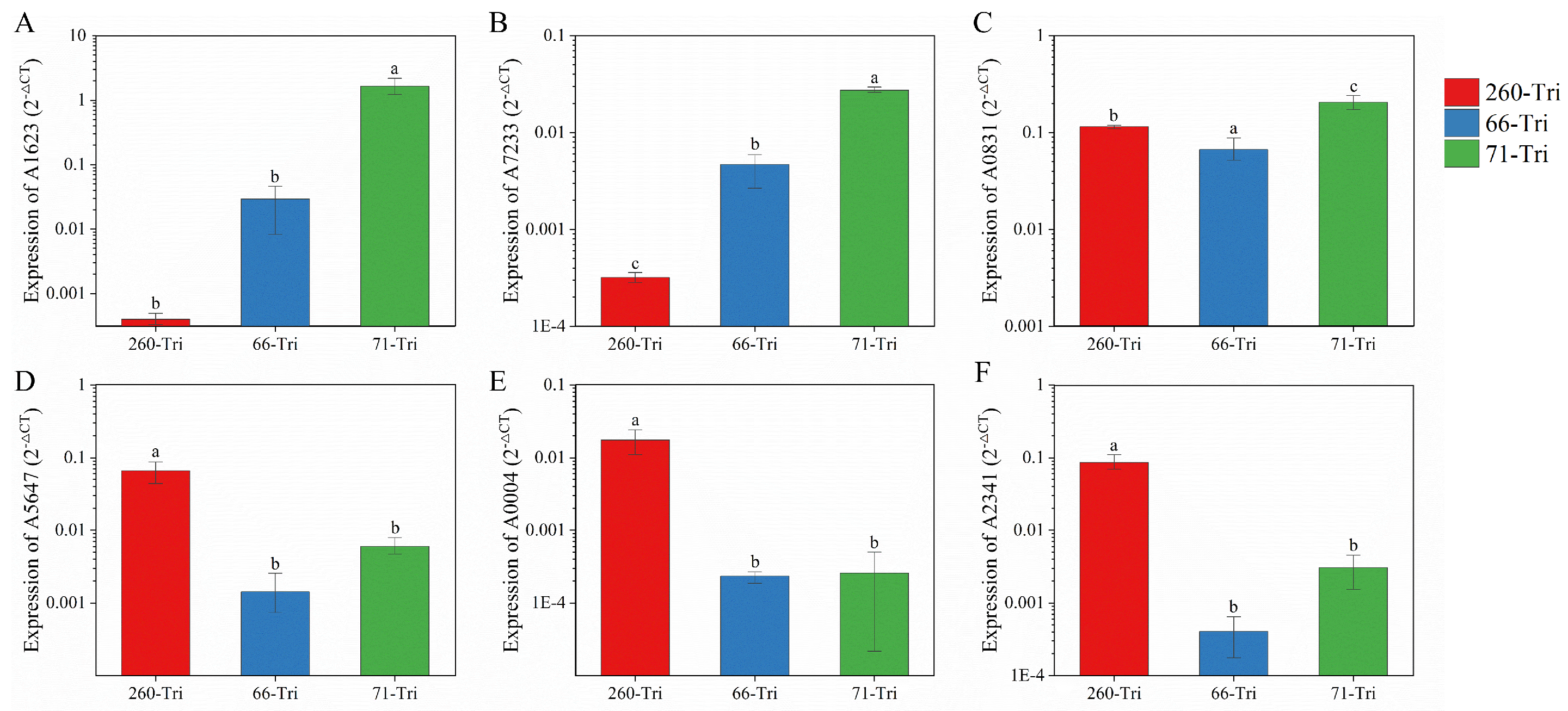
3.6. Real-Time Quantitative PCR (qRT-PCR) Validation of Transcriptomic Dates
3.7. Oxidative Stress
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | p-Value | Number |
|---|---|---|
| Ribosome | 74 | |
| Alanine, aspartate and glutamate metabolism | 0.046126 | 8 |
| Valine, leucine and isoleucine biosynthesis | 0.0616069 | 5 |
| Other glycan degradation | 0.0641183 | 4 |
| Glycosphingolipid biosynthesis—globo and isoglobo series | 0.0787579 | 3 |
| Pentose and glucuronate interconversions | 0.0979938 | 9 |
| Tryptophan metabolism | 0.107279 | 9 |
| Phenylalanine metabolism | 0.1135758 | 8 |
| Non-homologous end-joining | 0.123571 | 3 |
| Aminoacyl-tRNA biosynthesis | 0.1245206 | 8 |
| Tyrosine metabolism | 0.1384809 | 11 |
| Fatty acid degradation | 0.1396695 | 6 |
| 3-Oxocarboxylic acid metabolism | 0.1587337 | 7 |
| Carotenoid biosynthesis | 0.1619549 | 2 |
| Biosynthesis of amino acids | 0.1949744 | 18 |
| beta-Alanine metabolism | 0.2031291 | 6 |
| Arginine and proline metabolism | 0.2295227 | 8 |
| MAPK signaling pathway—yeast | 0.2319141 | 10 |
| Biosynthesis of antibiotics | 0.2343621 | 35 |
| Glycerolipid metabolism | 0.2381838 | 6 |
| Description | Term_Type | Corrected_p-Value | DEG_Item |
|---|---|---|---|
| translation | biological_process | 84 | |
| peptide biosynthetic process | biological_process | 84 | |
| amide biosynthetic process | biological_process | 85 | |
| peptide metabolic process | biological_process | 85 | |
| cellular amide metabolic process | biological_process | 86 | |
| organonitrogen compound biosynthetic process | biological_process | 124 | |
| cellular protein metabolic process | biological_process | 120 | |
| organonitrogen compound metabolic process | biological_process | 200 | |
| DNA integration | biological_process | 0.001247 | 17 |
| protein metabolic process | biological_process | 0.001247 | 143 |
| metabolic process | biological_process | 0.014184 | 498 |
| primary metabolic process | biological_process | 0.023669 | 375 |
| organic substance metabolic process | biological_process | 0.030208 | 397 |
| cellular nitrogen compound metabolic process | biological_process | 0.03328 | 253 |
| cellular nitrogen compound biosynthetic process | biological_process | 0.065234 | 176 |
| nitrogen compound metabolic process | biological_process | 0.082291 | 324 |
| small molecule metabolic process | biological_process | 0.09379 | 82 |
| biosynthetic process | biological_process | 0.10886 | 236 |
| organic substance biosynthetic process | biological_process | 0.13242 | 225 |
| nicotinamide metabolic process | biological_process | 0.15171 | 3 |
References
- Dixon, L.J.; Schlub, R.L.; Pernezny, K.; Datnoff, L.E. Host specialization and phylogenetic diversity of Corynespora cassiicola. Phytopathology 2009, 99, 1015–1027. [Google Scholar] [CrossRef] [PubMed]
- Doehlemann, G.; Ökmen, B.; Zhu, W.; Sharon, A. Plant Pathogenic Fungi. Microbiol. Spectr. 2017, 5, 5–51. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Ishii, H.; Stammler, G.; Koch, A.; Ogawara, T.; Tomita, Y.; Fountaine, J.M.; Ushio, S.; Seko, T.; Kobori, S. Distribution and molecular characterization of Corynespora cassiicola isolates resistant to boscalid. Plant Pathol. 2010, 59, 873–881. [Google Scholar] [CrossRef]
- Smith, L.J. Host Range, Phylogenetic, and Pathogenic Diversity of Corynespora cassiicola (Berk. & Curt.) Wei. Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2008. [Google Scholar]
- Barral, B.; Chillet, M.; Doizy, A.; Grassi, M.; Ragot, L.; Léchaudel, M.; Durand, N.; Rose, L.J.; Viljoen, A.; Schorr-Galindo, S. Diversity and Toxigenicity of Fungi that Cause Pineapple Fruitlet Core Rot. Toxins 2020, 12, 339. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, M.M.; Andolfi, A. Phytopathogenic Fungi and Toxicity. Toxins 2021, 13, 689. [Google Scholar] [CrossRef] [PubMed]
- Rahman, S.F.S.A.; Singh, E.; Pieterse, C.M.J.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, M.; Waśkiewicz, A.; Łukasz Stępień. Fusarium Cyclodepsipeptide Mycotoxins: Chemistry, Biosynthesis, and Occurrence. Toxins 2020, 12, 765. [Google Scholar] [CrossRef] [PubMed]
- Bruggen, A.H.C.V.; Gamliel, A.; Finckh, M.R. Plant disease management in organic farming systems. Pest Manag. Sci. 2016, 72, 30–44. [Google Scholar] [CrossRef]
- Van Bruggen, A.; Finckh, M.R. Plant Diseases and Management Approaches in Organic Farming Systems. Annu. Rev. Phytopathol. 2016, 54, 25–54. [Google Scholar] [CrossRef]
- Fernández-Ortuño, D.; Torés, J.A.; de Vicente, A.; Pérez-García, A. Mechanisms of resistance to QoI fungicides in phytopathogenic fungi. Int. Microbiol. 2008, 11, 1–9. [Google Scholar]
- Gisi, U.; Sierotzki, H.; Cook, A.; McCaffery, A.R. Mechanisms influencing the evolution of resistance to Qo inhibitor fungicides. Pest Manag. Sci. 2002, 58, 859–867. [Google Scholar] [CrossRef] [PubMed]
- Bolton, M.D.; Rivera, V.; Secor, G.A. Identification of the G143A mutation associated with QoI resistance in Cercospora beticola field isolates from Michigan, United States. Pest Manag. Sci. 2013, 69, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Castroagudín, V.L.; Ceresini, P.C.; de Oliveira, S.C.; Reges, J.T.A.; Maciel, J.L.N.; Bonato, A.L.V.; Dorigan, A.F.; McDonald, B.A. Resistance to QoI Fungicides Is Widespread in Brazilian Populations of the Wheat Blast Pathogen Magnaporthe oryzae. Phytopathology 2015, 105, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Xin, W.; Lu, F.; Li, T.; Li, M.; Wu, J.; Wang, J.; Zhou, M. Benzimidazole- and QoI-resistance in Corynespora cassiicola populations from greenhouse-cultivated cucumber: An emerging problem in China. Pestic. Biochem. Physiol. 2019, 153, 95–105. [Google Scholar] [CrossRef]
- Furuya, S.; Mochizuki, M.; Saito, S.; Kobayashi, H.; Takayanagi, T.; Suzuki, S. Monitoring of QoI fungicide resistance in Plasmopara viticola populations in Japan. Pest Manag. Sci. 2010, 66, 1268–1272. [Google Scholar] [CrossRef] [PubMed]
- Kiiker, R.; Juurik, M.; Heick, T.M.; Mäe, A. Changes in DMI, SDHI, and QoI Fungicide Sensitivity in the Estonian Zymoseptoria tritici Population between 2019 and 2020. Microorganisms 2021, 9, 814. [Google Scholar] [CrossRef] [PubMed]
- Lesniak, K.E.; Proffer, T.J.; Beckerman, J.; Sundin, G.W. Occurrence of QoI Resistance and Detection of the G143A Mutation in Michigan Populations of Venturia inaequalis. Plant Dis. 2011, 95, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Torriani, S.F.F.; Brunner, P.C.; McDonald, B.A.; Sierotzki, H. QoI resistance emerged independently at least 4 times in European populations of Mycosphaerella graminicola. Pest Manag. Sci. 2009, 65, 155–162. [Google Scholar] [CrossRef]
- Vega, B.; Dewdney, M.M. Distribution of QoI Resistance in Populations of Tangerine-Infecting Alternaria alternata in Florida. Plant Dis. 2014, 98, 67–76. [Google Scholar] [CrossRef]
- Vielba-Fernández, A.; Bellón-Gómez, D.; Torés, J.A.; de Vicente, A.; Pérez-García, A.; Fernández-Ortuño, D. Heteroplasmy for the Cytochrome b Gene in Podosphaera xanthii and its Role in Resistance to QoI Fungicides in Spain. Plant Dis. 2018, 102, 1599–1605. [Google Scholar] [CrossRef]
- Date, H.; Kataoka, E.; Tanina, K.; Sasaki, S.; Inoue, K.; Nasu, H.; Kasuyama, S. Sensitivity of Corynespora cassicola, causal agent of Corynespora leaf spot of cucumber, to thiophanate-methyl, diethofencarb and azoxystrobin. Ann. Phytopathol. Soc. Jpn. 2009, 70, 10–13. [Google Scholar] [CrossRef]
- Miguez, M.; Reeve, C.; Wood, P.M.; Hollomon, D.W. Alternative oxidase reduces the sensitivity of Mycosphaerella graminicola to QOI fungicides. Pest Manag. Sci. 2004, 60, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Olaya, G.; Zheng, D.; Köller, W. Differential responses of germinating Venturia inaequalis conidia to kresoxim-methyl. Pestic. Sci. 1998, 54, 230–236. [Google Scholar] [CrossRef]
- Wood, P.M.; Hollomon, D.W. A critical evaluation of the role of alternative oxidase in the performance of strobilurin and related fungicides acting at the Qo site of complex III. Pest Manag. Sci. 2003, 59, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Chen, S. Non-Target Site Mechanisms of Fungicide Resistance in Crop Pathogens: A Review. Microorganisms 2021, 9, 502. [Google Scholar] [CrossRef] [PubMed]
- Lalève, A.; Gamet, S.; Walker, A.S.; Debieu, D.; Toquin, V.; Fillinger, S. Site-directed mutagenesis of the P225, N230 and H272 residues of succinate dehydrogenase subunit B from Botrytis cinerea highlights different roles in enzyme activity and inhibitor binding. Environ. Microbiol. 2014, 16, 2253–2266. [Google Scholar] [CrossRef] [PubMed]
- Sierotzki, H.; Scalliet, G. A review of current knowledge of resistance aspects for the next-generation succinate dehydrogenase inhibitor fungicides. Phytopathology 2013, 103, 880–887. [Google Scholar] [CrossRef]
- Stammler, G.; Brix, H.D.; Nave, B.; Gold, R.; Schoefl, U.; Dehne, H.W.; Deising, H.B.; Gisi, U.; Kuck, K.H.; Russell, P.E. Studies on the Biological Performance of Boscalid and Its Mode of Action; Deutsche Phytomedizinische Gesellschaft e.V. Verlag: Braunschweig, Germany, 2008. [Google Scholar]
- Hu, M.; Cosseboom, S.D.; Schnabel, G. atrB-Associated Fludioxonil Resistance in Botrytis fragariae Not Linked to Mutations in Transcription Factor mrr1. Phytopathology 2019, 109, 839–846. [Google Scholar] [CrossRef]
- Guan, Y.; Wang, Y.; Li, B.; Shen, K.; Li, Q.; Ni, Y.J.; Huang, L. Mitophagy in carcinogenesis, drug resistance and anticancer therapeutics. Cancer Cell Int. 2021, 21, 350. [Google Scholar] [CrossRef]
- Yao, N.; Wang, C.; Hu, N.; Li, Y.; Liu, M.; Lei, Y.; Chen, M.; Chen, L.; Chen, C.; Lan, P.; et al. Inhibition of PINK1/Parkin-dependent mitophagy sensitizes multidrug-resistant cancer cells to B5G1, a new betulinic acid analog. Cell Death Dis. 2019, 10, 232. [Google Scholar] [CrossRef]
- Yang, S.L.; Yu, P.L.; Chung, K.R. The glutathione peroxidase-mediated reactive oxygen species resistance, fungicide sensitivity and cell wall construction in the citrus fungal pathogen Alternaria alternata. Environ. Microbiol. 2016, 18, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [PubMed]
- Minibayeva, F.V.; Dmitrieva, S.N.; Ponomareva, A.; Ryabovol, V. Oxidative stress-induced autophagy in plants: The role of mitochondria. Plant Physiol. Biochem. 2012, 59, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Narendra, D.P.; Jin, S.M.; Tanaka, A.; Suen, D.F.; Gautier, C.A.; Shen, J.; Cookson, M.R.; Youle, R.J.; Green, D.R. PINK1 Is Selectively Stabilized on Impaired Mitochondria to Activate Parkin. PLoS Biol. 2010, 8, e1000298. [Google Scholar] [CrossRef] [PubMed]
- Boyle, K.A.; Wickle, J.D.V.; Hill, R.B.; Marchese, A.; Kalyanaraman, B.; Dwinell, M.B. Mitochondria-targeted drugs stimulate mitophagy and abrogate colon cancer cell proliferation. J. Biol. Chem. 2018, 293, 14891–14904. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.C.; Chang, H.S.; Yeh, C.Y.; Chang, H.J.; Cheng, L.W.; Lin, T.T.; Liu, C.; Chen, S.T. Regulation of mitochondrial fusion and mitophagy by intra-tumoral delivery of membrane-fused mitochondria or Midiv-1 enhances sensitivity to doxorubicin in triple-negative breast cancer. Biomed. Pharmacother. 2022, 153, 113484. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, L.; Teng, Y.; Yin, H.; Wu, Q. PIWIL1 Drives Chemoresistance in Multiple Myeloma by Modulating Mitophagy and the Myeloma Stem Cell Population. Front. Oncol. 2022, 11, 783583. [Google Scholar] [CrossRef]
- Zheng, Y.; Huang, C.; Lu, L.; Yu, K.; Zhao, J.; Chen, M.; Liu, L.; Sun, Q.; Lin, Z.; Zheng, J.; et al. STOML2 potentiates metastasis of hepatocellular carcinoma by promoting PINK1-mediated mitophagy and regulates sensitivity to lenvatinib. J. Hematol. Oncol. 2021, 14, 16. [Google Scholar] [CrossRef]
- Sun, Y.; Shen, W.; Hu, S.; Lyu, Q.; Wang, Q.; Wei, T.; Zhu, W.; Zhang, J. METTL3 promotes chemoresistance in small cell lung cancer by inducing mitophagy. J. Exp. Clin. Cancer Res. 2023, 42, 65. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, L.; Li, Y.; Gao, J. Melatonin increases human cervical cancer HeLa cells apoptosis induced by cisplatin via inhibition of JNK/Parkin/mitophagy axis. Vitr. Cell. Dev. Biol. Anim. 2017, 54, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Sun, L.; Zhang, Y.; Wang, Y.; Zheng, J. Imbalanced GSH/ROS and sequential cell death. J. Biochem. Mol. Toxicol. 2021, 36, e22942. [Google Scholar] [CrossRef] [PubMed]
- Mhamdi, A.; Noctor, G.; Baker, A. Plant catalases: Peroxisomal redox guardians. Arch. Biochem. Biophys. 2012, 525, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Niu, B.; Liao, K.; Zhou, Y.; Wen, T.; Quan, G.; Pan, X.; Wu, C. Application of glutathione depletion in cancer therapy: Enhanced ROS-based therapy, ferroptosis, and chemotherapy. Biomaterials 2021, 277, 121110. [Google Scholar] [CrossRef] [PubMed]
- Paku, M.; Haraguchi, N.; Takeda, M.; Fujino, S.; Ogino, T.; Takahashi, H.; Miyoshi, N.; Uemura, M.; Mizushima, T.; Yamamoto, H.; et al. SIRT3-Mediated SOD2 and PGC-1α Contribute to Chemoresistance in Colorectal Cancer Cells. Ann. Surg. Oncol. 2021, 28, 4720–4732. [Google Scholar] [CrossRef] [PubMed]
- Spitz, D.R.; Phillips, J.W.; Adams, D.T.; Sherman, C.M.; Deen, D.F.; Li, G.C. Cellular resistance to oxidative stress is accompanied by resistance to cisplatin: The significance of increased catalase activity and total glutathione in hydrogen peroxide-resistant fibroblasts. J. Cell. Physiol. 1993, 156, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Jogaiah, S.; Mitani, S.; Nagaraj, A.K.; Shekar, S.H. Activity of cyazofamid against Sclerospora graminicola, a downy mildew disease of pearl millet. Pest Manag. Sci. 2007, 63, 722–727. [Google Scholar] [CrossRef]
- Mathieu, C.; Messaoudi, S.; Fattal, E.; Vergnaud-Gauduchon, J. Cancer drug resistance: Rationale for drug delivery systems and targeted inhibition of HSP90 family proteins. Cancer Drug Resist. 2019, 2, 18. [Google Scholar] [CrossRef]
- Zhou, F.; Cui, Y.X.; Wang, B.; Zhou, Y.D.; Li, S.W.; Zhang, Y.T.; Zhang, K.Y.; Chen, Z.Y.; Hu, H.; Li, C.W. Baseline sensitivity and potential resistance mechanisms for Fusarium pseudograminearum to fludioxonil. Plant Dis. 2022, 106, 2138–2144. [Google Scholar] [CrossRef]
- Yamamoto, E.S.; de Jesus, J.A.; Bezerra-Souza, A.; Brito, J.R.; Lago, J.H.G.; Laurenti, M.D.; Passero, L.F.D. Tolnaftate inhibits ergosterol production and impacts cell viability of Leishmania sp. Bioorganic Chem. 2020, 102, 104056. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Imran, M.; Xiao, L.; Hú, Z.; Li, G.; Zhang, F.; Liu, X. Difenoconazole Resistance Shift in Botrytis cinerea from Tomato in China Associated with Inducible Expression of CYP51. Plant Dis. 2020, 105, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hou, Y.; Mao, X.; Liu, F.; guo Zhou, M. Temperature-responded biological fitness of carbendazim-resistance Fusarium graminearum mutants conferring the F167Y, E198K and E198L substitutions. Plant Dis. 2021, 105, 3522–3530. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Zhu, G.; Xie, X.; Chai, A.; Li, L.; Shi, Y.; Li, B. Double Mutations in Succinate Dehydrogenase Are Involved in SDHI Resistance in Corynespora cassiicola. Microorganisms 2022, 10, 132. [Google Scholar] [CrossRef] [PubMed]
- Ishii, H.; Yano, K.; Date, H.; Furuta, A.; Sagehashi, Y.; Yamaguchi, T.; Sugiyama, T.; Nishimura, K.; Hasama, W. Molecular Characterization and Diagnosis of QoI Resistance in Cucumber and Eggplant Fungal Pathogens. Phytopathology 2007, 97, 1458–1466. [Google Scholar] [CrossRef] [PubMed]
- Semar, M.; Strobel, D.; Koch, A.; Klappach, K.; Stammler, G. Field efficacy of pyraclostrobin against populations of Pyrenophora teres containing the F129L mutation in the cytochrome b gene. J. Plant Dis. Prot. 2007, 114, 117–119. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, L.; Hu, J.; Duan, H.; Lin, D.X.; Liu, P.; Meng, Q.; Li, B.; Si, N.; Liu, C.; et al. Resistance Mechanisms and Molecular Docking Studies of Four Novel QoI Fungicides in Peronophythora litchii. Sci. Rep. 2015, 5, 17466. [Google Scholar] [CrossRef]
- Ashrafi, G.; Schwarz, T.L.; Schwarz, T. The pathways of mitophagy for quality control and clearance of mitochondria. Cell Death Differ. 2012, 20, 31–42. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, Q.; Gao, W.; Sehgal, S.A.; Wu, H. The multifaceted regulation of mitophagy by endogenous metabolites. Autophagy 2021, 18, 1216–1239. [Google Scholar] [CrossRef]
- Ferro, F.; Servais, S.; Besson, P.; Roger, S.; Dumas, J.F.; Brisson, L. Autophagy and mitophagy in cancer metabolic remodelling. Semin. Cell Dev. Biol. 2020, 98, 129–138. [Google Scholar] [CrossRef]
- Fivenson, E.M.; Lautrup, S.; Sun, N.; Scheibye-Knudsen, M.; Stevnsner, T.; Nilsen, H.; Bohr, V.A.; Fang, E.F. Mitophagy in neurodegeneration and aging. Neurochem. Int. 2017, 109, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhou, X.; Wang, Y.; Cao, H.; Guo, J.; Wang, P.; Yang, Y. Mitophagy—A New Target of Bone Disease. Biomolecules 2022, 12, 1420. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chu, Y.; Li, S.; Yu, L.; Deng, H.; Liao, C.; Liao, X.; Yang, C.M.; Qi, M.; Cheng, J.; et al. The oncoprotein MUC1 facilitates breast cancer progression by promoting Pink1-dependent mitophagy via ATAD3A destabilization. Cell Death Dis. 2022, 13, 899. [Google Scholar] [CrossRef] [PubMed]
- Wei, R.; Cao, J.; Yao, S. RETRACTED ARTICLE: Matrine promotes liver cancer cell apoptosis by inhibiting mitophagy and PINK1/Parkin pathways. Cell Stress Chaperones 2018, 23, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.T.; Teng, W.; Jeng, W.; Lin, C.; Lin, S.M.; Sheen, I.S. Combining immune checkpoint inhibitor with lenvatinib prolongs survival than lenvatinib alone in sorafenib-experienced hepatocellular carcinoma patients. Eur. J. Gastroenterol. Hepatol. 2020, 34, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cantero, A.; Fernández, L.L.; Guarro-Artigas, J.; Capilla, J. Azole resistance mechanisms in Aspergillus: Update and recent advances. Int. J. Antimicrob. Agents 2019, 55, 105807. [Google Scholar] [CrossRef] [PubMed]
- Belgareh-Touzé, N.; Cavellini, L.; Cohen, M.M. Ubiquitination of ERMES components by the E3 ligase Rsp5 is involved in mitophagy. Autophagy 2017, 13, 114–132. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Jiang, L.; Miao, H.; Lv, Y.; Zhang, Q.; Zhou, M.M.; Duan, W.; Huang, Y.; Wei, X. Autophagy regulation of ATG13 and ATG27 on biofilm formation and antifungal resistance in Candida albicans. Biofouling 2022, 38, 926–939. [Google Scholar] [CrossRef]
- Yen, W.L.; Klionsky, D.J. Atg27 is a Second Transmembrane Cycling Protein. Autophagy 2007, 3, 254–256. [Google Scholar] [CrossRef]
- Figueiredo, J.; Silva, M.S.; Figueiredo, A. Subtilisin-like proteases in plant defence: The past, the present and beyond. Mol. Plant Pathol. 2018, 19, 1017–1028. [Google Scholar] [CrossRef]
- Mao, K.; Klionsky, D.J. MAPKs regulate mitophagy in Saccharomyces cerevisiae. Autophagy 2011, 7, 1564–1565. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zheng, J.; Chen, S.; Meng, F.; Ning, J.; Lan, S.S. Oleandrin, a cardiac glycoside, induces immunogenic cell death via the PERK/elF2α/ATF4/CHOP pathway in breast cancer. Cell Death Dis. 2021, 12, 314. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yang, Z.; Liu, X.; Mao, K.; Nair, U.; Klionsky, D.J. Phosphatidylinositol 4-Kinases Are Required for Autophagic Membrane Trafficking*. J. Biol. Chem. 2012, 287, 37964–37972. [Google Scholar] [CrossRef]
- Korbecki, J.; Bobiński, R.; Dutka, M. Self-regulation of the inflammatory response by peroxisome proliferator-activated receptors. Inflamm. Res. 2019, 68, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Nordgren, M.; Fransen, M. Peroxisomal metabolism and oxidative stress. Biochimie 2014, 98, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Tripathi, D.N.; Jing, J.; Alexander, A.; Kim, J.; Powell, R.T.; Dere, R.; Tait-Mulder, J.; Lee, J.; Paull, T.T.; et al. ATM Functions at the Peroxisome to Induce Pexophagy in Response to ROS. Nat. Cell Biol. 2015, 17, 1259–1269. [Google Scholar] [CrossRef]
- Kim, J.A. Peroxisome Metabolism in Cancer. Cells 2020, 9, 17692. [Google Scholar] [CrossRef]
- Dong, P.T.; Jusuf, S.; Hui, J.; Zhan, Y.; Zhu, Y.; Liu, G.Y.; Cheng, J. Photoinactivation of catalase sensitizes a wide range of bacteria to ROS-producing agents and immune cells. JCI Insight 2021, 7, e153079. [Google Scholar] [CrossRef]
- Huang, Z.; Lu, J.; Liu, R.; Wang, P.S.; Hu, Y.; Fang, A.; Yang, Y.; Ling, Q.; Wei, B.C.; Yu, Y. SsCat2 encodes a catalase that is critical for the antioxidant response, QoI fungicide sensitivity, and pathogenicity of Sclerotinia sclerotiorum. Fungal Genet. Biol. 2021, 149, 103530. [Google Scholar] [CrossRef]
- Li, X.; Wu, X. Soluble epoxide hydrolase (Ephx2) silencing attenuates the hydrogen peroxide-induced oxidative damage in IEC-6 cells. Arch. Med Sci. 2019, 17, 1075–1086. [Google Scholar] [CrossRef]
- Appelqvist, H.; Wäster, P.; Kågedal, K.; Öllinger, K. The lysosome: From waste bag to potential therapeutic target. J. Mol. Cell Biol. 2013, 5, 214–226. [Google Scholar] [CrossRef]
- Feng, Y.; He, D.; Yao, Z.; Klionsky, D.J. The machinery of macroautophagy. Cell Res. 2013, 24, 24–41. [Google Scholar] [CrossRef]
- Liu, B.; Du, H.; Rutkowski, R.; Gartner, A.; Wang, X. LAAT-1 Is the Lysosomal Lysine/Arginine Transporter That Maintains Amino Acid Homeostasis. Science 2012, 337, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Adar, Y.Y.; Stark, M.; Bram, E.; Bram, E.; Nowak-Sliwinska, P.; Nowak-Sliwinska, P.; van den Bergh, H.; Szewczyk, G.M.; Sarna, T.J.; Składanowski, A.; et al. Imidazoacridinone-dependent lysosomal photodestruction: A pharmacological Trojan horse approach to eradicate multidrug-resistant cancers. Cell Death Dis. 2012, 3, e293. [Google Scholar] [CrossRef] [PubMed]
- Gotink, K.J.; Broxterman, H.J.; Labots, M.; de Haas, R.R.; Dekker, H.; Honeywell, R.J.; Rudek, M.A.; Beerepoot, L.V.; Musters, R.J.P.; Jansen, G.; et al. Lysosomal Sequestration of Sunitinib: A Novel Mechanism of Drug Resistance. Clin. Cancer Res. 2011, 17, 7337–7346. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, A.M.; Krise, J.P. Lysosomal sequestration of amine-containing drugs: Analysis and therapeutic implications. J. Pharm. Sci. 2007, 96, 729–746. [Google Scholar] [CrossRef] [PubMed]
- Zhitomirsky, B.; Assaraf, Y.G. Lysosomal sequestration of hydrophobic weak base chemotherapeutics triggers lysosomal biogenesis and lysosome-dependent cancer multidrug resistance. Oncotarget 2014, 6, 1143–1156. [Google Scholar] [CrossRef] [PubMed]
- Kazmi, F.; Hensley, T.; Pope, C.; Funk, R.S.; Loewen, G.J.; Buckley, D.B. Lysosomal sequestration (trapping) of lipophilic amine (Cationic amphiphilic) drugs in immortalized human hepatocytes (Fa2N-4 cells). Drug Metab. Dispos. 2013, 41, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Groth-Pedersen, L.; Ostenfeld, M.S.; Høyer-Hansen, M.H.; Nylandsted, J.; Jäättelä, M. Vincristine induces dramatic lysosomal changes and sensitizes cancer cells to lysosome-destabilizing siramesine. Cancer Res. 2007, 67, 2217–2225. [Google Scholar] [CrossRef]
- Herlevsen, M.C.; Oxford, G.; Owens, C.; Conaway, M.R.; Theodorescu, D. Depletion of major vault protein increases doxorubicin sensitivity and nuclear accumulation and disrupts its sequestration in lysosomes. Mol. Cancer Ther. 2007, 6, 1804–1813. [Google Scholar] [CrossRef]
- Smith, P.J.; Sykes, H.R.; Fox, M.E.; Furlong, I.J. Subcellular distribution of the anticancer drug mitoxantrone in human and drug-resistant murine cells analyzed by flow cytometry and confocal microscopy and its relationship to the induction of DNA damage. Cancer Res. 1992, 52, 4000–4008. [Google Scholar] [PubMed]
- Doherty, G.J.; McMahon, H.T. Mechanisms of endocytosis. Annu. Rev. Biochem. 2009, 78, 857–902. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Li, R.; Pan, J.; Ding, Z.; Lin, J. Endocytosis and its regulation in plants. Trends Plant Sci. 2015, 20, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Rauch, C.; Pluen, A. Multi drug resistance-dependent “vacuum cleaner” functionality potentially driven by the interactions between endocytosis, drug size and Pgp-like transporters surface density. Eur. Biophys. J. 2007, 36, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.X.; Meng, H.; Wang, Y.; He, H.; Meng, J.; Lu, F.J.; Wang, P.C.; Zhao, Y.; Gao, X.; Sun, B.; et al. Metallofullerene nanoparticles circumvent tumor resistance to cisplatin by reactivating endocytosis. Proc. Natl. Acad. Sci. USA 2010, 107, 7449–7454. [Google Scholar] [CrossRef] [PubMed]
- Sung, M.; Tan, X.; Lu, B.; Golas, J.; Hosselet, C.; Wang, F.; Tylaska, L.; King, L.; Zhou, D.; Dushin, R.A. Caveolae-mediated endocytosis as a novel mechanism of resistance to trastuzumab emtansine (T-DM1). Mol. Cancer Ther. 2017, 17, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Funakoshi, Y.; Hasegawa, H.; Kanaho, Y. Regulation of PIP5K activity by Arf6 and its physiological significance. J. Cell. Physiol. 2011, 226, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Malerød, L.; Stuffers, S.; Brech, A.; Stenmark, H.A. Vps22/EAP30 in ESCRT-II Mediates Endosomal Sorting of Growth Factor and Chemokine Receptors Destined for Lysosomal Degradation. Traffic 2007, 8, 1617–1629. [Google Scholar] [CrossRef]
- Purrington, C.B. Costs of resistance. Curr. Opin. Plant Biol. 2000, 3, 305–308. [Google Scholar] [CrossRef]
- Kemble, H.E.; Eisenhauer, C.; Couce, A.; Chapron, A.; Magnan, M.; Gautier, G.; Nagard, H.L.; Nghe, P.; Tenaillon, O. Flux, toxicity, and expression costs generate complex genetic interactions in a metabolic pathway. Sci. Adv. 2020, 6, eabb2236. [Google Scholar] [CrossRef]
- Kushalappa, A.C.; Hegde, N.G.; Yogendra, K. Metabolic pathway genes for editing to enhance multiple disease resistance in plants. J. Plant Res. 2022, 135, 705–722. [Google Scholar] [CrossRef] [PubMed]
- Nagiec, M.M.; Baltisberger, J.A.; Wells, G.B.; Lester, R.L.; Dickson, R.C. The LCB2 gene of Saccharomyces and the related LCB1 gene encode subunits of serine palmitoyltransferase, the initial enzyme in sphingolipid synthesis. Proc. Natl. Acad. Sci. USA 1994, 91, 7899–7902. [Google Scholar] [CrossRef] [PubMed]
- Neumetzler, L.; Humphrey, T.V.; Lumba, S.; Snyder, S.I.; Yeats, T.H.; Usadel, B.; Vasilevski, A.; Patel, J.G.; Rose, J.K.C.; Persson, S.; et al. The FRIABLE1 Gene Product Affects Cell Adhesion in Arabidopsis. PLoS ONE 2012, 7, e42914. [Google Scholar] [CrossRef] [PubMed]











| Sample | Raw Reads | Clean Reads | Clean Bases | Q20 (%) | Q30 (%) | GC (%) |
|---|---|---|---|---|---|---|
| Cc260-Tri-1 | 52,805,726 | 52,025,868 | 7.8 G | 97.91% | 94.31% | 57.21% |
| Cc260-Tri-2 | 50,946,058 | 50,132,632 | 7.52 G | 97.94% | 94.44% | 57.36% |
| Cc260-Tri-3 | 51,392,366 | 50,619,174 | 7.59 G | 98.11% | 94.91% | 57.24% |
| Cc241-Tri-1 | 54,353,210 | 53,409,354 | 8.01 G | 97.80% | 94.18% | 56.82% |
| Cc241-Tri-2 | 56,572,142 | 55,709,416 | 8.36 G | 97.94% | 94.50% | 56.93% |
| Cc241-Tri-3 | 67,258,428 | 66,177,436 | 9.93 G | 97.93% | 94.51% | 56.73% |
| Cc66-Tri-1 | 58,396,696 | 57,283,728 | 8.59 G | 97.88% | 94.39% | 56.96% |
| Cc66-Tri-2 | 63,712,892 | 62,669,002 | 9.4 G | 97.69% | 93.88% | 57.04% |
| Cc66-Tri-3 | 54,820,830 | 53,534,274 | 8.03 G | 97.95% | 94.57% | 57.04% |
| Cc71-Tri-1 | 61,482,532 | 60,734,460 | 9.11 G | 97.89% | 94.39% | 56.64% |
| Cc71-Tri-2 | 58,387,148 | 57,468,612 | 8.62 G | 97.98% | 94.67% | 56.80% |
| Cc71-Tri-3 | 53,542,268 | 52,693,836 | 7.9 G | 97.99% | 94.68% | 56.89% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, B.; Zhou, R.; Zhu, G.; Xie, X.; Chai, A.; Li, L.; Fan, T.; Li, B.; Shi, Y. Transcriptome Analysis Reveals the Involvement of Mitophagy and Peroxisome in the Resistance to QoIs in Corynespora cassiicola. Microorganisms 2023, 11, 2849. https://doi.org/10.3390/microorganisms11122849
Sun B, Zhou R, Zhu G, Xie X, Chai A, Li L, Fan T, Li B, Shi Y. Transcriptome Analysis Reveals the Involvement of Mitophagy and Peroxisome in the Resistance to QoIs in Corynespora cassiicola. Microorganisms. 2023; 11(12):2849. https://doi.org/10.3390/microorganisms11122849
Chicago/Turabian StyleSun, Bingxue, Rongjia Zhou, Guangxue Zhu, Xuewen Xie, Ali Chai, Lei Li, Tengfei Fan, Baoju Li, and Yanxia Shi. 2023. "Transcriptome Analysis Reveals the Involvement of Mitophagy and Peroxisome in the Resistance to QoIs in Corynespora cassiicola" Microorganisms 11, no. 12: 2849. https://doi.org/10.3390/microorganisms11122849
APA StyleSun, B., Zhou, R., Zhu, G., Xie, X., Chai, A., Li, L., Fan, T., Li, B., & Shi, Y. (2023). Transcriptome Analysis Reveals the Involvement of Mitophagy and Peroxisome in the Resistance to QoIs in Corynespora cassiicola. Microorganisms, 11(12), 2849. https://doi.org/10.3390/microorganisms11122849