
Effects of a Microbial Restoration Substrate on Plant Growth and Rhizosphere Microbial Community in a Continuous Cropping Poplar


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Design
2.2. Collecting Samples and Counting Culturable Microorganisms
2.3. Edaphic Properties Determination
2.4. DNA Isolation and PCR Amplification
2.5. Real-Time (q)PCR
2.6. Illumina MiSeq Sequencing
2.7. Data Processing for Sequencing
2.8. Statistical Analyses
3. Results
3.1. DBH and Plant Height Changes in Poplar Trees
3.2. Variations in Edaphic Characteristics between March 2018 and October 2020
3.3. Microbial Quantities in Poplar Rhizosphere Soils
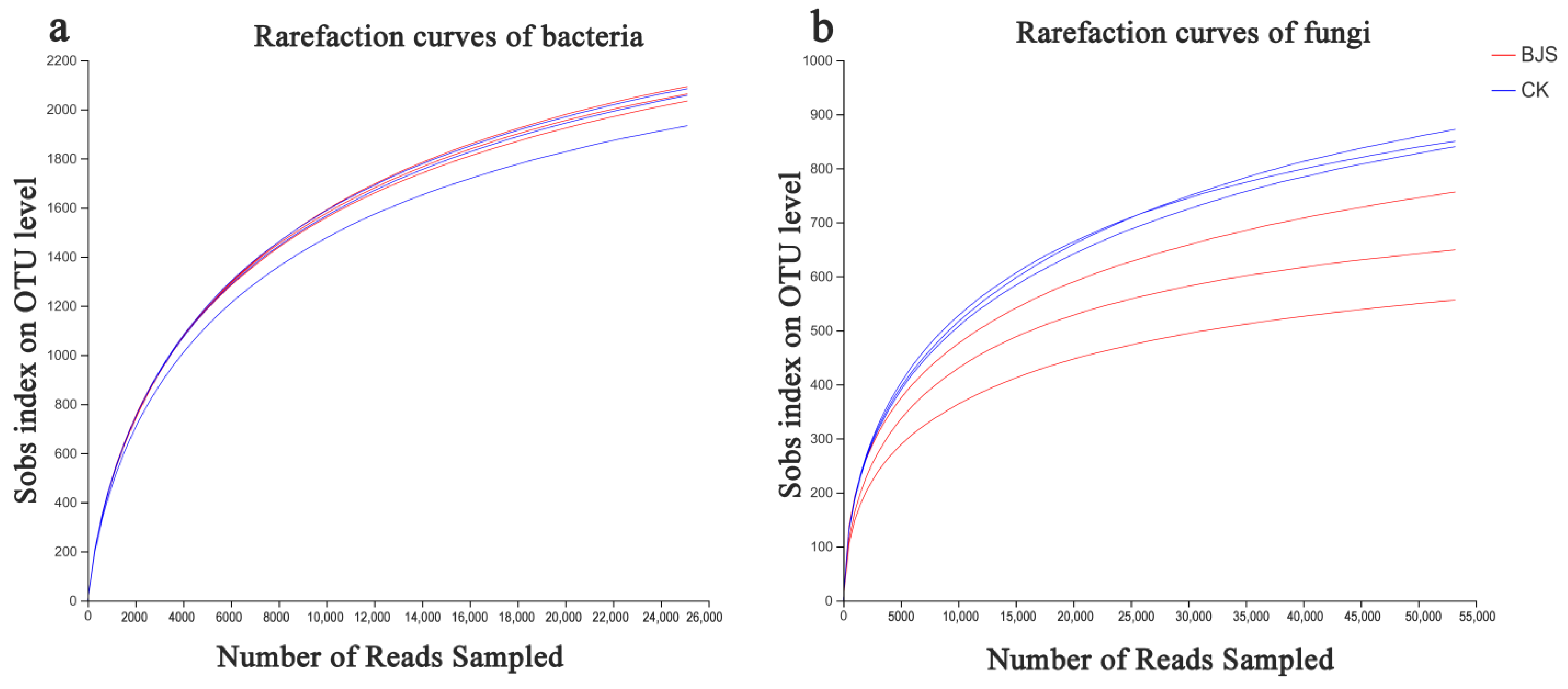
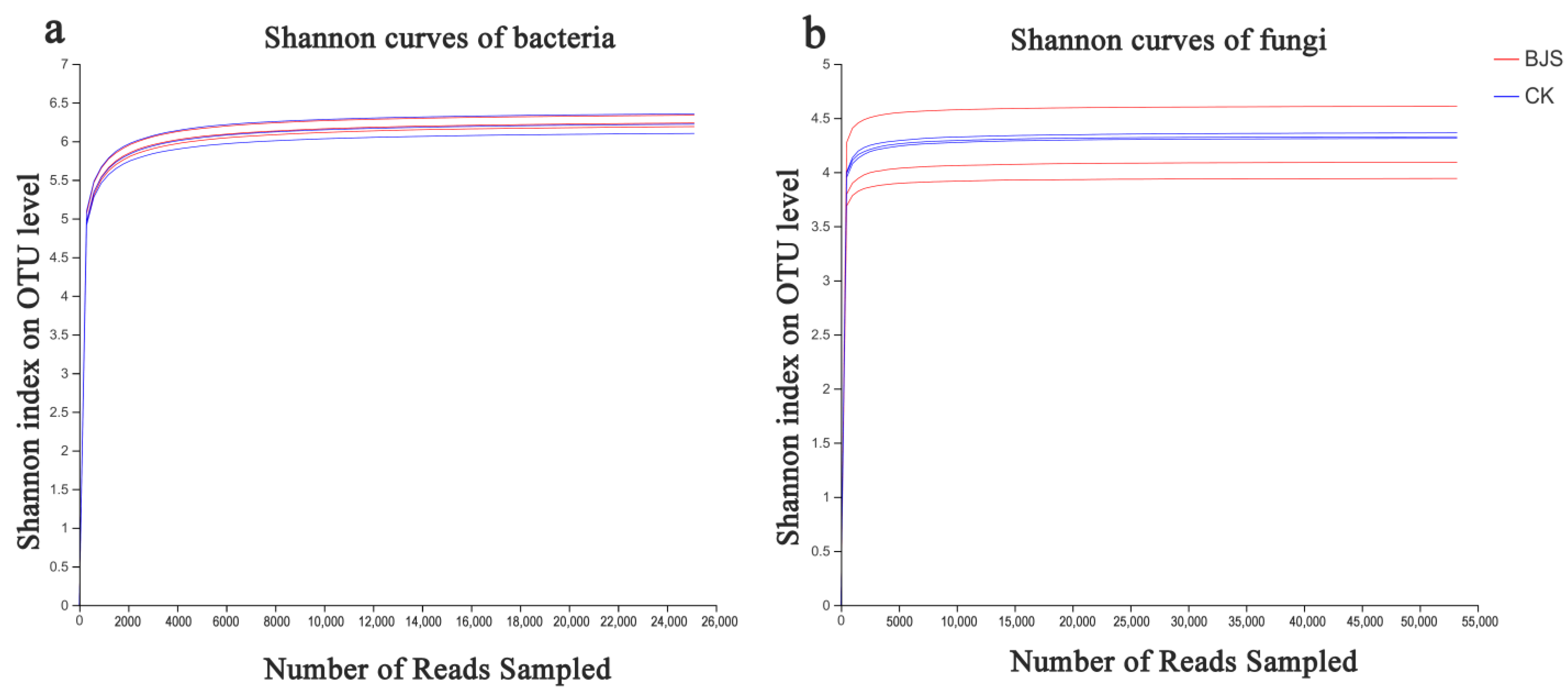
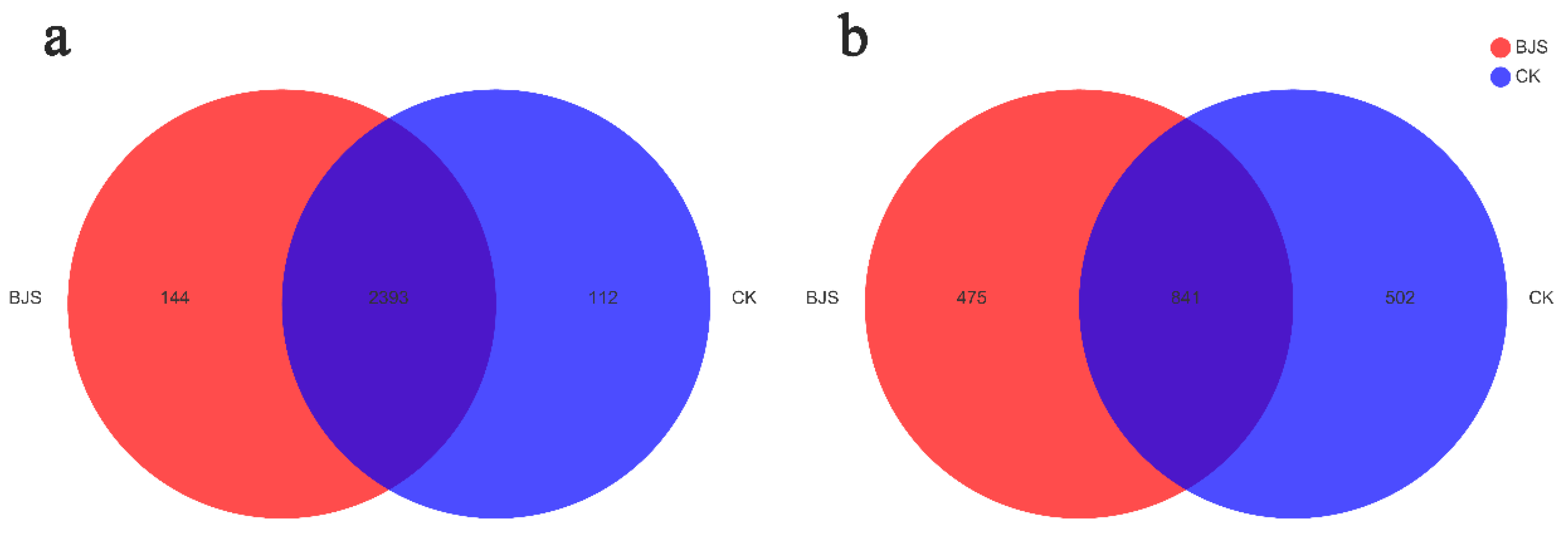
3.4. Estimates of Microbial Community Diversity and Species Richness
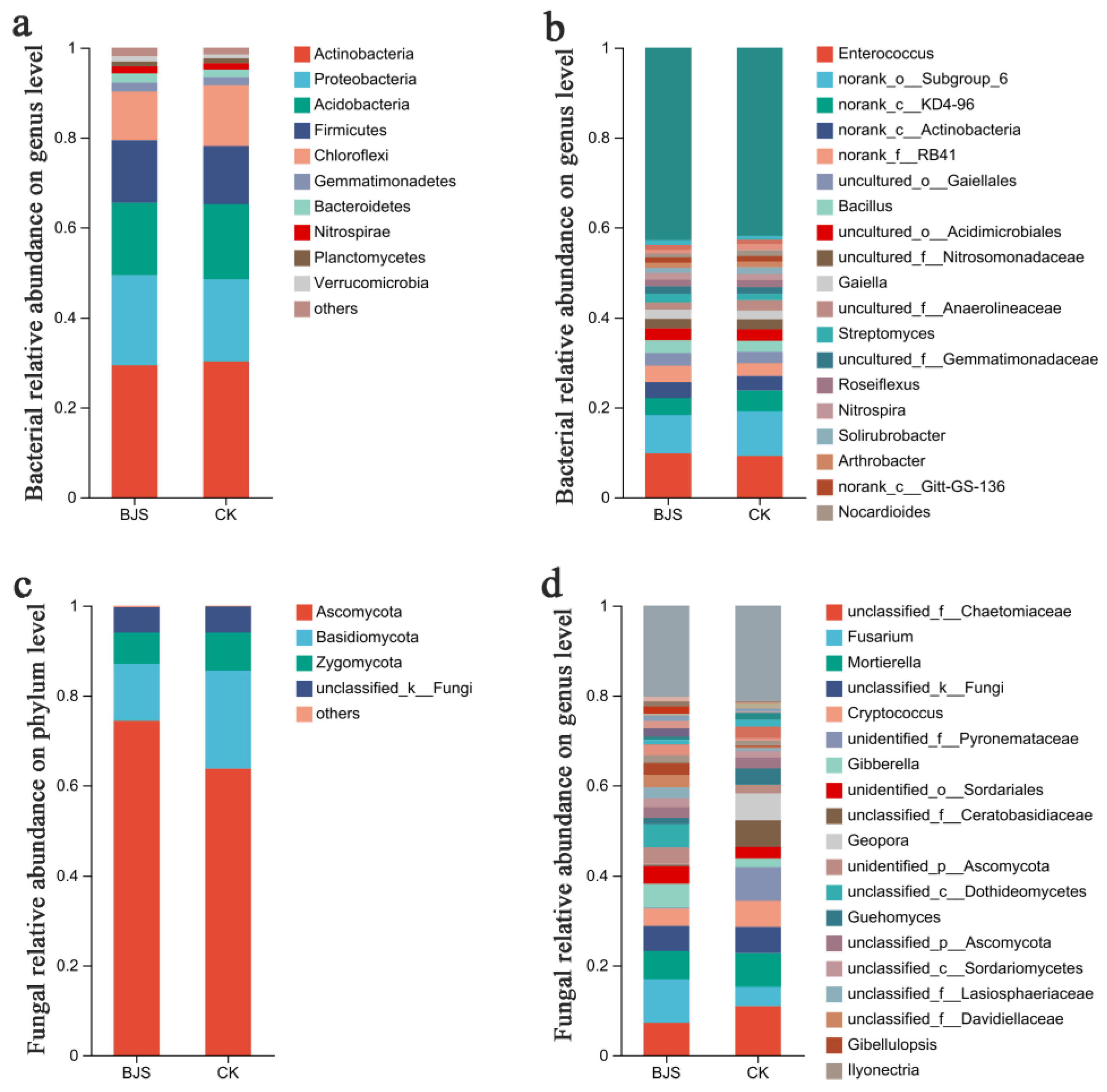
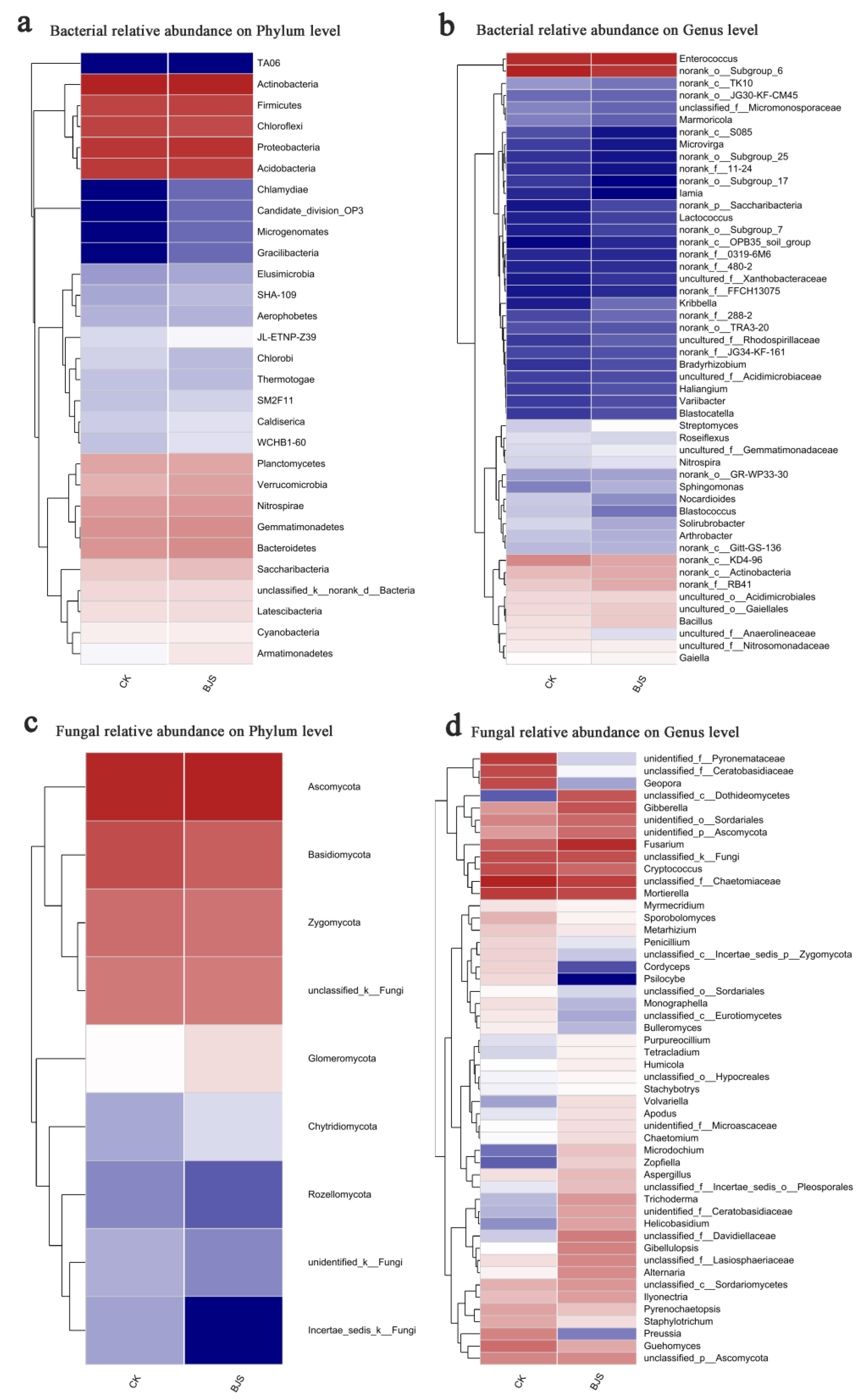
3.5. Composition and Structure of Communities Affected by BJS-1-3
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, C.; Lu, L.; Guo, Y.; Xu, H.; Li, R. Efficient Agrobacterium-Mediated Transformation of the Commercial Hybrid Poplar Populus alba × Populus glandulosa Uyeki. Int. J. Mol. Sci. 2019, 20, 2594. [Google Scholar] [CrossRef] [Green Version]
- Fortier, J.; Truax, B.; Gagnon, D.; Lambert, F. Abiotic and biotic factors controlling fine root biomass, carbon and nutrients in closed-canopy hybrid poplar stands on post-agricultural land. Sci. Rep. 2019, 9, 6296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, G. Populus: Arabidopsis for Forestry. Do We Need a Model Tree? Ann. Bot. 2002, 90, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Lv, J.; Zhao, Q.; Dong, Y.; Dong, K. Cinnamic Acid Increased the Incidence of Fusarium Wilt by Increasing the Pathogenicity of Fusarium oxysporum and Reducing the Physiological and Biochemical Resistance of Faba Bean, Which was Alleviated by Intercropping with Wheat. Front. Plant Sci. 2020, 11, 608389. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhao, S.; Zhao, J.; Zhang, K.; Jiang, J.; Guan, Z.; Chen, S.; Chen, F.; Fang, W. Deep tillage combined with biofertilizer following soil fumigation improved chrysanthemum growth by regulating the soil microbiome. MicrobiologyOpen 2020, 9, e1045. [Google Scholar] [CrossRef] [Green Version]
- Yao, Q.; Xu, Y.; Liu, X.; Liu, J.; Huang, X.; Yang, W.; Yang, Z.; Lan, L.; Zhou, J.; Wang, G. Dynamics of soil properties and fungal community structure in continuous-cropped alfalfa fields in Northeast China. PeerJ 2019, 7, e7127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Chen, Z.; Qian, J.; Wei, F.; Zhang, G.; Wang, Y.; Wei, G.; Hu, Z.; Dong, L.; Chen, S. Composition and function of rhizosphere microbiome of Panax notoginseng with discrepant yields. Chin. Med. 2020, 15, 85. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, X.; Liu, T.; Wang, H.; Yang, Y.; Chen, X.; Zhu, S. Analysis of bacterial and fungal communities in continuous-cropping ramie (Boehmeria nivea L. Gaud) fields in different areas in China. Sci. Rep. 2020, 10, 3264. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Zhang, W.; Zhang, L.; Liu, X.; Wei, H. Supramolecular Structure and Functional Analysis of the Type III Secretion System in Pseudomonas fluorescens 2P24. Front. Plant Sci. 2015, 6, 1190. [Google Scholar] [CrossRef] [Green Version]
- Ling, L.; Han, X.; Li, X.; Zhang, X.; Wang, H.; Zhang, L.; Cao, P.; Wu, Y.; Wang, X.; Zhao, J.; et al. A Streptomyces sp. NEAU-HV9: Isolation, Identification, and Potential as a Biocontrol Agent against Ralstonia solanacearum of Tomato Plants. Microorganisms 2020, 8, 351. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, Z.; Zhu, Y.; Wang, J.; Liu, B. Effects of a microbial restoration substrate on plant growth and rhizosphere bacterial community in a continuous tomato cropping greenhouse. Sci. Rep. 2020, 10, 13729. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, S.; Li, N.; Chen, H.; Jia, H.; Song, X.; Liu, G.; Ni, C.; Wang, Z.; Shao, H.; et al. Metagenomic insights into effects of wheat straw compost fertiliser application on microbial community composition and function in tobacco rhizosphere soil. Sci. Rep. 2019, 9, 6168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhu, S.; Liu, T.; Guo, B.; Li, F.; Bai, X. Identification of the rhizospheric microbe and metabolites that led by the continuous cropping of ramie (Boehmeria nivea L. Gaud). Sci. Rep. 2020, 10, 20408. [Google Scholar] [CrossRef] [PubMed]
- Arrebola, E.; Tienda, S.; Vida, C.; de Vicente, A.; Cazorla, F.M. Fitness Features Involved in the Biocontrol Interaction of Pseudomonas chlororaphis with Host Plants: The Case Study of PcPCL1606. Front. Microbiol. 2019, 10, 719. [Google Scholar] [CrossRef] [Green Version]
- Kuźniar, A.; Włodarczyk, K.; Grządziel, J.; Woźniak, M.; Furtak, K.; Gałązka, A.; Dziadczyk, E.; Skórzyńska-Polit, E.; Wolińska, A. New Insight into the Composition of Wheat Seed Microbiota. Int. J. Mol. Sci. 2020, 21, 4634. [Google Scholar] [CrossRef]
- Huang, K.; Jiang, Q.; Liu, L.; Zhang, S.; Liu, C.; Chen, H.; Ding, W.; Zhang, Y. Exploring the key microbial changes in the rhizosphere that affect the occurrence of tobacco root-knot nematodes. AMB Express 2020, 10, 72. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, W.; Liu, W.; Ahammed, G.J.; Wen, W.; Guo, S.; Shu, S.; Sun, J. Auxin is involved in arbuscular mycorrhizal fungi-promoted tomato growth and NADP-malic enzymes expression in continuous cropping substrates. BMC Plant Biol. 2021, 21, 48. [Google Scholar] [CrossRef]
- Shao, J.; Li, S.; Zhang, N.; Cui, X.; Zhou, X.; Zhang, G.; Shen, Q.; Zhang, R. Analysis and cloning of the synthetic pathway of the phytohormone indole-3-acetic acid in the plant-beneficial Bacillus amyloliquefaciens SQR9. Microb. Cell Factories 2015, 14, 130. [Google Scholar] [CrossRef] [Green Version]
- Belbahri, L.; Bouket, A.C.; Rekik, I.; Alenezi, F.N.; Vallat, A.; Luptakova, L.; Petrovova, E.; Oszako, T.; Cherrad, S.; Vacher, S.; et al. Comparative Genomics of Bacillus amyloliquefaciens Strains Reveals a Core Genome with Traits for Habitat Adaptation and a Secondary Metabolites Rich Accessory Genome. Front. Microbiol. 2017, 8, 1438. [Google Scholar] [CrossRef] [Green Version]
- Zubair, M.; Hanif, A.; Farzand, A.; Sheikh, T.M.M.; Khan, A.R.; Suleman, M.; Ayaz, M.; Gao, X. Genetic Screening and Expression Analysis of Psychrophilic Bacillus spp. Reveal Their Potential to Alleviate Cold Stress and Modulate Phytohormones in Wheat. Microorganisms 2019, 7, 337. [Google Scholar] [CrossRef]
- Moon, J.H.; Won, S.J.; Maung, C.E.H.; Choi, J.H.; Choi, S.I.; Ajuna, H.B.; Ahn, Y.S. Bacillus velezensis CE 100 Inhibits Root Rot Diseases (Phytophthora spp.) and Promotes Growth of Japanese Cypress (Chamaecyparis obtusa Endlicher) Seedlings. Microorganisms 2021, 9, 821. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Wei, Z.; Guan, Z.; Cai, Y.; Liao, X. Antifungal Activity of Isolated Bacillus amyloliquefaciens SYBC H47 for the Biocontrol of Peach Gummosis. PLoS ONE 2016, 11, e0162125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kačániová, M.; Terentjeva, M.; Galovičová, L.; Ivanišová, E.; Štefániková, J.; Valková, V.; Borotová, P.; Kowalczewski, P.Ł.; Kunová, S.; Felšöciová, S.; et al. Biological Activity and Antibiofilm Molecular Profile of Citrus aurantium Essential Oil and its Application in a Food Model. Molecules 2020, 25, 3956. [Google Scholar] [CrossRef] [PubMed]
- Scholz, R.; Vater, J.; Budiharjo, A.; Wang, Z.; He, Y.; Dietel, K.; Schwecke, T.; Herfort, S.; Lasch, P.; Borriss, R. Amylocyclicin, a Novel Circular Bacteriocin Produced by Bacillus amyloliquefaciens FZB42. J. Bacteriol. 2014, 196, 1842–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elarabi, N.I.; Abdelhadi, A.A.; Ahmed, R.H.; Saleh, I.; Arif, I.A.; Osman, G.; Ahmed, D.S. Bacillus aryabhattai FACU: A promising bacterial strain capable of manipulate the glyphosate herbicide residues. Saudi J. Biol. Sci. 2020, 27, 2207–2214. [Google Scholar] [CrossRef]
- Huang, M.; Bulut, A.; Shrestha, B.; Matera, C.; Grundler, F.M.W.; Schleker, A.S.S. Bacillus firmus I-1582 promotes plant growth and impairs infection and development of the cyst nematode Heterodera schachtii over two generations. Sci. Rep. 2021, 11, 14114. [Google Scholar] [CrossRef]
- Bonito, G.; Hameed, K.; Ventura, R.; Krishnan, J.; Schadt, C.W.; Vilgalys, R. Isolating a functionally relevant guild of fungi from the root microbiome of Populus. Fungal Ecol. 2016, 22, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Walkly, A. An examination of the Degtjareff method for determining soil organic matter and a proposed modification to the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Olsen, S.R. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Bibli 1954, 939, 18–19. [Google Scholar]
- Wang, P.; Chen, B.; Zhang, H. High throughput sequencing analysis of bacterial communities in soils of a typical Po-yang Lake wetland. Acta Ecol. Sin. 2017, 37, 1650–1658. [Google Scholar]
- Li, X.; Wang, Y.; Lin, J.; Liu, L.; Huang, Z. Analysis of Microbial Diversity in the Fermented Grains of Maotai-flavor Liquor Using High-throughput Sequencing. J. Fujian Norm. Univ. 2017, 33, 9. [Google Scholar]
- Hao, Y.T.; Wu, S.T.; Xiong, F.; Tran, N.T.; Jakovlić, I.; Zou, H.; Li, W.X.; Wang, G.T. Succession and Fermentation Products of Grass Carp (Ctenopharyngodon idellus) Hindgut Microbiota in Response to an Extreme Dietary Shift. Front. Microbiol. 2017, 8, 1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, M.; Qiu, T.; Sun, Y.; Wang, X. The abundance and diversity of antibiotic resistance genes in the atmospheric environment of com-posting plants. Environ. Int. 2018, 116, 229. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Amato, K.R.; Yeoman, C.J.; Kent, A.; Righini, N.; Carbonero, F.; Estrada, A.; Gaskins, H.R.; Stumpf, R.M.; Yildirim, S.; Torralba, M.; et al. Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes. ISME J. 2013, 7, 1344–1353. [Google Scholar] [CrossRef]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi—Recent updates and future perspectives. New Phytol. 2010, 186, 281–285. [Google Scholar] [CrossRef]
- Lu, Y.; Gao, P.; Wang, Y.; Li, W.; Cui, X.; Zhou, J.; Peng, F.; Dai, L. Earthworm activity optimized the rhizosphere bacterial community structure and further alleviated the yield loss in continuous cropping lily (Lilium lancifolium Thunb.). Sci. Rep. 2021, 11, 20840. [Google Scholar] [CrossRef]
- Peng, F.; Zhang, M.-Y.; Hou, S.-Y.; Chen, J.; Wu, Y.-Y.; Zhang, Y.-X. Insights into Streptomyces spp. isolated from the rhizospheric soil of Panax notoginseng: Isolation, antimicrobial activity and biosynthetic potential for polyketides and non-ribosomal peptides. BMC Microbiol. 2020, 20, 143. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, D.; Yang, Y.; Pan, Y.; Zhao, D.; Zhu, J.; Zhang, L.; Yang, Z. Dissecting the effect of continuous cropping of potato on soil bacterial communities as revealed by high-throughput sequencing. PLoS ONE 2020, 15, e0233356. [Google Scholar] [CrossRef]
- Wolfgang, A.; Zachow, C.; Müller, H.; Grand, A.; Temme, N.; Tilcher, R.; Berg, G. Understanding the Impact of Cultivar, Seed Origin and Substrate on Bacterial Diversity of the Sugar Beet Rhizosphere and Suppression of Soil-Borne Pathogens. Front. Plant Sci. 2020, 11, 560869. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Ren, Z.-H.; Zu, X.; Yu, X.-Y.; Zhu, H.-J.; Li, X.-J.; Zhong, J.; Liu, E.-M. Efficacy of Plant Growth-Promoting Bacteria Bacillus cereus YN917 for Biocontrol of Rice Blast. Front. Microbiol. 2021, 12, 684888. [Google Scholar] [CrossRef] [PubMed]
- Jiao, S.; Peng, Z.; Qi, J.; Gao, J.; Wei, G. Linking Bacterial-Fungal Relationships to Microbial Diversity and Soil Nutrient Cycling. mSystems 2021, 6, e01052-20. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Xiong, X.; Tan, L.; Deng, Y.; Du, X.; Yang, X.; Hu, Q. Soil microbial community assembly and stability are associated with potato (Solanum tuberosum L.) fitness under continuous cropping regime. Front. Plant Sci. 2022, 13, 1000045. [Google Scholar] [CrossRef] [PubMed]
- Aguirre de Cárcer, D. A conceptual framework for the phylogenetically constrained assembly of microbial communities. Microbiome 2019, 7, 142. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-T.; Whitman, W.B.; Coleman, D.C.; Chiu, C.-Y. Effects of Reforestation on the Structure and Diversity of Bacterial Communities in Subtropical Low Mountain Forest Soils. Front. Microbiol. 2018, 9, 1968. [Google Scholar] [CrossRef] [Green Version]
- Agnolucci, M.; Palla, M.; Cristani, C.; Cavallo, N.; Giovannetti, M.; De Angelis, M.; Gobbetti, M.; Minervini, F. Beneficial Plant Microorganisms Affect the Endophytic Bacterial Communities of Durum Wheat Roots as Detected by Different Molecular Approaches. Front. Microbiol. 2019, 10, 2500. [Google Scholar] [CrossRef] [Green Version]
- Velazquez-Martinez, V.; Valles-Rosales, D.; Rodriguez-Uribe, L.; Holguin, O.; Quintero-Quiroz, J.; Reyes-Jaquez, D.; Rodriguez-Borbon, M.I.; Villagrán-Villegas, L.Y.; Delgado, E. Antimicrobial, Shelf-Life Stability, and Effect of Maltodextrin and Gum Arabic on the Encapsulation Efficiency of Sugarcane Bagasse Bioactive Compounds. Foods 2021, 10, 116. [Google Scholar] [CrossRef]
- Ren, M.; Zhang, Z.; Wang, X.; Zhou, Z.; Chen, D.; Zeng, H.; Zhao, S.; Chen, L.; Hu, Y.; Zhang, C.; et al. Diversity and Contributions to Nitrogen Cycling and Carbon Fixation of Soil Salinity Shaped Microbial Communities in Tarim Basin. Front. Microbiol. 2018, 9, 431. [Google Scholar] [CrossRef] [Green Version]
- Challacombe, J.F.; Hesse, C.N.; Bramer, L.M.; McCue, L.A.; Lipton, M.; Purvine, S.; Nicora, C.; Gallegos-Graves, L.V.; Porras-Alfaro, A.; Kuske, C.R. Genomes and secretomes of Ascomycota fungi reveal diverse functions in plant biomass decomposition and pathogenesis. BMC Genom. 2019, 20, 976. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Lin, L.; Zhao, T.; Yuan, Y.; Tai, Y.; Ye, A. Effects of Continuous Cropping of Lilium Lancifolium on Rhizosphere Soil Physical and Chemical Properties and Fungal Community Structure. Res. Sq. 2021, preprints. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, F. Dynamics of the diversity of fungal and Fusarium communities during continuous crop-ping of cucumber in the greenhouse. In Proceedings of the 4th International Symposium on Modern Agriculture and Rural Economic Development in Asia, Harbin, China, 7–9 July 2012. [Google Scholar]
- Zhang, Y.; Tian, L.; Yan, D.-H.; He, W. Genome-Wide Transcriptome Analysis Reveals the Comprehensive Response of Two Susceptible Poplar Sections to Marssonina brunnea Infection. Genes 2018, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Zhang, C.; Wang, Z.; Duan, T.; Yu, B.; Jia, X.; Pang, J.; Ma, L.; Wang, Y.; Nan, Z. Effects of Co-infection by Soil-borne Fungal Pathogens on Disease Responses among Diverse Alfalfa Varieties. Front. Microbiol. 2021, 12, 664385. [Google Scholar] [CrossRef] [PubMed]
- Erper, I.; Ozer, G.; Kalendar, R.; Avci, S.; Yildirim, E.; Alkan, M.; Turkkan, M. Genetic Diversity and Pathogenicity of Rhizoctonia spp. Isolates Associated with Red Cabbage in Samsun (Turkey). J. Fungi 2021, 7, 234. [Google Scholar] [CrossRef] [PubMed]
- Dutta, M.; Saha, A.; Moin, M.; Kirti, P.B. Genome-wide identification, expression and bioinformatic analyses of GRAS transcription factor genes in rice. Front. Plant Sci. 2021, 12, 777285. [Google Scholar] [CrossRef]
- Zeng, J.; Liu, J.; Lu, C.; Ou, X.; Luo, K.; Li, C.; He, M.; Zhang, H.; Yan, H. Intercropping with Turmeric or Ginger Reduce the Continuous Cropping Obstacles That Affect Pogostemon cablin (Patchouli). Front. Microbiol. 2020, 11, 579719. [Google Scholar] [CrossRef]
- Basbagci, G.; Dolar, F.S. Characterization and pathogenicity of Rhizoctonia solani AG-5 isolates obtained from chickpea in Turkey. J. Phytopathol. 2022, 170, 141–147. [Google Scholar] [CrossRef]
- Ramírez, V.; Martínez, J.; Bustillos-Cristales, M.D.R.; Catañeda-Antonio, D.; Munive, J.; Baez, A. Bacillus cereus MH778713 elicits tomato plant protection against Fusarium oxysporum. J. Appl. Microbiol. 2021, 132, 470–482. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | DBH (mm) in 2018/10 | DBH (mm) in 2020/10 | Plant Height (m) in 2018/10 | Plant Height (m) in 2020/10 |
|---|---|---|---|---|
| BJS-1-3 | 38.52 ± 4.13 a | 122.58 ± 8.73 a | 4.15 ± 0.28 a | 11.27 ± 0.56 a |
| CK | 35.23 ± 4.12 b | 111.13 ± 8.84 b | 3.99 ± 0.30 a | 10.73 ± 0.48 b |
| OC (g/kg) | AP (mg/kg) | AK (mg/kg) | TN (mg/kg) | pH | ||
|---|---|---|---|---|---|---|
| March 2018 | BJS-1-3 | 6.06 ± 0.05 c | 46.23 ± 0.13 b | 131.44 ± 0.82 b | 771.78 ± 3.26 b | 7.21 ± 0.01 b |
| CK | 6.05 ± 0.07 c | 35.63 ± 2.33 b | 132.28 ± 1.44 b | 765.82 ± 7.43 b | 7.23 ± 0.02 b | |
| October 2020 | BJS-1-3 | 7.14 ± 0.02 a | 43.96 ± 0.70 a | 186.29 ± 2.40 a | 854.93 ± 3.94 a | 7.40 ± 0.01 a |
| CK | 6.27 ± 0.04 b | 35.38 ± 2.77 b | 139.20 ± 2.03 b | 791.58 ± 3.77 b | 7.07 ± 0.04 c | |
| Treatment | Culturable Microbial Contents after the Two Treatments | Total Microbial Contents after the Two Treatments | ||
|---|---|---|---|---|
| Bacterial × 107 (cfu/g Soil) | Fungal × 106 (cfu/g Soil) | Bacterial × 107 (Copies/μL) | Fungal × 104 (Copies/μL) | |
| BJS-1-3 | 9.56 ± 0.21 a | 5.75 ± 0.31 b | 1.24 ± 0.23 a | 2.07 ± 0.08 b |
| CK | 0.96 ± 0.17 b | 9.25 ± 0.49 a | 0.71 ± 0.02 b | 2.95 ± 0.10 a |
| Sample | Cutoff | OTUs | ACE | Chao | Shannon | Simpson | Coverage | |
|---|---|---|---|---|---|---|---|---|
| Bacterial | BJS-1-3 | 0.03 | 2209 ± 51.36 a | 2491.02 ± 62.03 a | 2451.81 ± 88.31 a | 6.24 ± 0.06 a | 0.011 ± 0.003 a | 0.980661 |
| CK | 0.03 | 2106 ± 106.71 a | 2384.41 ± 82.60 a | 2402.16 ± 78.39 a | 6.20 ± 0.12 a | 0.012 ± 0.004 a | 0.980821 | |
| Fungal | BJS-1-3 | 0.03 | 793 ± 68.93 a | 775.30 ± 50.50 b | 793.31 ± 63.1 b | 3.97 ± 0.30 b | 0.057 ± 0.019 a | 0.997483 |
| CK | 0.03 | 901 ± 7.55 a | 1032.86 ± 57.48 a | 1022.94 ± 93.32 a | 4.34 ± 0.02 a | 0.040 ± 0.005 b | 0.996280 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sui, J.; Yang, J.; Li, C.; Zhang, L.; Hua, X. Effects of a Microbial Restoration Substrate on Plant Growth and Rhizosphere Microbial Community in a Continuous Cropping Poplar. Microorganisms 2023, 11, 486. https://doi.org/10.3390/microorganisms11020486
Sui J, Yang J, Li C, Zhang L, Hua X. Effects of a Microbial Restoration Substrate on Plant Growth and Rhizosphere Microbial Community in a Continuous Cropping Poplar. Microorganisms. 2023; 11(2):486. https://doi.org/10.3390/microorganisms11020486
Chicago/Turabian StyleSui, Junkang, Jiayi Yang, Chenyu Li, Lingxiao Zhang, and Xuewen Hua. 2023. "Effects of a Microbial Restoration Substrate on Plant Growth and Rhizosphere Microbial Community in a Continuous Cropping Poplar" Microorganisms 11, no. 2: 486. https://doi.org/10.3390/microorganisms11020486
APA StyleSui, J., Yang, J., Li, C., Zhang, L., & Hua, X. (2023). Effects of a Microbial Restoration Substrate on Plant Growth and Rhizosphere Microbial Community in a Continuous Cropping Poplar. Microorganisms, 11(2), 486. https://doi.org/10.3390/microorganisms11020486