
Molecular Phylogenetic Analysis of Paracoccidioides Species Complex Present in Paracoccidioidomycosis Patient Tissue Samples


 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
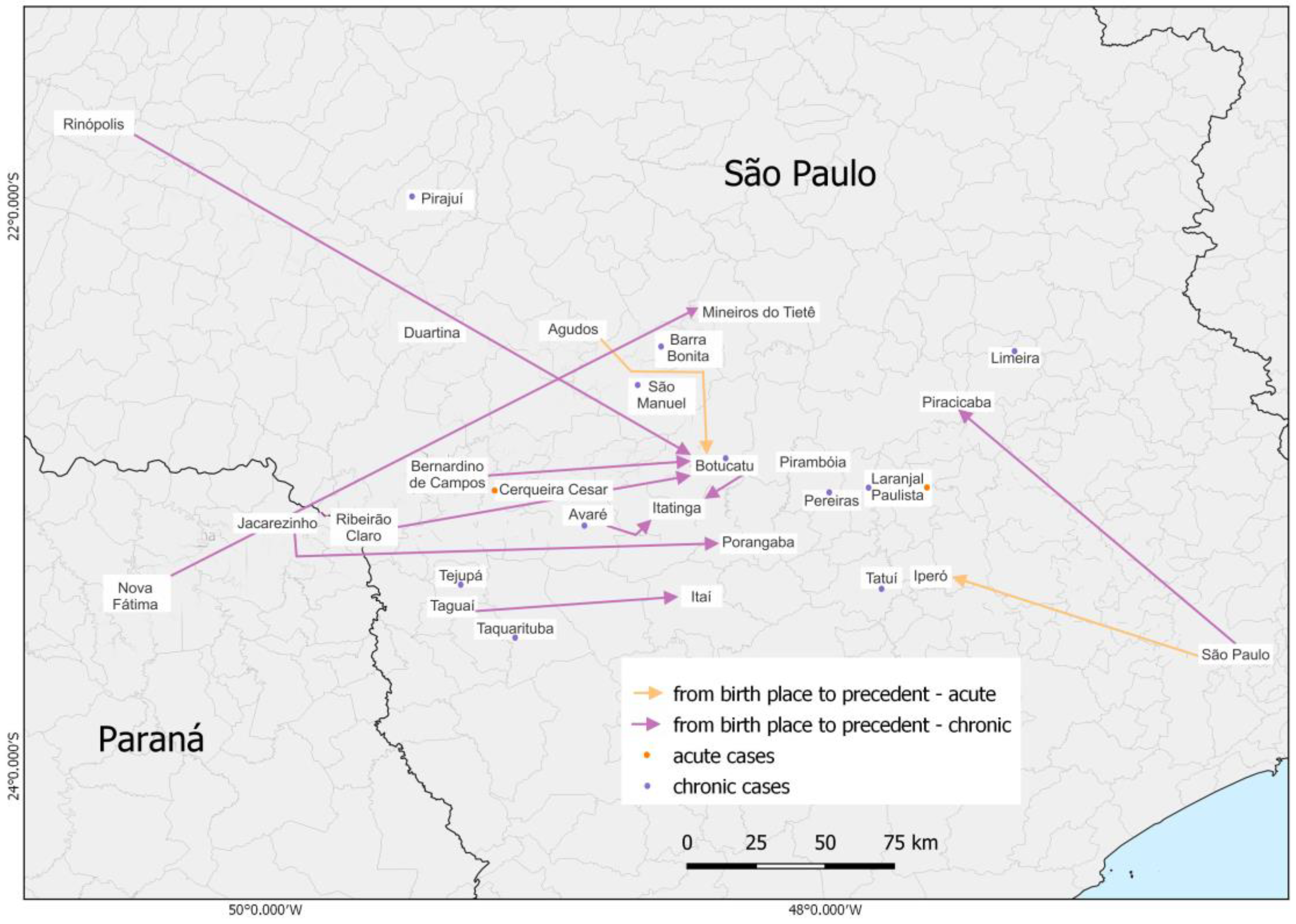
2.1. Study Setting, Data, and Sample Collection
2.2. DNA Extraction
2.3. PCR Amplification of Target Loci ITS, ARF, CHS2, and Agarose Gel Electrophoresis
2.4. Purification and Sequencing of Target Loci ITS, ARF, and CHS2
2.5. Sequences Analysis
3. Results
3.1. DNA Extraction and PCR Amplification of Target Loci ITS, ARF, and CHS2, and Agarose Gel Electrophoresis
3.2. Clinical and Socio-Demographic Data
3.3. Purification and Sequences Analysis: ITS, CHS2, and ARF Target Loci
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; Valle, A.; Bagagli, E.; Benard, G.; et al. Brazilian guidelines for the clinical management of paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2017, 50, 715–740. [Google Scholar] [CrossRef] [PubMed]
- Restrepo, A.; Tobón, A.M.; Cano, L.E. Paracoccidioidomycosis. In Mandell, Douglas and Bennett’s Principles and Practice of Infectious Diseases, 8th ed.; Bennett, J.E., Dolin, R., Blzser, M.J., Eds.; Elsevier: Philadelphia, PA, USA, 2014; Volume 1, pp. 2995–3002. [Google Scholar]
- Restrepo, A.; Gonzalez, A.; Agudelo, C.A. Paracoccidioidomycosis. In Essentials of Clinical Mycology, 2nd ed.; Springer: New York, NY, USA, 2011; pp. 367–385. [Google Scholar]
- Mendes, R.P.; Cavalcante, R.S.; Marques, S.A.; Marques, M.E.A.; Venturini, J.; Sylvestre, T.F.; Paniago, A.M.M.; Pereira, A.C.; da Silva, J.F.; Fabro, A.T.; et al. Paracoccidioidomycosis: Current Perspectives from Brazil. Open Microbiol. J. 2017, 11, 224–282. [Google Scholar] [CrossRef] [Green Version]
- Mayayo, E.; López-Aracil, V.; Fernández-Torres, B.; Mayayo, R.; Domínguez, M. Report of an imported cutaneous disseminated case of paracoccidioidomycosis. Rev. Iberoam. Micol. 2007, 24, 44–46. [Google Scholar] [CrossRef]
- Martinez, R. New Trends in Paracoccidioidomycosis Epidemiology. J. Fungi. 2017, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Krakhecke-Teixeira, A.G.; Yamauchi, D.H.; Rossi, A.; de Sousa, H.R.; Garces, H.G.; Júnior, J.L.; Júnior, A.O.S.; Felipe, M.S.S.; Bagagli, E.; de Andrade, H.F., Jr.; et al. Clinical and Eco-Epidemiological Aspects of a Novel Hyperendemic Area of Paracoccidioidomycosis in the Tocantins-Araguaia Basin (Northern Brazil), Caused by Paracoccidioides sp. J. Fungi. 2022, 8, 502. [Google Scholar] [CrossRef]
- Teixeira, M.M.; Theodoro, R.C.; de Carvalho, M.J.; Fernandes, L.; Paes, H.C.; Hahn, R.C.; Mendoza, L.; Bagagli, E.; San-Blas, G.; Felipe, M.S. Phylogenetic analysis reveals a high level of speciation in the Paracoccidioides genus. Mol. Phylogenetics Evol. 2009, 52, 273–283. [Google Scholar] [CrossRef]
- Teixeira, M.M.; Theodoro, R.C.; Nino-Vega, G.; Bagagli, E.; Felipe, M.S. Paracoccidioides species complex: Ecology, phylogeny, sexual reproduction, and virulence. PLoS Pathog. 2014, 10, e1004397. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, A.; Hernandez, O. New insights into a complex fungal pathogen: The case of Paracoccidioides spp. Yeast 2016, 33, 113–128. [Google Scholar] [CrossRef] [Green Version]
- Salgado-Salazar, C.; Jones, L.R.; Restrepo, Á.; McEwen, J.G. The human fungal pathogen Paracoccidioides brasiliensis (Onygenales: Ajellomycetaceae) is a complex of two species: Phylogenetic evidence from five mitochondrial markers. Cladistics Int. J. Willi Hennig Soc. 2010, 26, 613–624. [Google Scholar] [CrossRef]
- Matute, D.R.; McEwen, J.G.; Puccia, R.; Montes, B.A.; San-Blas, G.; Bagagli, E.; Rauscher, J.T.; Restrepo, A.; Morais, F.; Niño-Vega, G.; et al. Cryptic speciation and recombination in the fungus Paracoccidioides brasiliensis as revealed by gene genealogies. Mol. Biol. Evol. 2006, 23, 65–73. [Google Scholar] [CrossRef]
- Turissini, D.A.; Gomez, O.M.; Teixeira, M.M.; McEwen, J.G.; Matute, D.R. Species boundaries in the human pathogen Paracoccidioides. Fungal Genet. Biol. FG B 2017, 106, 9–25. [Google Scholar] [CrossRef] [PubMed]
- Bagagli, E.; Sano, A.; Coelho, K.I.; Alquati, S.; Miyaji, M.; de Camargo, Z.P.; Gomes, G.M.; Franco, M.; Montenegro, M.R. Isolation of Paracoccidioides brasiliensis from armadillos (Dasypus noveminctus) captured in an endemic area of paracoccidioidomycosis. Am. J. Trop. Med. Hyg. 1998, 58, 505–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, A.L.; Tobón, A.; Restrepo, A.; Queiroz-Telles, F.; Nucci, M. Epidemiology of endemic systemic fungal infections in Latin America. Med. Mycol. 2011, 49, 785–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desjardins, C.A.; Champion, M.D.; Holder, J.W.; Muszewska, A.; Goldberg, J.; Bailão, A.M.; Brigido, M.M.; Ferreira, M.E.; Garcia, A.M.; Grynberg, M.; et al. Comparative genomic analysis of human fungal pathogens causing paracoccidioidomycosis. PLoS Genet. 2011, 7, e1002345. [Google Scholar] [CrossRef] [Green Version]
- Matute, D.R.; Sepulveda, V.E.; Quesada, L.M.; Goldman, G.H.; Taylor, J.W.; Restrepo, A.; McEwen, J.G. Microsatellite analysis of three phylogenetic species of Paracoccidioides brasiliensis. J. Clin. Microbiol. 2006, 44, 2153–2157. [Google Scholar] [CrossRef] [Green Version]
- Mde, M.T.; Theodoro, R.C.; Lda, S.D.; Nicola, A.M.; Bagagli, E.; Felipe, M.S. Molecular and morphological data support the existence of a sexual cycle in species of the genus Paracoccidioides. Eukaryot. Cell 2013, 12, 380–389. [Google Scholar] [CrossRef] [Green Version]
- Theodoro, R.C.; Teixeira Mde, M.; Felipe, M.S.; Paduan Kdos, S.; Ribolla, P.M.; San-Blas, G.; Bagagli, E. Genus Paracoccidioides: Species recognition and biogeographic aspects. PloS ONE 2012, 7, e37694. [Google Scholar] [CrossRef] [Green Version]
- Hahn, R.C.; Rodrigues, A.M.; Fontes, C.J.; Nery, A.F.; Tadano, T.; Queiroz Lde, P., Jr.; de Camargo, Z.P. Fatal fungemia due to Paracoccidioides lutzii. Am. J. Trop. Med. Hyg. 2014, 91, 394–398. [Google Scholar] [CrossRef] [Green Version]
- Marques-da-Silva, S.H.; Rodrigues, A.M.; de Hoog, G.S.; Silveira-Gomes, F.; Camargo, Z.P. Occurrence of Paracoccidioides lutzii in the Amazon region: Description of two cases. Am. J. Trop. Med. Hyg. 2012, 87, 710–714. [Google Scholar] [CrossRef] [Green Version]
- Mde, M.T.; Theodoro, R.C.; Oliveira, F.F.; Machado, G.C.; Hahn, R.C.; Bagagli, E.; San-Blas, G.; Soares Felipe, M.S. Paracoccidioides lutzii sp. nov.: Biological and clinical implications. Med. Mycol. 2014, 52, 19–28. [Google Scholar] [CrossRef] [Green Version]
- Cocio, T.A.; Nascimento, E.; Kress, M.; Bagagli, E.; Martinez, R. Characterization of a Paracoccidioides spp. strain from southeastern Brazil genotyped as Paracoccidioides restrepiensis (PS3) and review of this phylogenetic species. Genet. Mol. Biol. 2020, 43, e20190201. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; de Waard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, P.D.; Gregory, T.R. The promise of DNA barcoding for taxonomy. Syst. Biol. 2005, 54, 852–859. [Google Scholar] [CrossRef]
- Schindel, D.E.; Miller, S.E. DNA barcoding a useful tool for taxonomists. Nature 2005, 435, 17. [Google Scholar] [CrossRef] [Green Version]
- Meyer, C.P.; Paulay, G.J.P.B. DNA barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 3, e422. [Google Scholar] [CrossRef] [Green Version]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [Green Version]
- Kurtzman, C.P.; Robnett, C.J. Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie Leeuwenhoek 1998, 73, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Ricci, G.; Campanini, E.B.; Nishikaku, A.S.; Puccia, R.; Marques, M.; Bialek, R.; Rodrigues, A.M.; Batista, W.L. PbGP43 Genotyping Using Paraffin-Embedded Biopsies of Human Paracoccidioidomycosis Reveals a Genetically Distinct Lineage in the Paracoccidioides brasiliensis Complex. Mycopathologia 2022, 187, 157–168. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press, Inc.: Cambridge, MA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Zeminian de Oliveira, L.B.; Della Coletta, A.M.; Gardizani, T.P.; Barrozo, L.V.; Miot, H.A.; Faveri, J.D.; Dias-Melicio, L.A. Paracoccidioimycosis and white individuals: Susceptibility and biogeographic aspects in an important endemic area in Brazil. PLoS Negl. Trop. Dis. 2021, 15, e0009086. [Google Scholar] [CrossRef]
- Schwartz, I.S.; Wiederhold, N.P.; Hanson, K.E.; Patterson, T.F.; Sigler, L. Blastomyces helicus, a New Dimorphic Fungus Causing Fatal Pulmonary and Systemic Disease in Humans and Animals in Western Canada and the United States. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2019, 68, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arantes, T.D.; Theodoro, R.C.; Da Graça Macoris, S.A.; Bagagli, E. Detection of Paracoccidioides spp. in environmental aerosol samples. Med. Mycol. 2013, 51, 83–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queiroz-Telles, F.V.; Pietrobom, P.M.P.; Júnior, M.R.; Baptista, R.M.; Peçanha, P.M. New Insights on Pulmonary Paracoccidioidomycosis. Semin. Respir. Crit. Care Med. 2020, 41, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Queiroz-Telles, F.; Escuissato, D.L. Pulmonary paracoccidioidomycosis. Semin. Respir. Crit. Care Med. 2011, 32, 764–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro, B.G.; Hahn, R.C.; Camargo, Z.P.D.; Rodrigues, A.M. Molecular Tools for Detection and Identification of Paracoccidioides Species: Current Status and Future Perspectives. J. Fungi 2020, 6, 293. [Google Scholar] [CrossRef]
- Garcia, N.M.; Del Negro, G.M.; Heins-Vaccari, E.M.; de Melo, N.T.; de Assis, C.M.; Cda, S.L. Paracoccidioides brasiliensis, a new sample isolated from feces of a penguin (Pygoscelis adeliae). Rev. Inst. Med. Trop. Sao Paulo 1993, 35, 227–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cda, S.L.; Vidal, M.S.; Pereira, C.N.; Heins-Vaccari, E.M.; de Melo, N.T.; Sakai-Valente, N.; Arriagada, G.L. Paracoccidioides cerebriformis Moore, 1935. Mycologic and immunochemical study. Rev. Inst. Med. Trop. Sao Paulo 1997, 39, 141–144. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, J.F.; Farrer, R.A.; Desjardins, C.A.; Gallo, J.E.; Sykes, S.; Sakthikumar, S.; Misas, E.; Whiston, E.A.; Bagagli, E.; Soares, C.M.; et al. Genome Diversity, Recombination, and Virulence across the Major Lineages of Paracoccidioides. mSphere 2016, 1, e00213-16. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, M.M.; Cattana, M.E.; Matute, D.R.; Muñoz, J.F.; Arechavala, A.; Isbell, K.; Schipper, R.; Santiso, G.; Tracogna, F.; Sosa, M.; et al. Genomic diversity of the human pathogen Paracoccidioides across the South American continent. Fungal Genet. Biol. FG B 2020, 140, 103395. [Google Scholar] [CrossRef] [PubMed]
- Hahn, R.C.; Hagen, F.; Mendes, R.P.; Burger, E.; Nery, A.F.; Siqueira, N.P.; Guevara, A.; Rodrigues, A.M.; Camargo, Z.P.D. Paracoccidioidomycosis: Current Status and Future Trends. Clin. Microbiol. Rev. 2022, 35, e00233-00221. [Google Scholar] [CrossRef]
- Pereira, A.C.; de Souza, V.B.; Dias-Baptista, I.M.F. Extração de DNA de biópsias de pele fixadas em formalina tamponada e embebidas em parafina e amplificação por PCR. Hansen. Int. Hansen. E Outras Doenças Infecc. 2008, 33, 25–30. [Google Scholar]
- Thavarajah, R.; Mudimbaimannar, V.K.; Elizabeth, J.; Rao, U.K.; Ranganathan, K. Chemical and physical basics of routine formaldehyde fixation. J. Oral Maxillofac. Pathol. JOMFP 2012, 16, 400–405. [Google Scholar] [CrossRef] [Green Version]
- Stanta, G.; Bonin, S.; Perin, R. RNA extraction from formalin-fixed and paraffin-embedded tissues. Methods Mol. Biol. 1998, 86, 23–26. [Google Scholar] [CrossRef]
- Specht, K.; Richter, T.; Müller, U.; Walch, A.; Werner, M.; Höfler, H. Quantitative gene expression analysis in microdissected archival formalin-fixed and paraffin-embedded tumor tissue. Am. J. Pathol. 2001, 158, 419–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Titford, M.E.; Horenstein, M.G. Histomorphologic assessment of formalin substitute fixatives for diagnostic surgical pathology. Arch. Pathol. Lab. Med. 2005, 129, 502–506. [Google Scholar] [CrossRef]
- Feldman, M.Y. Reactions of nucleic acids and nucleoproteins with formaldehyde. Prog. Nucleic Acid Res. Mol. Biol. 1973, 13, 1–49. [Google Scholar] [CrossRef] [PubMed]
- Jackson, V. Studies on histone organization in the nucleosome using formaldehyde as a reversible cross-linking agent. Cell 1978, 15, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Möller, K.; Rinke, J.; Ross, A.; Buddle, G.; Brimacombe, R. The use of formaldehyde in RNA-protein cross-linking studies with ribosomal subunits from Escherichia coli. Eur. J. Biochem. 1977, 76, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Brutlag, D.; Schlehuber, C.; Bonner, J. Properties of formaldehyde-treated nucleohistone. Biochemistry 1969, 8, 3214–3218. [Google Scholar] [CrossRef]
- Ilyin, Y.V.; Georgiev, G.P. Heterogeneity of deoxynucleoprotein particles as evidencec by ultracentrifugation of cesium chloride density gradient. J. Mol. Biol. 1969, 41, 299–303. [Google Scholar] [CrossRef]
- Varshavsky, A.J.; Ilyin, Y.V. Salt treatment of chromatin induces redistribution of histones. Biochim. Biophys. Acta BBA Nucleic Acids Protein Synth. 1974, 340, 207–217. [Google Scholar] [CrossRef]
- Varshavsky, A.J.; Sundin, O.; Bohn, M. A stretch of “late” SV40 viral DNA about 400 bp long which includes the origin of replication is specifically exposed in SV40 minichromosomes. Cell 1979, 16, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.T.; Haselkorn, T.; Bunce, M.; Sanchez, J.J.; Lucas, S.B.; Jewell, L.D.; Van Marck, E.; Worobey, M. The isolation of nucleic acids from fixed, paraffin-embedded tissues-which methods are useful when? PloS ONE 2007, 2, e537. [Google Scholar] [CrossRef]
- Kocjan, B.J.; Hošnjak, L.; Poljak, M. Commercially available kits for manual and automatic extraction of nucleic acids from formalin-fixed, paraffin-embedded (FFPE) tissues. Acta Dermatovenerol. Alp. Pannonica Adriat. 2015, 24, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Moelans, C.B.; ter Hoeve, N.; van Ginkel, J.W.; ten Kate, F.J.; van Diest, P.J. Formaldehyde substitute fixatives. Analysis of macroscopy, morphologic analysis, and immunohistochemical analysis. Am. J. Clin. Pathol. 2011, 136, 548–556. [Google Scholar] [CrossRef]
- Greer, C.E.; Peterson, S.L.; Kiviat, N.B.; Manos, M.M. PCR amplification from paraffin-embedded tissues. Effects of fixative and fixation time. Am. J. Clin. Pathol. 1991, 95, 117–124. [Google Scholar] [CrossRef]
- Sengüven, B.; Baris, E.; Oygur, T.; Berktas, M. Comparison of methods for the extraction of DNA from formalin-fixed, paraffin-embedded archival tissues. Int. J. Med. Sci. 2014, 11, 494–499. [Google Scholar] [CrossRef] [Green Version]
- Kocjan, B.J.; Hošnjak, L.; Poljak, M. Detection of alpha human papillomaviruses in archival formalin-fixed, paraffin-embedded (FFPE) tissue specimens. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2016, 76 (Suppl. 1), S88–S97. [Google Scholar] [CrossRef]
- Lin, J.; Kennedy, S.H.; Svarovsky, T.; Rogers, J.; Kemnitz, J.W.; Xu, A.; Zondervan, K.T. High-quality genomic DNA extraction from formalin-fixed and paraffin-embedded samples deparaffinized using mineral oil. Anal. Biochem. 2009, 395, 265–267. [Google Scholar] [CrossRef] [Green Version]
- Xie, R.; Chung, J.Y.; Ylaya, K.; Williams, R.L.; Guerrero, N.; Nakatsuka, N.; Badie, C.; Hewitt, S.M. Factors influencing the degradation of archival formalin-fixed paraffin-embedded tissue sections. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2011, 59, 356–365. [Google Scholar] [CrossRef] [Green Version]
- Daugaard, I.; Kjeldsen, T.E.; Hager, H.; Hansen, L.L.; Wojdacz, T.K. The influence of DNA degradation in formalin-fixed, paraffin-embedded (FFPE) tissue on locus-specific methylation assessment by MS-HRM. Exp. Mol. Pathol. 2015, 99, 632–640. [Google Scholar] [CrossRef]
- Barrozo, L.V.; Benard, G.; Silva, M.E.S.; Bagagli, E.; Marques, S.A.; Mendes, R.P. First description of a cluster of acute/subacute paracoccidioidomycosis cases and its association with a climatic anomaly. PLoS Negl. Trop. Dis. 2010, 4, e643. [Google Scholar] [CrossRef] [Green Version]
- do Valle, A.C.F.; de Macedo, P.M.; Almeida-Paes, R.; Romão, A.R.; Lazéra, M.D.S.; Wanke, B. Paracoccidioidomycosis after Highway Construction, Rio de Janeiro, Brazil. Emerg. Infect. Dis. 2017, 23, 1917–1919. [Google Scholar] [CrossRef] [Green Version]
- Arantes, T.D.; Theodoro, R.C.; Teixeira Mde, M.; Sde, M.B.; Bagagli, E. Environmental Mapping of Paracoccidioides spp. in Brazil Reveals New Clues into Genetic Diversity, Biogeography and Wild Host Association. PLoS Negl. Trop. Dis. 2016, 10, e0004606. [Google Scholar] [CrossRef] [Green Version]
- Bagagli, E.; Bosco, S.M.; Theodoro, R.C.; Franco, M. Phylogenetic and evolutionary aspects of Paracoccidioides brasiliensis reveal a long coexistence with animal hosts that explain several biological features of the pathogen. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2006, 6, 344–351. [Google Scholar] [CrossRef]
- Bagagli, E.; Franco, M.; Sde, M.B.; Hebeler-Barbosa, F.; Trinca, L.A.; Montenegro, M.R. High frequency of Paracoccidioides brasiliensis infection in armadillos (Dasypus novemcinctus): An ecological study. Med. Mycol. 2003, 41, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Machado, G.C.; Moris, D.V.; Arantes, T.D.; Silva, L.R.; Theodoro, R.C.; Mendes, R.P.; Vicentini, A.P.; Bagagli, E. Cryptic species of Paracoccidioides brasiliensis: Impact on paracoccidioidomycosis immunodiagnosis. Mem. Inst. Oswaldo Cruz 2013, 108, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Bagagli, E.; Theodoro, R.C.; Bosco, S.M.; McEwen, J.G. Paracoccidioides brasiliensis: Phylogenetic and ecological aspects. Mycopathologia 2008, 165, 197–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrycyk, M.F.; Garcia Garces, H.; Bosco, S.M.G.; de Oliveira, S.L.; Marques, S.A.; Bagagli, E. Ecology of Paracoccidioides brasiliensis, P. lutzii and related species: Infection in armadillos, soil occurrence and mycological aspects. Med. Mycol. 2018, 56, 950–962. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) | Amplicon | Reference |
|---|---|---|---|---|
| Nested ITS first round | ITS4 | TCCTCCGCTTATTGATATGC | 634 bp | [31] |
| ITS5 | GGAAGTAAAAGTCGTAACAAGG | |||
| Nested ITS second round | PbITS-E | GAGCTTTGACGTCTGAGACC | 450 bp | [19] |
| Pb-ITST | GTATCCCTACCTGATCCGAG | |||
| ARF | ARF-FWD | TCTCATGGTTGGCCTCGATGCTGCC | 407 bp | [12] |
| ARF-REV | GAGCCTCGACGACACGGTCACGATC | |||
| CHS2 | CHS2 E2–4 Fwd | CTTAACGGTGCCTTCTTTGCGG | 549 bp | [12] |
| CHS2 E2–4 Rev | GTGAAAGTATTGTTGCCCAGCG |
| Characteristics | Acute (n = 11) | Chronic (n = 38) | p-Value | |
|---|---|---|---|---|
| Patient’ age (years) * | 27.2(8–45) | 53.8 (27–81) | - | |
| Gender ** | Male | 54.55% (6/11) | 86.84% (33/38) | 0.0327 |
| Female | 45.45% (5/11) | 13.16% (5/38) | 0.0327 | |
| Ethnicity ** | White | 81.82% (9/11) | 81.58% (31/38) | NS (p < 0.05) |
| Non-white | 18.18% (2/11) | 18.42% (7/38) | NS (p < 0.05) | |
| Smokers ** | Smoker | 18.18% (2/11) | 89.47% (34/38) | <0.0001 |
| Non-smoker | 81.82% (9/11) | 10.53% (4/38) | <0.0001 | |
| Alcoholics ** | Alcoholic | 0% (0/11) | 63.16% (24/38) | 0.0002 |
| Non-alcoholic | 100% (11/11) | 36.84% (14/38) | 0.0002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira, L.B.Z.; Della Coletta, A.M.; Gardizani, T.P.; Garces, H.G.; Bagagli, E.; Trilles, L.; Barrozo, L.V.; Marques, S.d.A.; Faveri, J.D.; Dias-Melicio, L.A. Molecular Phylogenetic Analysis of Paracoccidioides Species Complex Present in Paracoccidioidomycosis Patient Tissue Samples. Microorganisms 2023, 11, 562. https://doi.org/10.3390/microorganisms11030562
de Oliveira LBZ, Della Coletta AM, Gardizani TP, Garces HG, Bagagli E, Trilles L, Barrozo LV, Marques SdA, Faveri JD, Dias-Melicio LA. Molecular Phylogenetic Analysis of Paracoccidioides Species Complex Present in Paracoccidioidomycosis Patient Tissue Samples. Microorganisms. 2023; 11(3):562. https://doi.org/10.3390/microorganisms11030562
Chicago/Turabian Stylede Oliveira, Luciana Bonome Zeminian, Amanda Manoel Della Coletta, Taiane Priscila Gardizani, Hans Garcia Garces, Eduardo Bagagli, Luciana Trilles, Ligia Vizeu Barrozo, Sílvio de Alencar Marques, Julio De Faveri, and Luciane Alarcão Dias-Melicio. 2023. "Molecular Phylogenetic Analysis of Paracoccidioides Species Complex Present in Paracoccidioidomycosis Patient Tissue Samples" Microorganisms 11, no. 3: 562. https://doi.org/10.3390/microorganisms11030562
APA Stylede Oliveira, L. B. Z., Della Coletta, A. M., Gardizani, T. P., Garces, H. G., Bagagli, E., Trilles, L., Barrozo, L. V., Marques, S. d. A., Faveri, J. D., & Dias-Melicio, L. A. (2023). Molecular Phylogenetic Analysis of Paracoccidioides Species Complex Present in Paracoccidioidomycosis Patient Tissue Samples. Microorganisms, 11(3), 562. https://doi.org/10.3390/microorganisms11030562