


Elevated Temperature-Induced Epimicrobiome Shifts in an Invasive Seaweed Gracilaria vermiculophylla

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of G. vermiculophylla Populations
2.2. Experimental Design and Setup
2.3. Sampling of Epibacteria from G. vermiculophylla
2.4. Genomic DNA Extraction and 16S rRNA Gene Amplicon Sequencing
2.5. Sequence Quality Filtering and Taxonomy Annotation
2.6. Statistical Analysis
3. Results
3.1. Sequencing Summary of 16S rRNA Gene Amplicon Sequencing
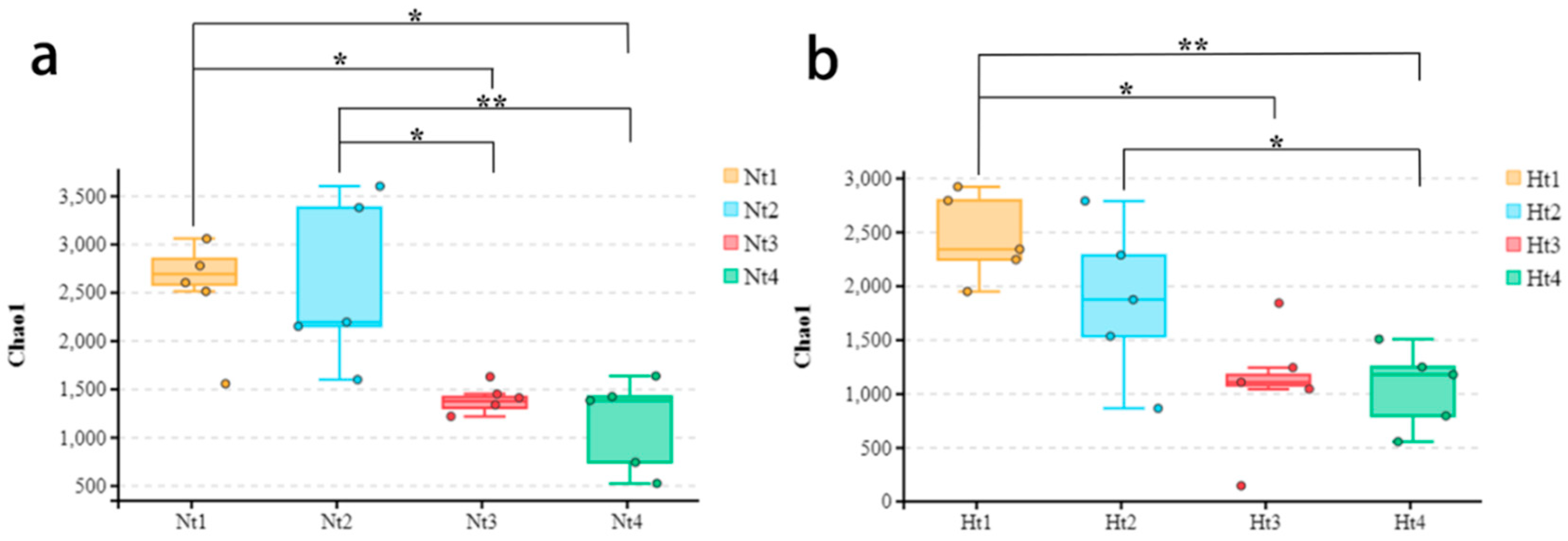
3.2. Effects of Elevated Temperature on Epibacterial Diversity Communities
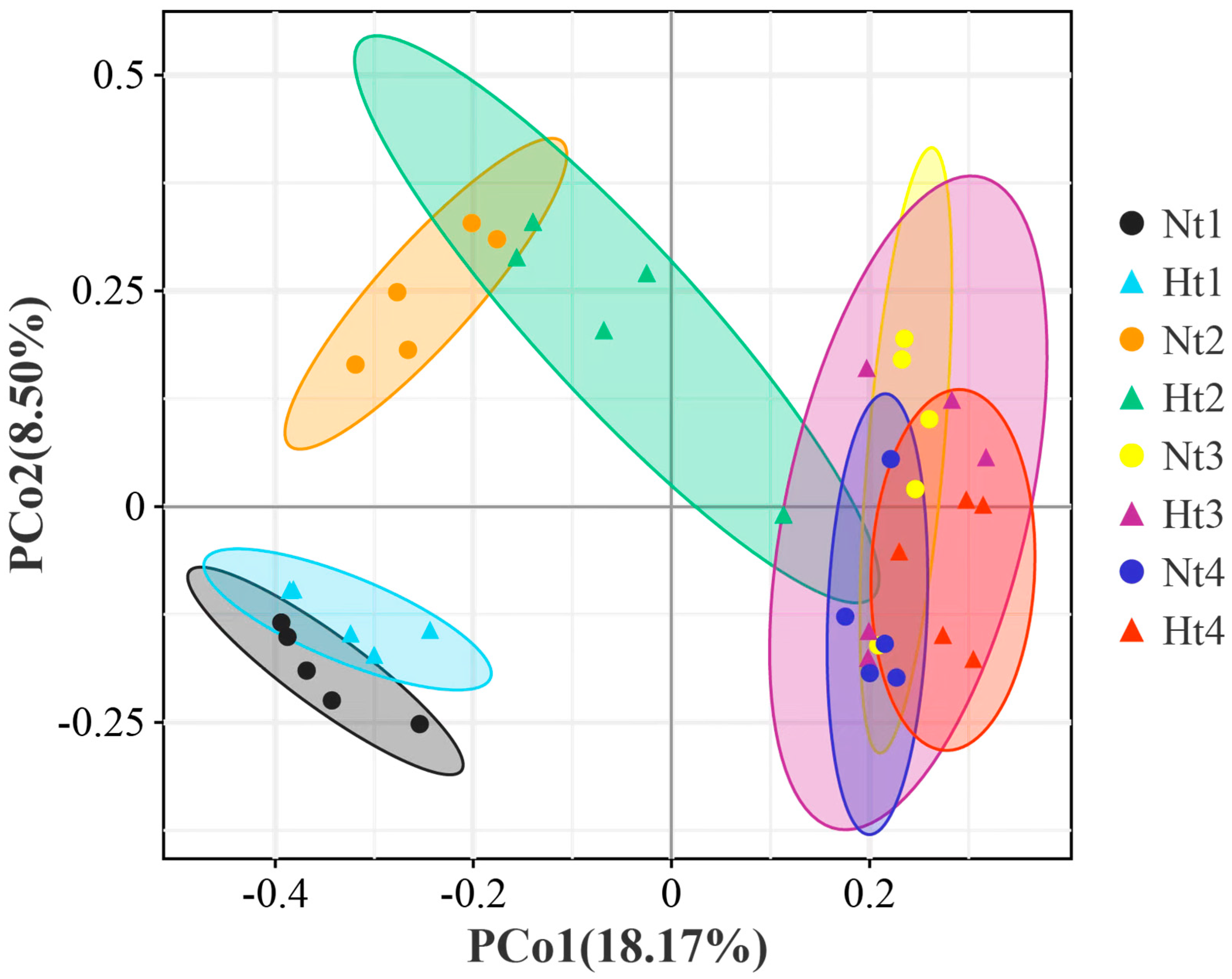
3.3. Species Composition of Epibacterial Communities on G. vermiculophylla
3.4. Core Epibacterial ASVs on G. vermiculophylla at Two Temperature Levels
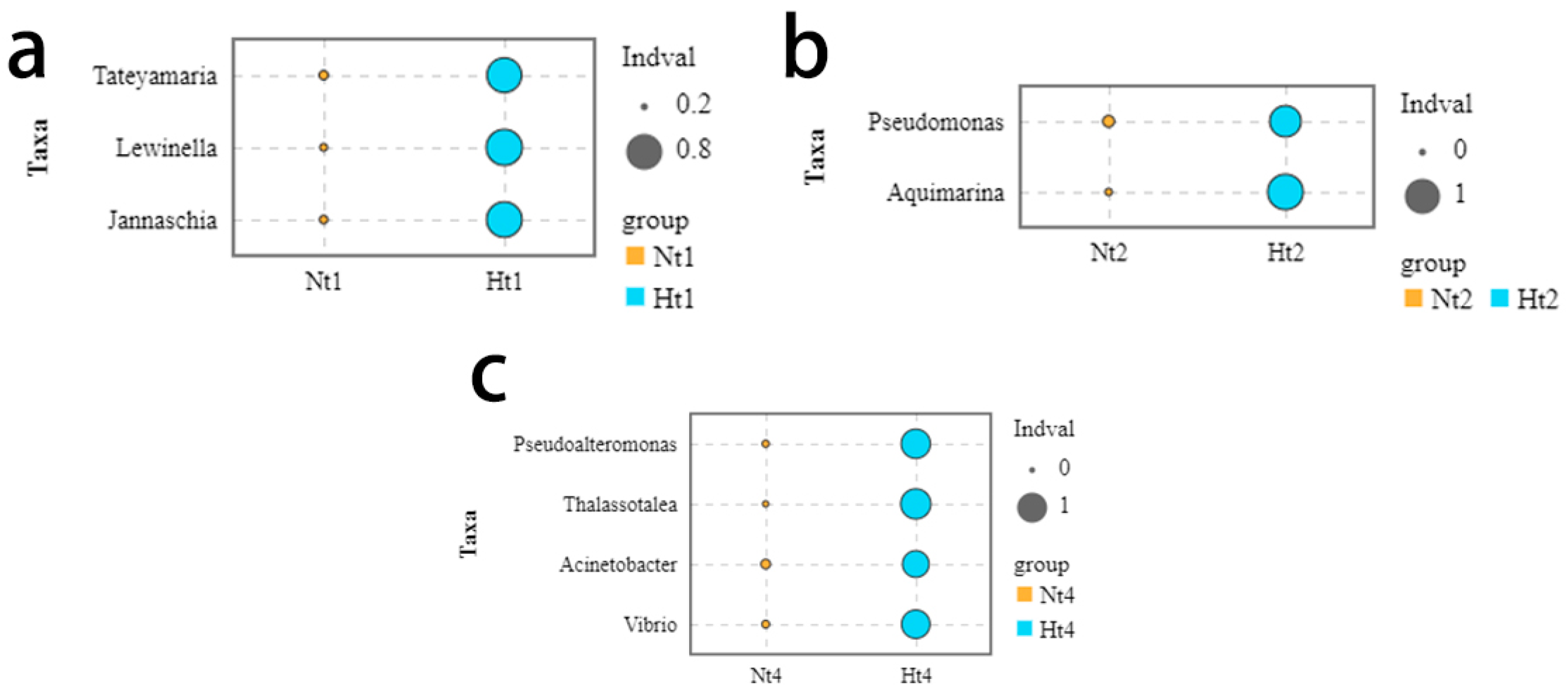
3.5. Indicator Species over Time at Two Temperature Level
3.6. Functional Prediction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goecke, F.; Labes, A.; Wiese, J.; Imhoff, J.F. Chemical interactions between marine macroalgae and bacteria. Mar. Ecol. Prog. Ser. 2010, 409, 267–299. [Google Scholar] [CrossRef]
- Egan, S.; Harder, T.; Burke, C.; Steinberg, P.; Kjelleberg, S.; Thomas, T. The seaweed holobiont: Understanding seaweed–bacteria interactions. FEMS Microbiol. Rev. 2013, 37, 462–476. [Google Scholar] [CrossRef] [Green Version]
- Mensch, B.; Neulinger, S.C.; Künzel, S.; Wahl, M.; Schmitz, R.A. Warming, but Not Acidification, Restructures Epibacterial Communities of the Baltic Macroalga Fucus vesiculosus with Seasonal Variability. Front. Microbiol. 2020, 11, 1471. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.; Portetelle, D.; Michel, G.; Vandenbol, M. Microorganisms living on. macroalgae: Diversity, interactions, and biotechnological applications. Appl. Microbiol. Biotechnol. 2014, 98, 2917–2935. [Google Scholar] [CrossRef] [Green Version]
- Stratil, S.B.; Neulinger, S.C.; Knecht, H.; Friedrichs, A.K.; Wahl, M. Temperature-driven shifts in the epibiotic bacterial community composition of the brown macroalga Fucus vesiculosus. MicrobiologyOpen 2013, 2, 338–349. [Google Scholar] [CrossRef] [Green Version]
- Minich, J.J.; Morris, M.; Brown, M.; Doane, M.; Edwards, M.S.; Michael, T.P.; Dinsdale, E.A. Elevated temperature drives kelp microbiome dysbiosis, while elevated carbon dioxide induces water microbiome disruption. PLoS ONE 2018, 13, e0192772. [Google Scholar] [CrossRef]
- Saha, M.; Ferguson, R.M.W.; Dove, S.; Künzel, S.; Meichssner, R.; Neulinger, S.C.; Petersen, F.O.; Weinberger, F. Salinity and Time Can Alter Epibacterial Communities of an Invasive Seaweed. Front. Microbiol. 2020, 10, 2870. [Google Scholar] [CrossRef] [Green Version]
- Mensch, B.; Neulinger, S.C.; Graiff, A.; Pansch, A.; Künzel, S.; Fischer, M.; Schmitz, R.A. Restructuring of Epibacterial Communities on Fucus vesiculosus forma mytili in Response to Elevated pCO2 and Increased Temperature Levels. Front. Microbiol. 2016, 7, 434. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T. Marine Algae of Japan; Uchida Rokakuho Publishing: Tokyo, Japan, 1998. [Google Scholar]
- Krueger-Hadfield, S.A.; Kollars, N.M.; Strand, A.E.; Byers, J.E.; Shainker, S.J.; Terada, R.; Greig, T.W.; Hammann, M.; Murray, D.C.; Weinberger, F.; et al. Genetic identification of source and likely vector of a widespread marine invader. Ecol. Evol. 2017, 7, 4432–4447. [Google Scholar] [CrossRef] [Green Version]
- Saha, M.; Wiese, J.; Weinberger, F.; Wahl, M. Rapid adaptation to controlling new microbial epibionts in the invaded range promotes invasiveness of an exotic seaweed. J. Ecol. 2016, 104, 969–978. [Google Scholar] [CrossRef] [Green Version]
- Inderjit; Chapman, D.; Ranelletti, M.; Kaushik, S. Invasive marine algae: An ecological perspective. Bot. Rev. 2006, 72, 153–178. [Google Scholar]
- Williams, S.L.; Smith, J.E. A Global Review of the Distribution, Taxonomy, and Impacts of Introduced Seaweeds. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 327–359. [Google Scholar] [CrossRef] [Green Version]
- Saha, M.; Weinberger, F. Microbial “gardening” by a seaweed holobiont: Surface metabolites attract protective and deter pathogenic epibacterial settlement. J. Ecol. 2019, 107, 2255–2265. [Google Scholar] [CrossRef]
- Bonthond, G.; Bayer, T.; Krueger-Hadfield, S.A.; Barboza, F.R.; Nakaoka, M.; Valero, M.; Wang, G.; Künzel, S.; Weinberger, F. How do microbiota associated with an invasive seaweed vary across scales? Mol. Ecol. 2020, 29, 2094–2108. [Google Scholar] [CrossRef]
- Bonthond, G.; Shalygin, S.; Bayer, T.; Weinberger, F. Draft genome and description of Waterburya agarophytonicola gen. nov. sp. nov. (Pleurocapsales, Cyanobacteria): A seaweed symbiont. Antonie Leeuwenhoek 2021, 114, 2189–2203. [Google Scholar] [CrossRef]
- Elken, J.; Lehmann, A.; Myrberg, K. Recent Change—Marine Circulation and Stratification. In Second Assessment of Climate Change for the Baltic Sea Basin. Regional Climate Studies; The BACC II Author Team, Ed.; Springer: Cham, Switzerland, 2015. [Google Scholar] [CrossRef] [Green Version]
- Wahl, M.; Buchholz, B.; Winde, V.; Golomb, D.; Guy-Haim, T.; Müller, J.D.; Rilov, G.; Scotti, M.; Böttcher, M.E. A mesocosm concept for the simulation of near-natural shallow underwater climates: The Kiel Outdoor Benthocosms (KOB). Limnol. Oceanogr. Methods 2015, 13, 651–663. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Gloeckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucl. Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Wickham, H.; Chang, W. ggplot2: An implementation of the Grammar of Graphics. R Package Version 0.7, 2008, 3. Available online: http://CRAN.R-project.org/package=ggplot2 (accessed on 7 December 2022).
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.4-6. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 7 December 2022).
- Chen, H.; Boutros, P.C. VennDiagram: A package for the generation of highly-customizable Venn and Euler diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouillot, D.; Wilson, J. Can We Tell How a Community Was Constructed? A Comparison of Five Evenness Indices for Their Ability to Identify Theoretical Models of Community Construction. Theor. Popul. Biol. 2002, 61, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.W.; Roberts, M.D.W. Package ‘labdsv’: Ordination and Multivariate Analysis for Ecology. R Package Version1.6-1[CP/OL]. 2016. Available online: http://CRAN.R-project.org/package=labdsv (accessed on 7 December 2022).
- McGeoch, M.A.; Van Rensburg, B.J.; Botes, A. The verification and application of bioindicators: A case study of dung beetles in a savanna ecosystem. J. Appl. Ecol. 2002, 39, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Aßhauer, K.P.; Wemheuer, B.; Daniel, R.; Meinicke, P. Tax4Fun: Predicting functional profiles from metagenomic 16S rRNA data. Bioinformatics 2015, 31, 2882–2884. [Google Scholar] [CrossRef] [Green Version]
- Wahl, M.; Barboza, F.R.; Buchholz, B.; Dobretsov, S.; Guy-Haim, T.; Rilov, G.; Schuett, R.; Wolf, F.; Vajedsamiei, J.; Yazdanpanah, M.; et al. Pulsed pressure: Fluctuating impacts of multifactorial environmental change on a temperate macroalgal community. Limnol. Oceanogr. 2021, 66, 4210–4226. [Google Scholar] [CrossRef]
- Wahl, M.; Werner, F.J.; Buchholz, B.; Raddatz, S.; Graiff, A.; Matthiessen, B.; Karsten, U.; Hiebenthal, C.; Hamer, J.; Ito, M.; et al. Season affects strength and direction of the interactive impacts of ocean warming and biotic stress in a coastal seaweed ecosystem. Limnol. Oceanogr. 2020, 65, 807–827. [Google Scholar] [CrossRef] [Green Version]
- Yokoya, N.S.; Kakita, H.; Obika, H.; Kitamura, T. Effects of environmental factors and plant growth regulators on growth of the red alga Gracilaria vermiculophylla from Shikoku Island, Japan. Hydrobiologia 1999, 339–347. [Google Scholar] [CrossRef]
- Weinberger, F.; Buchholz, B.; Karez, R.; Wahl, M. The invasive red alga Gracilaria vermiculophylla in the Baltic Sea: Adaptation to brackish water may compensate for light limitation. Aquat. Biol. 2008, 3, 251–264. [Google Scholar] [CrossRef]
- Hu, Z.-M.; Juan, L.-B. Adaptation mechanisms and ecological consequences of seaweed invasions: A review case of agarophyte Gracilaria vermiculophylla. Biol. Invasions 2014, 16, 967–976. [Google Scholar] [CrossRef]
- Hammann, M.; Wang, G.; Boo, S.M.; Aguilar-Rosas, L.E.; Weinberger, F. Selection of heat-shock resistance traits during the invasion of the seaweed Gracilaria vermiculophylla. Mar. Biol. 2016, 163, 1–11. [Google Scholar] [CrossRef]
- Mancuso, F.P.; Sofie, D.; Anne, W.; Laura, A.; Olivier, D.C. Diversity and temporal dynamics of the epiphytic bacterial communities associated with the canopy-forming seaweed Cystoseira compressa (Esper) Gerloff and Nizamuddin. Front. Microbiol. 2016, 7, 476. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Lee, S.Y.; Kim, W.; Yoon, J.-H. Pseudoalteromonas caenipelagi sp. nov., isolated from a tidal flat. Int. J. Syst. Evol. Microbiol. 2020, 70, 6301–6306. [Google Scholar] [CrossRef]
- Shnit-Orland, M.; Sivan, A.; Kushmaro, A. Antibacterial Activity of Pseudoalteromonas in the Coral Holobiont. Microb. Ecol. 2012, 64, 851–859. [Google Scholar] [CrossRef]
- Liu, H.; Zheng, F.; Sun, X.; Hong, X.; Dong, S.; Wang, B.; Tang, X.; Wang, Y. Identification of the pathogens associated with skin ulceration and peristome tumescence in cultured sea cucumbers Apostichopus japonicus (Selenka). J. Invertebr. Pathol. 2010, 105, 236–242. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, N.; Li, Q.; Ding, J.; Zhan, Y.; Chang, Y. Isolation and characterization of bacteria associated with a syndrome disease of sea urchin Strongylocentrotus intermedius in North China. Aquac. Res. 2012, 44, 691–700. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Y.; Saha, M.; Zhuang, Y.; Chang, L.; Xiao, L.; Wang, G. Pseudoalteromonas piscicida X-8 causes bleaching disease in farmed Saccharina japonica. Aquaculture 2021, 546, 737354. [Google Scholar] [CrossRef]
- Wang, G.; Ren, Y.; Wang, S.; Hou, M.; Weinberger, F. Shifting chemical defence or novel weapons? A review of defence traits in Agarophyton vermiculophyllum and other invasive seaweeds. Mar. Life Sci. Technol. 2022, 4, 138–149. [Google Scholar] [CrossRef]
- Rubio-Portillo, E.; Gago, J.F.; Martínez-García, M.; Vezzulli, L.; Rosselló-Móra, R.; Antón, J.; Ramos-Esplá, A.A. Vibrio communities in scleractinian corals differ according to health status and geographic location in the Mediterranean Sea. Syst. Appl. Microbiol. 2018, 41, 131–138. [Google Scholar] [CrossRef]
- Wright, A.C.; Hill, R.T.; A Johnson, J.; Roghman, M.C.; Colwell, R.R.; Morris, J.G. Distribution of Vibrio vulnificus in the Chesapeake Bay. Appl. Environ. Microbiol. 1996, 62, 717–724. [Google Scholar] [CrossRef] [Green Version]
- Ben-Haim, Y.; Zicherman-Keren, M.; Rosenberg, E. Temperature-Regulated Bleaching and Lysis of the Coral Pocillopora damicornis by the Novel Pathogen Vibrio coralliilyticus. Appl. Environ. Microbiol. 2003, 69, 4236–4242. [Google Scholar] [CrossRef] [Green Version]
- Bally, M.; Garrabou, J. Thermodependent bacterial pathogens and mass mortalities in temperate benthic communities: A new case of emerging disease linked to climate change. Glob. Chang. Biol. 2007, 13, 2078–2088. [Google Scholar] [CrossRef]
- Song, J.K.; Kim, D.; Eun, J.-B.; Choi, B.-D.; Oh, M.J.; Jung, S.J. Identification of cellulolytic bacteria associated with tunic softness syndrome in the sea squirt, Halocynthia roretzi. Food Sci. Biotechnol. 2012, 21, 1405–1411. [Google Scholar] [CrossRef]
- Lian, F.-B.; Jiang, S.; Ren, T.-Y.; Zhou, B.-J.; Du, Z.-J. Thalassotalea algicola sp. nov., an alginate-utilizing bacterium isolated from a red alga. Antonie Van Leeuwenhoek 2021, 114, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.T.; Liu, Y.; Zhong, Z.P.; Liu, H.C.; Liu, Z.P. Thalassotalea marina sp. nov., isolated from a marine recirculating aquaculture system, reclassification of Thalassomonas eurytherma as Thalassotalea eurytherma comb. nov. and emended description of the genus Thalassotalea. Int. J. Syst. Evol. Microbiol. 2015, 65, 4710–4715. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Y.-W.; Li, S.-N.; Zhang, D.-C. Thalassotalea profundi sp. nov. isolated from a deep-sea seamount. Int. J. Syst. Evol. Microbiol. 2017, 67, 3739–3743. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Jung, Y.-T.; Kang, C.-H.; Park, J.-M.; Yoon, J.-H. Thalassotalea ponticola sp. nov., isolated from seawater, reclassification of Thalassomonas fusca as Thalassotalea fusca comb. nov. and emended description of the genus Thalassotalea. Int. J. Syst. Evol. Microbiol. 2014, 64, 3676–3682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.; Zhang, D.; Gui, J.; Wang, J.; Zhu, X.; Lai, Q.; Wang, W.; Xu, H. Thalassotalea mangrovi sp. nov., a bacterium isolated from marine mangrove sediment. Int. J. Syst. Evol. Microbiol. 2019, 69, 3644–3649. [Google Scholar] [CrossRef]
- Kim, K.Y.; Nguyen, T.H.; Kurihara, H.; Kim, S.M. Alpha-glucosidase inhibitory activity of bromophenol purified from the red alga Polyopes lancifolia. J. Food Sci. 2010, 75, H145–H150. [Google Scholar]
- Bengtsson, M.; Sjøtun, K.; Øvreås, L. Seasonal dynamics of bacterial biofilms on the kelp Laminaria hyperborea. Aquat. Microb. Ecol. 2010, 60, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Serebryakova, A.; Aires, T.; Viard, F.; Serrão, E.A.; Engelen, A.H. Summer shifts of bacterial communities associated with the invasive brown seaweed Sargassum muticum are location and tissue dependent. PLoS ONE 2018, 13, e0206734. [Google Scholar] [CrossRef]
- Singh, R.P.; Reddy, C. Seaweed-microbial interactions: Key functions of seaweed-associated bacteria. FEMS Microbiol. Ecol. 2014, 88, 213–230. [Google Scholar] [CrossRef] [Green Version]
- Marzinelli, E.M.; Campbell, A.H.; Valdes, E.Z.; Vergés, A.; Nielsen, S.; Wernberg, T.; de Bettignies, T.; Bennett, S.; Caporaso, J.G.; Thomas, T.; et al. Continental-scale variation in seaweed host-associated bacterial communities is a function of host condition, not geography. Environ. Microbiol. 2015, 17, 4078–4088. [Google Scholar] [CrossRef] [PubMed]
- Romine, M.F.; A Rodionov, D.; Maezato, Y.; Osterman, A.L.; Nelson, W.C. Underlying mechanisms for syntrophic metabolism of essential enzyme cofactors in microbial communities. ISME J. 2017, 11, 1434–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesus, A.; Correia-Da-Silva, M.; Afonso, C.; Pinto, M.; Cidade, H. Isolation and Potential Biological Applications of Haloaryl Secondary Metabolites from Macroalgae. Mar. Drugs 2019, 17, 73. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Wang, G.; Xiao, L.; Xu, X.; Liu, X.; Xu, P.; Lin, X. Bis(2,3-dibromo-4,5-dihydroxybenzyl) Ether, a Marine Algae Derived Bromophenol, Inhibits the Growth of Botrytis cinerea and Interacts with DNA Molecules. Mar. Drugs 2014, 12, 3838–3851. [Google Scholar] [CrossRef]
- Lu, J.; Zhang, X.; Qiu, Q.; Chen, J.; Xiong, J. Identifying Potential Polymicrobial Pathogens: Moving Beyond Differential Abundance to Driver Taxa. Microb. Ecol. 2020, 80, 447–458. [Google Scholar] [CrossRef]
- Xiang, J.; Saha, M.; Zhong, K.; Zhang, Q.; Zhang, D.; Jueterbock, A.; Krueger-Hadfield, S.A.; Wang, G.; Weinberger, F.; Hu, Z. Genome-scale signatures of adaptive gene expression changes in an invasive seaweed Gracilaria vermiculophylla. Mol. Ecol. 2022, 32, 613–627. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diffs | Df | SumsOfSqs | MeanSqs | F Value | R2 | p-Value | Significant |
|---|---|---|---|---|---|---|---|
| Nt1 vs. Ht1 | 1 | 0.3561 | 0.3561 | 2.6577 | 0.2494 | 0.031 | * |
| Nt2 vs. Ht2 | 1 | 0.4314 | 0.4314 | 2.0887 | 0.2070 | 0.011 | * |
| Nt3 vs. Ht3 | 1 | 0.3312 | 0.3312 | 0.8277 | 0.0938 | 0.886 | |
| Nt4 vs. Ht4 | 1 | 0.6326 | 0.6326 | 1.5713 | 0.1642 | 0.007 | ** |
| Nt1 vs. Nt2 vs. Nt3 vs. Nt4 | 3 | 2.8824 | 0.9608 | 3.4462 | 0.3925 | 0.001 | ** |
| Ht1 vs. Ht2 vs. Ht3 vs. Ht4 | 3 | 2.5944 | 0.8648 | 2.9528 | 0.3564 | 0.001 | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Düsedau, L.; Ren, Y.; Hou, M.; Wahl, M.; Hu, Z.-M.; Wang, G.; Weinberger, F. Elevated Temperature-Induced Epimicrobiome Shifts in an Invasive Seaweed Gracilaria vermiculophylla. Microorganisms 2023, 11, 599. https://doi.org/10.3390/microorganisms11030599
Düsedau L, Ren Y, Hou M, Wahl M, Hu Z-M, Wang G, Weinberger F. Elevated Temperature-Induced Epimicrobiome Shifts in an Invasive Seaweed Gracilaria vermiculophylla. Microorganisms. 2023; 11(3):599. https://doi.org/10.3390/microorganisms11030599
Chicago/Turabian StyleDüsedau, Luisa, Yifei Ren, Minglei Hou, Martin Wahl, Zi-Min Hu, Gaoge Wang, and Florian Weinberger. 2023. "Elevated Temperature-Induced Epimicrobiome Shifts in an Invasive Seaweed Gracilaria vermiculophylla" Microorganisms 11, no. 3: 599. https://doi.org/10.3390/microorganisms11030599
APA StyleDüsedau, L., Ren, Y., Hou, M., Wahl, M., Hu, Z.-M., Wang, G., & Weinberger, F. (2023). Elevated Temperature-Induced Epimicrobiome Shifts in an Invasive Seaweed Gracilaria vermiculophylla. Microorganisms, 11(3), 599. https://doi.org/10.3390/microorganisms11030599