

Current Trends in Bioaugmentation Tools for Bioremediation: A Critical Review of Advances and Knowledge Gaps


Abstract
:1. Introduction
2. Sources of Microorganisms for Bioaugmentation
3. Obtaining Microorganism Degraders: Sources and Methods
4. Preparation of the Inoculum
5. Application of Immobilised Cells

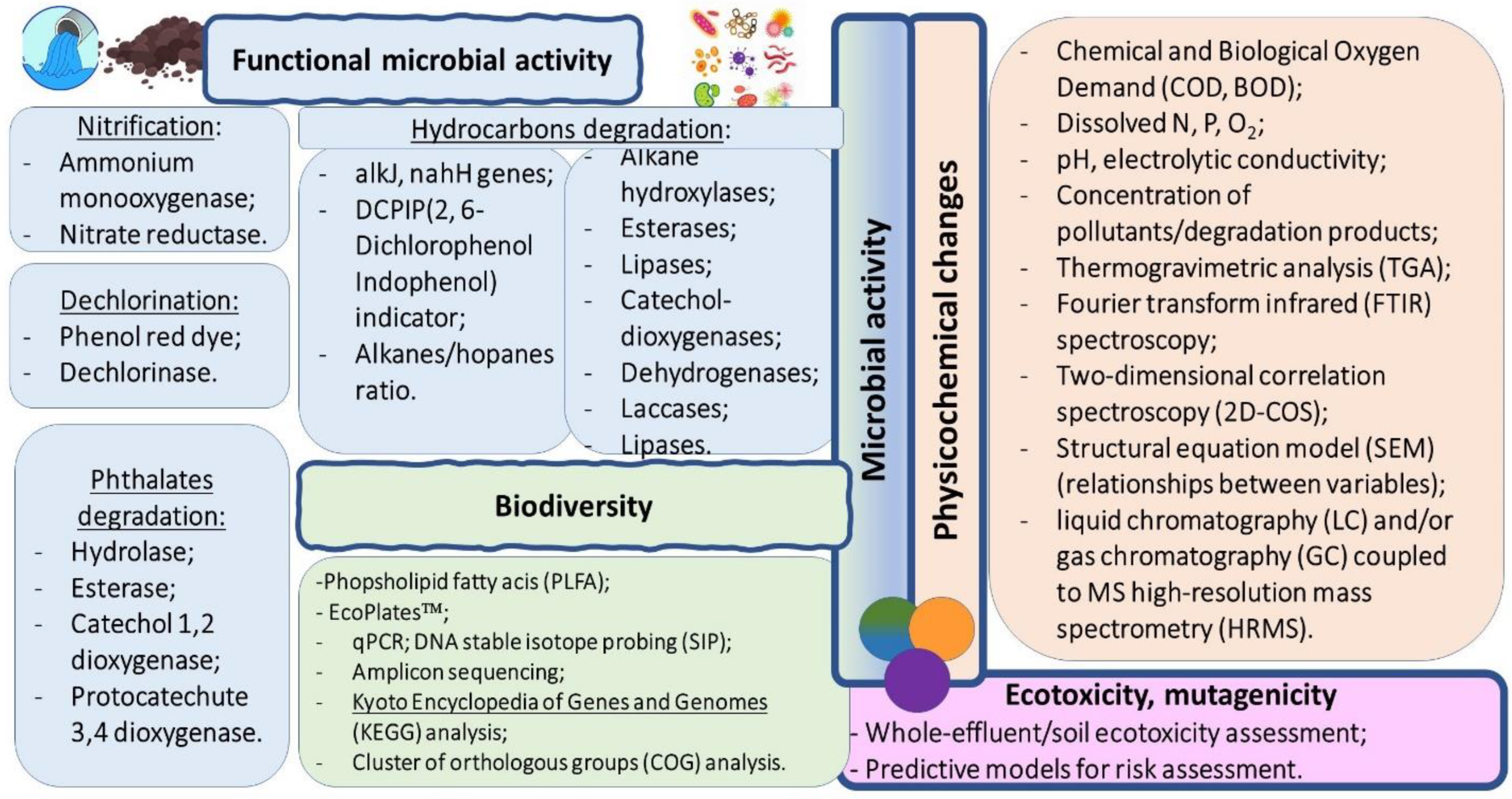{kind=link}
| Carrier, Technique | Microorganism | Effect | Reference |
|---|---|---|---|
| Shell encapsulation | Bradyrhizobium | Protector: albumin-trehalose. Stabilized desiccated cells for 4 months at high relative humidity, increased the glass transition temperature | [62] |
| Bark biochar | Activated sludge | Vertical flow mesocosm-constructed wetland. Removal (>40%) of irbesartan and carbamazepine | [63] |
| Biochar | 10% adapted thermophilic Methanosarcina thermophila on 2 g/L biochar | Anaerobic digestion. Methane yield increased up to 35% | [64] |
| Perlite | 1010 CFU/g | Anaerobic digestion. Biodegradation of olive cake | [65] |
| Fe-modified zeolite grains | 1010 cells/g of biocomposite (selected microbial consortium) | Soil microcosm. 80% herbicide MCPA (2-methyl-4-chlorophenoxyacetic acid) | [66] |
| High-density polyethylene (HDPE) carrier | Sphingomonas rubra BH3T as a quorum-sensing bacteria | Moving bed biofilm reactor (MBBR) at 5 °C. Higher chemical oxygen demand and NH4 +-N removal rate (93% and 75%). Co-culture with Nitrospira. The increased biofilm thickness (60.23%) during the whole operating time, accompanied by more potent adhesion force (61.59%), was related to increased polysaccharides and proteins in the biofilm | [61] |
| Gel-immobilization enhanced with biochar | Ammonia-tolerant methanogens | Continuous biogas reactor. Long-term ammonia resistance | [67] |
| Aerobic granules with microalgae | Aerobic granular sludge from aquaculture WW | Continuous flow granular reactors. Removed up to 77% and 80% of ammonium and nitrite | [68]. |
| Algal organic matter (AOM) and humic substances (HS) | Injection of contaminant-degrading microorganisms | Decrease in removal of cyanotoxins like microcystin-LR (MC-LR). Decreases in MC-LR biodegradation rate of 1.6- and 3.4-fold in the presence of AOM and HS, respectively | [69] |
6. Bioaugmentation Strategy
7. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ahmad, I.; Abdullah, N.; Koji, I.; Yuzir, A.; Mohamad, S.E.; Show, P.L.; Cheah, W.Y.; Khoo, K.S. The role of restaurant wastewater for producing bioenergy towards a circular bioeconomy: A review on composition, environmental impacts, and sustainable integrated management. Environ. Res. 2022, 214, 113854. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, S.; Geng, N.; Wu, Z.; Xiong, W.; Su, H. Artificially constructing mixed bacteria system for bioaugmentation of nitrogen removal from saline wastewater at low temperature. J. Environ. Manag. 2022, 324, 116351. [Google Scholar] [CrossRef] [PubMed]
- Bokade, P.; Gaur, V.K.; Tripathi, V.; Bobate, S.; Manickam, N.; Bajaj, A. Bacterial remediation of pesticide polluted soils: Exploring the feasibility of site restoration. J. Hazard. Mater. 2023, 441, 129906. [Google Scholar] [CrossRef]
- Hudari, M.S.B.; Richnow, H.; Vogt, C.; Nijenhuis, I. Effect of temperature on microbial reductive dehalogenation of chlorinated ethenes: A review. FEMS Microbiol. Ecol. 2022, 98, fiac081. [Google Scholar] [CrossRef]
- Dutta, N.; Usman, M.; Ashraf, M.A.; Luo, G.; Zhang, S. A critical review of recent advances in the bio-remediation of chlorinated substances by microbial dechlorinators. Chem. Eng. J. Adv. 2022, 12, 100359. [Google Scholar] [CrossRef]
- Demir, Ö.; Atasoy, A.D.; Çalış, B.; Çakmak, Y.; Di Capua, F.; Sahinkaya, E.; Uçar, D. Impact of temperature and biomass augmentation on biosulfur-driven autotrophic denitrification in membrane bioreactors treating real nitrate-contaminated groundwater. Sci. Total Environ. 2022, 853, 158470. [Google Scholar] [CrossRef]
- Dagliya, M.; Satyam, N.; Sharma, M.; Garg, A. Experimental study on mitigating wind erosion of calcareous desert sand using spray method for microbially induced calcium carbonate precipitation. J. Rock Mech. Geotech. Eng. 2022, 14, 1556–1567. [Google Scholar] [CrossRef]
- Kalneniece, K.; Gudra, D.; Lielauss, L.; Selga, T.; Fridmanis, D.; Terauds, J.; Muter, O. Batch-mode stimulation of hydrocarbons biodegradation in freshwater sediments from historically contaminated Alūksne lake. J. Contam. Hydrol. 2022, 253, 104103. [Google Scholar] [CrossRef]
- Gao, D.; Zhao, H.; Wang, L.; Li, Y.; Tang, T.; Bai, Y.; Liang, H. Current and emerging trends in bioaugmentation of organic contaminated soils: A review. J. Environ. Manag. 2022, 320, 115799. [Google Scholar] [CrossRef]
- Sharma, P.; Parakh, S.K.; Singh, S.P.; Parra-Saldivar, R.; Kim, S.-H.; Varjani, S.; Tong, Y.W. A critical review on microbes-based treatment strategies for mitigation of toxic pollutants. Sci. Total Environ. 2022, 834, 155444. [Google Scholar] [CrossRef]
- Ntroumpogianni, G.C.; Giannoutsou, E.; Karagouni, A.D.; Savvides, A.L. Bacterial Isolates from Greek Sites and Their Efficacy in Degrading Petroleum. Sustainability 2022, 14, 9562. [Google Scholar] [CrossRef]
- Kalniņš, M.; Andersone-Ozola, U.; Gudra, D.; Sierina, A.; Fridmanis, D.; Ievinsh, G.; Muter, O. Effect of bioaugmentation on the growth and rhizosphere microbiome assembly of hydroponic cultures of Mentha aquatica. Ecol. Genet. Genom. 2022, 22, 100107. [Google Scholar] [CrossRef]
- Zorza, L.; Kalnins, M.; Gudra, D.; Megnis, K.; Fridmanis, D.; Rapoport, A.; Muter, O. Changes in Bacterial Community Structure in Wastewaters in the presence of Saccharomyces Cerevisiae and Benzalkonium Chloride. IOP Conf. Ser. Earth Environ. Sci. 2022, 987, 012010. [Google Scholar] [CrossRef]
- Rimkus, A.; Gudrā, D.; Dubova, L.; Fridmanis, D.; Alsiņa, I.; Muter, O. Stimulation of sewage sludge treatment by carbon sources and bioaugmentation with a sludge-derived microbial consortium. Sci. Total Environ. 2021, 783, 146989. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Jia, Y.; Li, J.; Li, P.; Wang, Y.; Yan, F.; Wu, M.; Fang, W.; Xu, F.; Qiu, Z. Influence of microbial augmentation on contaminated manure composting: Metal immobilization, matter transformation, and bacterial response. J. Hazard. Mater. 2023, 441, 129762. [Google Scholar] [CrossRef]
- Pi, Y.R.; Bao, M.T. Investigation of kinetics in bioaugmentation of crude oil via high-throughput sequencing: Enzymatic activities, bacterial community composition and functions. Pet. Sci. 2022, 19, 1905–1914. [Google Scholar] [CrossRef]
- Meyer, D.D.; Beker, S.A.; Bucker, F.; Peralba, M.C.R.; Frazzon, A.P.G.; Osti, J.F.; Andreazza, R.; Camargo, F.A.O.; Bento, F.M. Bioremediation strategies for diesel and biodiesel in oxisol from southern Brazil. Int. Biodeterior. Biodegrad. 2014, 95, 356–363. [Google Scholar] [CrossRef]
- Ye, T.; Liu, H.; Qi, W.; Qu, J. Removal of pharmaceutical in a biogenic/chemical manganese oxide system driven by manganese-oxidizing bacteria with humic acids as sole carbon source. J. Environ. Sci. 2023, 126, 734–741. [Google Scholar] [CrossRef]
- Guo, Y.; Gao, J.; Zhao, Y.; Liu, Y.; Zhao, M.; Li, Z. Mitigating the inhibition of antibacterial agent chloroxylenol on nitrification system-The role of Rhodococcus ruber in a bioaugmentation system. J. Hazard. Mater. 2023, 447, 130758. [Google Scholar] [CrossRef]
- Zhu, G.; Zhang, H.; Yuan, R.; Huang, M.; Liu, F.; Li, M.; Zhang, Y.; Rittmann, B.E. How Comamonas testosteroni and Rhodococcus ruber enhance nitrification in the presence of quinoline. Water Res. 2023, 229, 119455. [Google Scholar] [CrossRef]
- Bai, F.; Tian, H.; Wang, C.; Ma, J. Treatment of nanofiltration concentrate of landfill leachate using advanced oxidation processes incorporated with bioaugmentation. Environ. Pollut. 2023, 318, 120827. [Google Scholar] [CrossRef]
- Larkin, M.J.; Kulakov, L.A.; Allen, C.C.R. Biodegradation and Rhodococcus-Masters of catabolic versatility. Curr. Opin. Biotechnol. 2005, 16, 282–290. [Google Scholar] [CrossRef]
- Liang, Y.; Van Nostrand, J.D.; Deng, Y.; He, Z.; Wu, L.; Zhang, X.; Li, G.; Zhou, J. Functional gene diversity of soil microbial communities from five oil-contaminated fields in China. ISME J. 2011, 5, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Lara-Moreno, A.; Morillo, E.; Merchán, F.; Madrid, F.; Villaverde, J. Bioremediation of a trifluralin contaminated soil using bioaugmentation with novel isolated bacterial strains and cyclodextrin. Sci. Total Environ. 2022, 840, 156695. [Google Scholar] [CrossRef]
- Lara-Moreno, A.; Morillo, E.; Merchán, F.; Madrid, F.; Villaverde, J. Chlorpyrifos Removal in an Artificially Contaminated Soil Using Novel Bacterial Strains and Cyclodextrin. Evaluation of Its Effectiveness by Ecotoxicity Studies. Agronomy 2022, 12, 1971. [Google Scholar] [CrossRef]
- Zhou, N.; Guo, H.; Liu, Q.; Zhang, Z.; Sun, J.; Wang, H. Bioaugmentation of polycyclic aromatic hydrocarbon (PAH)-contaminated soil with the nitrate-reducing bacterium PheN7 under anaerobic condition. J. Hazard. Mater. 2022, 439, 129643. [Google Scholar] [CrossRef] [PubMed]
- Fontalvo, N.P.M.; Gamero, W.B.M.; Ardila, H.A.M.; Gonzalez, A.F.P.; Ramos, C.G.; Muñoz, A.E.P. Removal of Nitrogenous Compounds from Municipal Wastewater Using a Bacterial Consortium: An Opportunity for More Sustainable Water Treatments. Water. Air. Soil Pollut. 2022, 233, 339. [Google Scholar] [CrossRef]
- You, Y.; Chi, Y.; Chen, X.; Wang, J.; Wang, R.; Li, r.; Chu, S.; Yang, X.; Zhang, D.; Zhou, P. A sustainable approach for bioremediation of secondary salinized soils: Studying remediation efficiency and soil nitrate transformation by bioaugmentation. Chemosphere 2022, 300, 134580. [Google Scholar] [CrossRef]
- Zenteno-Rojas, A.; Rincón-Molina, C.I.; Martinez-Romero, E.; Manzano-Gomez, L.A.; Rincón-Molina, F.A.; Ruiz-Valdiviezo, V.M.; Rodriguez, R.I.C.; Maldonato, J.J.V.; Rincón-Rosales, R. Biostimulation and Bioaugmentation of Soils Contaminated with Decachlorobiphenyl (PCB-209) Using Native Bacterial Strains Individually and in Consortia. Sustainability 2022, 14, 9068. [Google Scholar] [CrossRef]
- Pekkoh, J.; Chaichana, C.; Thurakit, T.; Phinyo, K.; Lomakool, S.; Ruangrit, K.; Duangjan, K.; Suwannarach, N.; Kumla, J.; Cheirsilp, B.; et al. Dual-bioaugmentation strategy to enhance the formation of algal-bacteria symbiosis biofloc in aquaculture wastewater supplemented with agricultural wastes as an alternative nutrient sources and biomass support materials. Bioresour. Technol. 2022, 359, 127469. [Google Scholar] [CrossRef]
- Chen, X.; Lin, H.; Dong, Y.; Li, B.; Liu, C.; Yin, T. Mechanisms underlying enhanced bioremediation of sulfamethoxazole and zinc(II) by Bacillus sp. SDB4 immobilized on biochar. J. Clean. Prod. 2022, 370, 133483. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, M.; Li, C.; Pan, X.; Lv, N.; Ye, Z.; Zhu, G.; Zhao, Q.; Cai, G. Inoculating indoleacetic acid bacteria promotes the enrichment of halotolerant bacteria during secondary fermentation of composting. J. Environ. Manag. 2022, 322, 116021. [Google Scholar] [CrossRef]
- Liu, Q.; Gao, S.; Zhou, Q.; Xu, R.; Li, Z.; Hou, Y.; Sun, Y.; Huang, C. Bio-augmentation of the filler-enhanced denitrifying sulfide removal process in expanded granular sludge bed reactors. Environ. Res. 2022, 212, 113253. [Google Scholar] [CrossRef]
- Wang, Q.; Kong, J.; Liang, J.; El-Din, M.G.; Zhao, P.; Xie, W.; Chen, C. Nitrogen removal intensification of aerobic granular sludge through bioaugmentation with ‘heterotrophic nitrification-aerobic denitrification’ consortium during petroleum wastewater treatment. Bioresour. Technol. 2022, 361, 127719. [Google Scholar] [CrossRef]
- Borella, L.; Novello, G.; Gasparotto, M.; Renella, G.; Roverso, M.; Bogialli, S.; Filippini, F.; Sforza, E. Design and experimental validation of an optimized microalgae-bacteria consortium for the bioremediation of glyphosate in continuous photobioreactors. J. Hazard. Mater. 2023, 441, 129921. [Google Scholar] [CrossRef]
- Szentgyörgyi, F.; Benedek, T.; Fekete, D.; Táncsics, A.; Harkai, P.; Kriszt, B. Development of a bacterial consortium from Variovorax paradoxus and Pseudomonas veronii isolates applicable in the removal of BTEX. AMB Express 2022, 12, 4. [Google Scholar] [CrossRef]
- Ali, N.; Khanafer, M.; Al-Awadhi, H. Indigenous oil-degrading bacteria more efficient in soil bioremediation than microbial consortium and active even in super oil-saturated soils. Front. Microbiol. 2022, 13, 950051. [Google Scholar] [CrossRef]
- Wang, Z.; Su, J.; Ali, A.; Sun, Y.; Li, Y.; Yang, W.; Zhang, R. Enhanced removal of fluoride, nitrate, and calcium using self-assembled fungus-flexible fiber composite microspheres combined with microbially induced calcium precipitation. Chemosphere 2022, 302, 134848. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Guo, Z.; Zhang, L.; Hu, H.; Li, C.; Lin, Z.; Zhen, Z.; Zhou, J.L. A novel aerobic denitrifying phosphate-accumulating bacterium efficiently removes phthalic acid ester, total nitrogen and phosphate from municipal wastewater. J. Water Process Eng. 2023, 52, 103532. [Google Scholar] [CrossRef]
- Li, A.; Fan, J.; Jia, Y.; Tang, X.; Chen, J.; Shen, C. Phenotype and metabolism alterations in PCB-degrading Rhodococcus biphenylivorans TG9T under acid stress. J. Environ. Sci. 2023, 127, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Meng, N.; Su, J.; Li, Y.; Gu, J.; Wang, Y.; Wang, J.; Qu, Y.; Zhao, Z.; Sun, Y. Unraveling the skatole biodegradation process in an enrichment consortium using integrated omics and culture-dependent strategies. J. Environ. Sci. 2023, 127, 688–699. [Google Scholar] [CrossRef]
- Aguilar-Romero, I.; van Dillewijn, P.; Nesme, J.; Sorensen, S.J.; Nogales, R.; Delgado-Moreno, L.; Romero, E. A novel and affordable bioaugmentation strategy with microbial extracts to accelerate the biodegradation of emerging contaminants in different media. Sci. Total Environ. 2022, 834, 155234. [Google Scholar] [CrossRef] [PubMed]
- Curiel-Alegre, S.; Velasco-Arroyo, B.; Rumbo, C.; Khan, A.H.A.; Tamayo-Ramos, J.A.; Rad, C.; Gallego, J.L.R.; Barros, R. Evaluation of biostimulation, bioaugmentation, and organic amendments application on the bioremediation of recalcitrant hydrocarbons of soil. Chemosphere 2022, 307, 135638. [Google Scholar] [CrossRef]
- Pimenov, N.V.; Nikolaev, Y.A.; Dorofeev, A.G.; Grachev, V.A.; Kallistova, A.Y.; Kanapatskii, T.A.; Litti, Y.V.; Gruzdev, E.V.; Begmatov, S.A.; Ravin, N.V.; et al. Introduction of Exogenous Activated Sludge as a Way to Enhance the Efficiency of Nitrogen Removal in the Anammox Process. Microbiol. Russ. Fed. 2022, 91, 356–363. [Google Scholar] [CrossRef]
- Saeed, M.; Ilyas, N.; Jayachandran, K.; Shabir, S.; Akhtar, N.; Shahzad, A.; Sayyed, R.Z.; Bano, A. Advances in Biochar and PGPR engineering system for hydrocarbon degradation: A promising strategy for environmental remediation. Environ. Pollut. 2022, 305, 119282. [Google Scholar] [CrossRef]
- Liu, A.; Wang, W.; Chen, X.; Zheng, X.; Fu, W.; Wang, G.; Ji, J.; Guan, C. Phytoremediation of DEHP and heavy metals co-contaminated soil by rice assisted with a PGPR consortium: Insights into the regulation of ion homeostasis, improvement of photosynthesis and enrichment of beneficial bacteria in rhizosphere soil. Environ. Pollut. 2022, 314, 120303. [Google Scholar] [CrossRef]
- Li, Q.; Xing, Y.; Huang, B.; Chen, X.; Ji, X.; Fu, X.; Li, T.; Wang, J.; Chen, G.; Zhang, Q. Rhizospheric mechanisms of Bacillus subtilis bioaugmentation-assisted phytostabilization of cadmium-contaminated soil. Sci. Total Environ. 2022, 825, 154136. [Google Scholar] [CrossRef]
- Tyagi, M.; da Fonseca, M.M.R.; de Carvalho, C.C.C.R. Bioaugmentation and biostimulation strategies to improve the effectiveness of bioremediation processes. Biodegradation 2011, 22, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Alao, M.B.; Adebayo, E.A. Current advances in microbial bioremediation of surface and ground water contaminated by hydrocarbon. Dev. Wastewater Treat. Res. Processes 2022, 89–116. [Google Scholar] [CrossRef]
- Okeke, E.S.; Okoye, C.O.; Ezeorba, T.P.C.; Mao, G.; Chen, Y.; Xu, H.; Song, C.; Feng, W.; Wu, X. Emerging bio-dispersant and bioremediation technologies as environmentally friendly management responses toward marine oil spill: A comprehensive review. J. Environ. Manag. 2022, 322, 116123. [Google Scholar] [CrossRef]
- Kayastha, V.; Patel, J.; Kathrani, N.; Varjani, S.; Bilal, M.; Show, P.L.; Kim, S.-H.; Bontempi, E.; Bhatia, S.K.; Bui, X.-T. New Insights in factors affecting ground water quality with focus on health risk assessment and remediation techniques. Environ. Res. 2022, 212, 113171. [Google Scholar] [CrossRef] [PubMed]
- Ke, Z.; Wang, S.; Dai, W.; Jia, W.; Mu, Y.; Jiang, J.; Chen, K. Engineering of the chloroaniline-catabolic plasmid pDCA-1 and its potential for genetic bioaugmentation. Int. Biodeterior. Biodegrad. 2022, 172, 105435. [Google Scholar] [CrossRef]
- Varner, P.M.; Allemann, M.N.; Michener, J.K.; Gunsch, C.K. The effect of bacterial growth strategies on plasmid transfer and naphthalene degradation for bioremediation. Environ. Technol. Innov. 2022, 28, 102910. [Google Scholar] [CrossRef]
- Zhou, X.; Li, J.; Zhang, J.; Deng, F.; Chen, Y.; Zhou, P.; Li, D. Bioaugmentation mechanism on humic acid formation during composting of food waste. Sci. Total Environ. 2022, 830, 154783. [Google Scholar] [CrossRef]
- Foster, K.R.; Bell, T. Competition, not cooperation, dominates interactions among culturable microbial species. Curr. Biol. 2012, 22, 1845–1850. [Google Scholar] [CrossRef] [Green Version]
- Hosoda, K.; Tsuda, S.; Kadowaki, K.; Nakamura, Y.; Nakano, T.; Ishii, K. Population-reaction model and microbial experimental ecosystems for understanding hierarchical dynamics of ecosystems. BioSystems 2016, 140, 28–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muter, O.; Makarenkova, G.; Vecstaudza, D.; Shvirksts, K.; Grube, M.; Kalnenieks, U. Optimization of bacterial biomass production for bioaugmentation of the hydrocarbons contaminated sites. Res. J. Biotechnol. 2019, 14, 39–51. [Google Scholar]
- Pornkulwat, P.; Khan, E.; Powtongsook, S.; Mhuantong, W.; Chawengkijwanich, C.; Limpiyakorn, T. Influence of ammonia and NaCl on nitrifying community and activity: Implications for formulating nitrifying culture augmentation. Sci. Total Environ. 2022, 833, 155132. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.K.; Mustapha, H.I.; Singh, B.; Sharma, Y.C. Bioremediation of petroleum contaminated soil-water resources using neat biodiesel: A review. Sustain. Energy Technol. Assess. 2022, 53, 102703. [Google Scholar] [CrossRef]
- Dike, C.C.; Hakeem, I.G.; Rani, A.; Surapaneni, A.; Khudur, L.; Shah, K.; Ball, A.S. The co-application of biochar with bioremediation for the removal of petroleum hydrocarbons from contaminated soil. Sci. Total Environ. 2022, 849, 157753. [Google Scholar] [CrossRef]
- Fu, H.; Wang, J.; Ren, H.; Ding, L. Acceleration of start-up of moving bed biofilm reactor at low temperature by adding specialized quorum sensing bacteria. Bioresour. Technol. 2022, 358, 127249. [Google Scholar] [CrossRef] [PubMed]
- Aukema, K.G.; Wang, M.; de Souza, B.; O’Keane, S.; Clipsham, M.; Wackett, L.P.; Aksan, A. Core-shell encapsulation formulations to stabilize desiccated Bradyrhizobium against high environmental temperature and humidity. Microb. Biotechnol. 2022, 15, 2391–2400. [Google Scholar] [CrossRef]
- Lei, Y.; Rijnaarts, H.; Langenhoff, A. Mesocosm constructed wetlands to remove micropollutants from wastewater treatment plant effluent: Effect of matrices and pre-treatments. Chemosphere 2022, 305, 135306. [Google Scholar] [CrossRef]
- Lee, J.T.E.; Lim, E.Y.; Zhang, L.; Tsui, T.-H.; Tian, H.; Yan, M.; Lim, S.; Majid, M.A.; Jong, M.-C.; Zhang, J.; et al. Methanosarcina thermophila bioaugmentation and its synergy with biochar growth support particles versus polypropylene microplastics in thermophilic food waste anaerobic digestion. Bioresour. Technol. 2022, 360, 127531. [Google Scholar] [CrossRef] [PubMed]
- Ivankovic, T.; Kontek, M.; Mihalic, V.; Ressler, A.; Jurisic, V. Perlite as a Biocarrier for Augmentation of Biogas-Producing Reactors from Olive (Olea europaea) Waste. Appl. Sci. 2022, 12, 8808. [Google Scholar] [CrossRef]
- Gorodylova, N.; Seron, A.; Michel, K.; Joulian, C.; Delorme, F.; Soulier, C.; Bresch, S.; Garreau, C.; Giovanelli, F.; Michel, C. Zeolite-supported biofilms as inoculants for the treatment of MCPA-polluted soil and sand by bioaugmentation: A microcosm study. Appl. Soil Ecol. 2022, 180, 104614. [Google Scholar] [CrossRef]
- Yan, Y.; Yan, M.; Ravenni, G.; Angelidaki, I.; Fu, D.; Fotidis, I.A. Biochar enhanced bioaugmentation provides long-term tolerance under increasing ammonia toxicity in continuous biogas reactors. Renew. Energy 2022, 195, 590–597. [Google Scholar] [CrossRef]
- Santorio, S.; Rio, A.V.D.; Amorim, C.L.; Couto, A.T.; Arregui, L.; Castro, P.M.L.; Mosquera-Corral, A. Microalgae-bacterial biomass outperforms PN-anammox biomass for oxygen saving in continuous-flow granular reactors facing extremely low-strength freshwater aquaculture streams. Chemosphere 2022, 308, 136184. [Google Scholar] [CrossRef]
- Baranwal, P.; Kang, D.W.; Seo, Y. Impacts of algal organic matter and humic substances on microcystin-LR removal and their biotransformation during the biodegradation process. Sci. Total Environ. 2022, 852, 157993. [Google Scholar] [CrossRef]
- Linsong, H.; Lianhua, L.; Ying, L.; Changrui, W.; Yongming, S. Bioaugmentation with methanogenic culture to improve methane production from chicken manure in batch anaerobic digestion. Chemosphere 2022, 303, 135127. [Google Scholar] [CrossRef]
- Lee, J.T.E.; Dutta, N.; Zhang, L.; Tsui, T.T.H.; Lim, S.; Tio, Z.K.; Lim, E.Y.; Sun, J.; Zhang, J.; Wang, C.-H.; et al. Bioaugmentation of Methanosarcina thermophila grown on biochar particles during semi-continuous thermophilic food waste anaerobic digestion under two different bioaugmentation regimes. Bioresour. Technol. 2022, 360, 127590. [Google Scholar] [CrossRef]
- Liu, M.H.; Hsiao, C.M.; Wang, Y.S.; Chen, W.Y.; Hung, J.M. Tandem modified Fenton oxidation and bioremediation to degrade lubricant-contaminated soil. Int. Biodeterior. Biodegrad. 2019, 143, 104738. [Google Scholar] [CrossRef]
- Teng, T.; Liang, J.; Zhu, J.; Jin, P.; Zhang, D. Altered active pyrene degraders in biosurfactant-assisted bioaugmentation as revealed by RNA stable isotope probing. Environ. Pollut. 2022, 313, 120192. [Google Scholar] [CrossRef]
- Mansouri, A.; Cregut, M.; Jouanneau, S.; Thouand, G.; Durand, M.J. Evaluation of Biomonitoring Strategies to Assess Performance of a Bioremediation Bioprocess. Sustainability 2022, 14, 10932. [Google Scholar] [CrossRef]
- Vasilyeva, G.; Mikhedova, E.; Zinnatshina, L.; Strijakova, E.; Akhmetov, L.; Sushkova, S.; Ortega-Calvo, J.-J. Use of natural sorbents for accelerated bioremediation of grey forest soil contaminated with crude oil. Sci. Total Environ. 2022, 850, 157952. [Google Scholar] [CrossRef] [PubMed]
- Gibert, O.; Sánchez, D.; Cortina, J.L. Removal of nitrate and pesticides from groundwater by nano zero-valent iron injection pulses under biostimulation and bioaugmentation scenarios in continuous-flow packed soil columns. J. Environ. Manag. 2022, 321, 115965. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Tian, K.; Meng, F.; Li, S.; Li, H.; Yu, Y.; Qiu, Q.; Chang, M.; Huo, H. Effects of Bisphenol A Stress on Activated Sludge in Sequential Batch Reactors and Functional Recovery. Appl. Sci. 2022, 12, 8026. [Google Scholar] [CrossRef]
- Tran, H.T.; Nguyen, M.K.; Hoang, H.G.; Hutchison, J.M.; Vu, C.T. Composting and green technologies for remediation of phthalate (PAE)-contaminated soil: Current status and future perspectives. Chemosphere 2022, 307, 135989. [Google Scholar] [CrossRef]
- Jabbar, N.M.; Alardhi, S.M.; Mohammed, A.K.; Salih, I.K.; Albayati, T.M. Challenges in the implementation of bioremediation processes in petroleum-contaminated soils: A review. Environ. Nanotechnol. Monit. Manag. 2022, 18, 100694. [Google Scholar] [CrossRef]

| Culture for Bioaugmentation | Type of Contaminant | Removal Efficiency (RE), Environment, Co-Cultures | Reference | |
|---|---|---|---|---|
| Genus | Species/ Strain | |||
| Bacillus | B. megaterium and B. safensis | Chlorpyrifos (CLP) | RE in solution: 99% of CLP in a solution with an initial concentration of 10 mg/L after 60 days. RE in soil: 61–65% after 100 days. | [25] |
| B. firmus | Phenanthrene | RE: 1.73 mg/kg soil day for 56 days. Soil, anaerobic nitrate-reducing environment. | [26] | |
| B. subtilis, B. licheniformis, B. megaterium, B. cereus | Nitrogen in WW | RE: 92% of nitrate at the laboratory and 62% outdoors. A plug flow system; Arthrobacter sp., Acinetobacter parafneus, Corynebacterium sp., and Streptomyces globisporus | [27] | |
| B. megaterium | Nitrogen in WW | RE: 62.76% after 29 d. Secondary salinized soil, pot experiment. Improved NO3- removal rate. | [28] | |
| Bacillus sp. | Decachlorobiphenyl | RE: up to 10.51% after 21 days. Initial conc. 200 mg/kg soil. Staphylococcus sp. and Acinetobacter sp. Consortia better than individual strains. | [29] | |
| Bacillus spp. and B. aryabhattai | Aquaculture WW | Algal–bacterial bioflocs and microbe–rice bran complexes, Scenedesmus dimorphus and Chlorella sp. 1:1 | [30] | |
| B. paramycoides | Sulfamethoxazole (SMX) and Zn2+ | Biochar-immobilized. After five rounds of reuse, RE: 43.24% for SMX and 50.34% for Zn2+ | [31] | |
| B. safensis | Secondary composting | Indoleacetic acid (IAA)-production, assimilation of soluble salt, condensation and aromatization of humus, accumulation of dissolved organic nitrogen and carbon. Corynebacterium stationis subsp. safensis. | [32] | |
| Pseudomonas | Pseudomonas sp. | Sulphide | RE: two-fold increase of elemental sulfur generation. Establishment of a stronger biofilm structure. Granular sludge bed reactor. Arcobacter, Azoarcus | [33] |
| P. mendocina, P. putida | Nitrogen removal in petroleum WW | RE: 92.4% of NH4 +-N; 79.8% of total nitrogen. Heterotrophic nitrification, aerobic denitrification. Brucella sp., Paracoccus sp. | [34] | |
| P. plecoglossicida | Crude oil | 76.7% ability to degrade crude oil in a liquid broth. 14 days. | [11] | |
| P. stutzeri | Glyphosate | RE: 53% and 79% for synthetic and real WW with 5 mg/L glyphosate. Continuous photobioreactors. Acclimation to glyphosate from 5 to 50 mg L-1. Comamonas odontotermitis; Sinomonas atrocyanea, Chlorella protothecoides. | [35] | |
| Pseudomonas sp. | Benzene, toluene, ethylbenzene, and p-xylene (BTEX) | RE: 20 mg/L in a mineral salt medium for 6 h. Variovorax paradoxus | [36] | |
| P. songnenensis | Oil-contaminated desert soil | RE: 73.6%, 69.3%, 50% and 50% in soils polluted with 1%, 10%, 20% and 30% oil, respectively, after 6 months. Actinotalea ferrariae, Arthrobacter ginsengisoli, Dietzia cinnamea, Dietzia papillomatosis | [37] | |
| Pseudomonas sp. | Fluoride | RE: 77.54%, 99.39%, and 67.25% for F-, NO3-, and Ca2+, respectively, for 8 h. Microbially induced calcium precipitation (MICP). Self-assembled fungus-flexible fibre composite microspheres. Phoma sp. | [38] | |
| Rhodococcus | R. ruber | Chloroxylenol (PCMX) | RE: 97% during 22–30 cycles. 25 mg/L PCMX within 180 min in activated sludge system. Aerobic 100 mL batch, 4% inoculum (v/v). R. ruber became dominant within 30 cycles. Detoxify nitrification system. | [19] |
| R. ruber | Quinoline and its toxic intermediate 2-hydroxyl quinoline (2-HQ) | RE: for quinoline, 100% for 1.5 h. For 2-HQ, max. appearance of 0.025 mM within 0.5 h, 100% degradation after 1 h. Aerobic 100 mL batch, 15% inoculum (v/v), initial concentration of quinoline of 0.25 mM. Accelerated nitrification and enriched Nitrospira. | [20] | |
| R. erythropolis (Genetically engineered expressing Nirs and AMO (rRho-NM).) | Nanofiltration concentrate of landfill leachate | RE: total organic carbon 63–89%; NH4–N 72–82%; total nitrogen 63–87%; chemical oxygen demand 81–95%, dependent on treatment mode. Integrated system of advanced oxidation processes cooperated with rRho-NM. Bioaugmentation to the aerobic fluidized reactor (2 L), inoculum 106/mL. | [21] | |
| R. pyridinivorans | Di-(2-ethylhexyl)phthalate (DEHP) | RE: 89.94% of DEHP within 84 h. Initial DEHP conc. 5 mg/L DEHP in 10 mL municipal WW (batch). Aerobic denitrifying phosphate-accumulating bacterial strain RL-GZ01. | [39] | |
| R. biphenylivorans | Biphenyl and polychlorinated biphenyl (PCB) 31 | RE: biphenyl 96% within 5 days, PCB31 92% for 3 days. Initial conc. biphenyl 500 mg/L, aerobic 5 mL batch cultivation, pH 7.0. | [40] | |
| Rhodococcus sp. | 3-Methylindole (skatole) | RE: >99% for 24 h. Initial skatole conc. 60 mg/L. Degradation performance in consortium. Inoculum 2%. | [41] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muter, O. Current Trends in Bioaugmentation Tools for Bioremediation: A Critical Review of Advances and Knowledge Gaps. Microorganisms 2023, 11, 710. https://doi.org/10.3390/microorganisms11030710
Muter O. Current Trends in Bioaugmentation Tools for Bioremediation: A Critical Review of Advances and Knowledge Gaps. Microorganisms. 2023; 11(3):710. https://doi.org/10.3390/microorganisms11030710
Chicago/Turabian StyleMuter, Olga. 2023. "Current Trends in Bioaugmentation Tools for Bioremediation: A Critical Review of Advances and Knowledge Gaps" Microorganisms 11, no. 3: 710. https://doi.org/10.3390/microorganisms11030710
APA StyleMuter, O. (2023). Current Trends in Bioaugmentation Tools for Bioremediation: A Critical Review of Advances and Knowledge Gaps. Microorganisms, 11(3), 710. https://doi.org/10.3390/microorganisms11030710