
Screening of Native Trichoderma Species for Nickel and Copper Bioremediation Potential Determined by FTIR and XRF
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Soil Samples
2.2. Determination of DTPA-Extractable Metal Concentration in Soil Samples
2.3. Selection of Nickel- and Copper-Tolerant Isolates and Metal Tolerance Test of Trichoderma Spp.
2.4. Morphological Characterization
2.5. Fourier-Transform Infrared Spectroscopy (FTIR) and X-ray Fluorescence Spectroscopy (XRF) Analysis of Fungal Cells
2.6. Statistical Analysis
3. Results and Discussion
3.1. Micro Morphological Characteristics
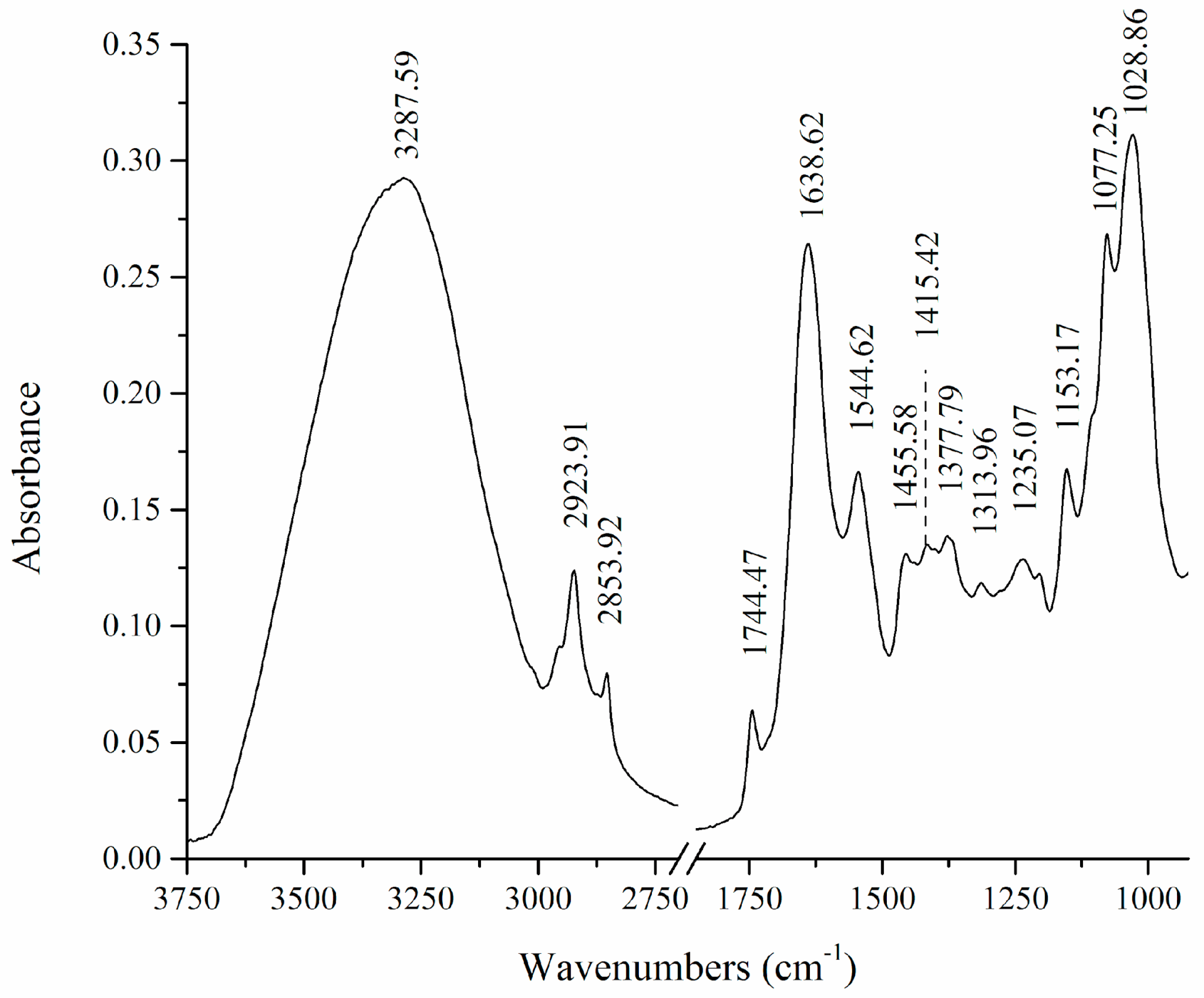
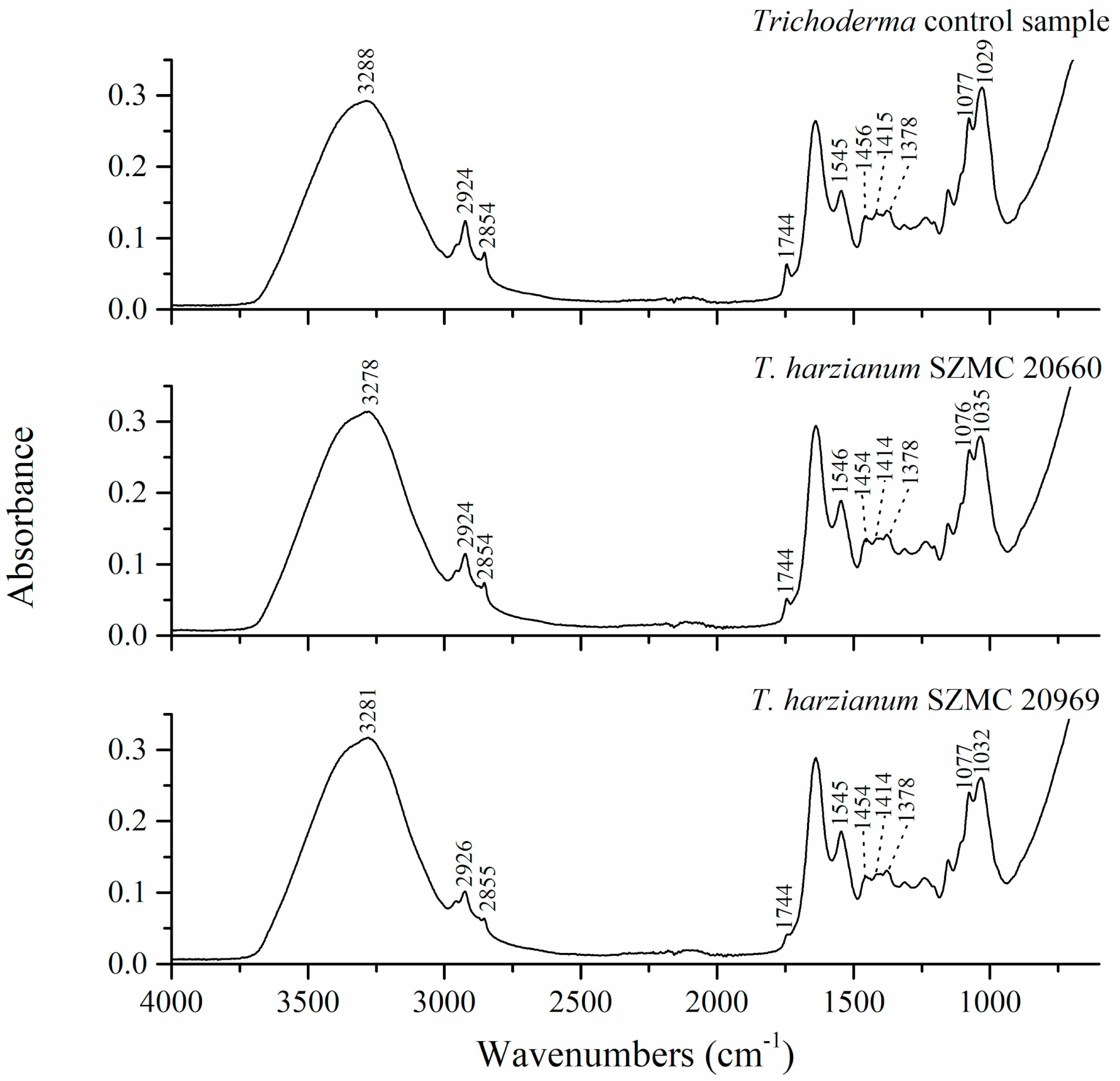
3.2. FTIR Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cecchi, G.; Marescotti, P.; Di Piazza, S.; Zotti, M. Native fungi as metal remediators: Silver myco-accumulation from metal contaminated waste-rock dumps (Libiola Mine, Italy). J. Environ. Sci. Health B 2017, 52, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Ramamurthy, A.S.; Memarian, R. Phytoremediation of mixed soil contaminants. Water Air Soil Pollut. 2012, 223, 511–518. [Google Scholar] [CrossRef]
- Binish, M.B.; Binu, P.; Gopikrishna, V.G.; Mohan, M. Potential of anaerobic bacteria in bioremediation of metal-contaminated marine and estuarine environment. In Microbial Biodegradation and Bioremediation; Elsevier: Amsterdam, The Netherlands, 2022; pp. 305–326. [Google Scholar]
- Oyetibo, G.O.; Miyauchi, K.; Huang, Y.; Chien, M.F.; Ilori, M.O.; Amund, O.O.; Endo, G. Biotechnological remedies for the estuarine environment polluted with heavy metals and persistent organic pollutants. Int. Biodeterior. Biodegrad. 2017, 119, 614–625. [Google Scholar] [CrossRef]
- Liu, S.H.; Guang, M.Z.; Qiu, Y.N.; Yang, L.; Lu, Z.; Lu, H.J.; Xiao, T.; Piao, X.; Chen, Z.; Min, C. Bioremediation mechanisms of combined pollution of PAHs and heavy metals by bacteria and fungi. Bioresour. Technol. 2017, 224, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Taghavi, S.; Lesaulnier, C.; Monchy, S.; Wattiez, R.; Mergeay, M.; van der Lelie, D. Lead (II) resistance in Cupriavidus metallidurans CH34: Interplay between plasmid and chromosomally-located functions. Anton. Leeuw. Int. J. G 2009, 96, 171. [Google Scholar] [CrossRef]
- Maldonado, J.; Diestra, E.; Huang, L.; Domènech, A.M.; Villagrasa, E.; Puyen, Z.M.; Solé, A. Isolation and identification of a bacterium with high tolerance to lead and copper from a marine microbial mat in Spain. Ann. Microbiol. 2010, 60, 113120. [Google Scholar] [CrossRef]
- Naik, M.M.; Dubey, S.K. Lead-enhanced siderophore production and alteration in cell morphology in a Pb-resistant Pseudomonas aeruginosa strain 4EA. Curr. Microbiol. 2011, 62, 409414. [Google Scholar] [CrossRef]
- Ayangbenro, A.S.; Babalola, O.O. A new strategy for heavy metal polluted environments: A review of microbial biosorbents. Int. J. Environ. Res. Public Health 2017, 14, 94. [Google Scholar] [CrossRef]
- Zhang, H.; Yuan, X.; Xiong, T.; Wang, H.; Jiang, L. Bioremediation of co-contaminated soil with heavy metals and pesticides: Influence factors, mechanisms and evaluation methods. J. Chem. Eng. 2020, 398, 125657. [Google Scholar] [CrossRef]
- Dixit, R.; Malaviya, D.; Pandiyan, K.; Singh, U.B.; Sahu, A.; Shukla, R.; Singh, B.; Rai, J.; Sharma, P.; Lade, H.; et al. Bioremediation of heavy metals from soil and aquatic environment: An overview of principles and criteria of fundamental processes. Sustainability 2015, 7, 2189–2212. [Google Scholar] [CrossRef] [Green Version]
- Tufail, M.A.; Iltaf, J.; Zaheer, T.; Tariq, L.; Amir, M.B.; Fatima, R.; Asbat, A.; Kabeer, T.; Fahad, M.; Naeem, H.; et al. Recent advances in bioremediation of heavy metals and persistent organic pollutants: A review. Sci. Total Environ. 2022, 850, 157961. [Google Scholar] [CrossRef] [PubMed]
- Su, S.M.; Zeng, X.B.; Li, L.F.; Duan, R.; Bai, L.Y.; Li, A.G.; Jiang, S. Arsenate reduction and methylation in the cells of Trichoderma asperellum SM-12F1, Penicillium janthinellum SM-12F4, and Fusarium oxysporum CZ-8F1 investigated with X-ray absorption near edge structure. J. Hazard. Mater. 2012, 243, 364–367. [Google Scholar] [CrossRef] [PubMed]
- Oladipo, O.G.; Awotoye, O.O.; Olayinka, A.; Bezuidenhout, C.C.; Maboeta, M.S. Heavy metal tolerance traits of filamentous fungi isolated from gold and gemstone mining sites. Braz. J. Microbiol. 2018, 49, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Ramrakhiani, L.; Majumder, R.; Khowala, S. Removal of hexavalent chromium by heat inactivated fungal biomass of Termitomyces clypeatus: Surface characterization and mechanism of biosorption. J. Chem. Eng. 2011, 171, 1060–1068. [Google Scholar] [CrossRef]
- Iram, S.; Shabbir, R.; Zafar, H.; Javaid, M. Biosorption and bioaccumulation of copper and lead by heavy metal-resistant fungal isolates. Arab. J. Sci. Eng. 2015, 40, 1867–1873. [Google Scholar] [CrossRef]
- Amirnia, S.; Ray, M.B.; Margaritis, A. Heavy metals removal from aqueous solutions using Saccharomyces cerevisiae in a novel continuous bioreactor–biosorption system. J. Chem. Eng. 2015, 264, 863–872. [Google Scholar] [CrossRef]
- Thakare, M.; Sarma, H.; Datar, S.; Roy, A.; Pawar, P.; Gupta, K.; Pandit, S.; Prasad, R. Understanding the holistic approach to plant-microbe remediation technologies for removing heavy metals and radionuclides from soil. Curr. Biotechnol. 2021, 3, 84–98. [Google Scholar] [CrossRef]
- Martins, L.R.; Lyra, F.H.; Rugani, M.M.; Takahashi, J.A. Bioremediation of metallic ions by eight Penicillium species. Environ. Eng. 2016, 142, C4015007. [Google Scholar] [CrossRef]
- Kaewdoung, B.; Thanawat, S.; Geoffrey, M.G.; Anthony, W.; Prakitsin, S. Heavy metal tolerance and biotransformation of toxic metal compounds by new isolates of wood-rotting fungi from Thailand. Geomicrobiol. J. 2016, 33, 283–288. [Google Scholar] [CrossRef]
- Zhao, W.W.; Zhu, G.Q.; Andrew, J.D.; Qi, C.; Hong, Y.M.; Peng, Z.; Jian, L.; Ma, X.K. Removal and biomineralization of Pb2+ in water by fungus Phanerochaete chrysoporium. J. Clean. Prod. 2020, 260, 120980. [Google Scholar] [CrossRef]
- Timková, I.; Sedláková-Kaduková, J.; Pristaš, P. Biosorption and bioaccumulation abilities of actinomycetes/streptomycetes isolated from metal contaminated sites. Separations 2018, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Ayele, A.; Godeto, Y.G. Bioremediation of chromium by microorganisms and its mechanisms related to functional groups. J. Chem. 2021, 2021, 1–21. [Google Scholar] [CrossRef]
- Zhang, D.; Yin, C.; Abbas, N.; Mao, Z.; Zhang, Y. Multiple heavy metal tolerance and removal by an earthworm gut fungus Trichoderma brevicompactum QYCD-6. Sci. Rep. 2020, 10, 6940. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, D.; Sheng, F.; Qing, H. Adsorption characteristics of Copper (II), Zinc (II) and Mercury (II) by four kinds of immobilized fungi residues. Ecotoxicol. Environ. Saf. 2018, 147, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Dou, K.; Wang, P.; Chenthamara, K.; Chen, J.; Druzhinina, I.S. The Current State of Trichoderma Taxonomy and Species Identification. In Advances in Trichoderma Biology for Agricultural Applications; Springer: Berlin/Heidelberg, Germany, 2022; pp. 3–35. [Google Scholar]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The current status of its application in agriculture for the biocontrol of fungal phytopathogens and stimulation of plant growth. Int. J. Mol. Sci. 2022, 23, 2329. [Google Scholar] [CrossRef]
- Alothman, Z.A.; Bahkali, A.H.; Elgorban, A.M.; Al-Otaibi, M.S.; Ghfar, A.A.; Gabr, S.A.; Wabaidur, S.M.; Habila, M.A.; Ahmed, A.Y.B.H. Bioremediation of explosive TNT by Trichoderma viride. Molecules 2020, 25, 1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Barrera, L.; Cristiani-Urbina, E. Hexavalent chromium removal by a Trichoderma inhamatum fungal strain isolated from tannery effluent. Water Air Soil Pollut. 2008, 187, 327–336. [Google Scholar] [CrossRef]
- Nongmaithem, N.; Roy, A.; Bhattacharya, P.M. Screening of Trichoderma isolates for their potential of biosorption of nickel and cadmium. Braz. J. Microbiol. 2016, 47, 305–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiquee, S.; Aishah, S.N.; Azad, S.A.; Shafawati, S.N.; Naher, L. Tolerance and biosorption capacity of Zn2+, Pb2+, Ni3+ and Cu2+ by filamentous fungi (Trichoderma harzianum, T. aureoviride and T. virens). Adv. Biosci. Biotechnol. 2013, 4, 570–583. [Google Scholar] [CrossRef] [Green Version]
- Maldaner, J.; Steffen, G.P.K.; Missio, E.L.; Saldanha, C.W.; de Morais, R.M.; Nicoloso, F.T. Tolerance of Trichoderma isolates to increasing concentrations of heavy metals. Int. J. Environ. Stud. 2021, 78, 185–197. [Google Scholar] [CrossRef]
- Panagos, P.; Cristiano, B.; Emanuele, L.; Arwyn, J.; Pasquale, B.; Simone, S.; Alberto, O.; Luca, M. Potential sources of anthropogenic copper inputs to European agricultural soils. Sustainability 2018, 10, 2380. [Google Scholar] [CrossRef] [Green Version]
- Racić, G.; Körmöczi, P.; Kredics, L.; Raičević, V.; Mutavdžić, B.; Vrvić, M.M.; Panković, D. Effect of the edaphic factors and metal content in soil on the diversity of Trichoderma spp. Environ. Sci. Pollut. Res. 2017, 24, 3375–3386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelegrino, M.T.; Kohatsu, M.Y.; Seabra, A.B.; Monteiro, L.R.; Gomes, D.G.; Oliveira, H.C.; Rolim, W.R.; de Jesus, T.A.; Batista, B.L.; Lange, C.N. Effects of copper oxide nanoparticles on growth of lettuce (Lactuca sativa L.) seedlings and possible implications of nitric oxide in their antioxidative defense. Environ. Monit. Asses. 2020, 192, 1–14. [Google Scholar] [CrossRef]
- Chen, M.; Ding, S.; Gao, S.; Xu, S.; Yang, C.; Wu, Y.; Gong, M.; Wang, D.; Wang, Y. Long-term effects of sediment dredging on controlling cobalt, zinc, and nickel contamination determined by chemical fractionation and passive sampling. Chemosphere 2019, 220, 476–485. [Google Scholar] [CrossRef]
- Lindsay, W.L.; Norvell, W. Development of a DTPA soil test for zinc, iron, manganese, and copper. Soil Sci. Soc. Am. J. 1978, 42, 421–428. [Google Scholar] [CrossRef]
- King, A.D., Jr.; Hocking, A.D.; Pitt, J.I. Dichloran-rose bengal medium for enumeration and isolation of molds from foods. Appl. Environ. Microbiol. 1979, 37, 959–964. [Google Scholar] [CrossRef] [Green Version]
- Van Volkshuisvesting, M. Circular on Target Values and Intervention Values for Soil Remediation-Dutch Target and Intervention Values; Netherlands Government Gazette: Amsterdam, The Netherlands, 2000.
- Glen, K.; Boligłowa, E. Evaluation of the Fungistatic Activity of Plant Extracts In Vitro Tests. J. Agric. Eng. Res. 2012, 57, 104–109. [Google Scholar]
- Rifai, M.A. A revision of the genus Trichoderma. Mycol. Papers 1969, 116, 1–56. [Google Scholar]
- Bissett, J. A revision of the genus Trichoderma. II. Infrageneric classification. Can. J. Bot. 1991, 69, 2357–2372. [Google Scholar] [CrossRef]
- Rahman, M.A.; Halfar, J. First evidence of chitin in calcified coralline algae: New insights into the calcification process of Clathromorphum compactum. Sci. Rep. 2014, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lao, C.; Luo, L.; Shen, Y.; Zhu, S. Micro X-ray Fluorescence and X-ray Absorption near Edge Structure Analysis of Heavy Metals in Micro-organism. X-Ray Fluoresc. Biol. Sci. Princ. Instrum. Appl. 2022, 73–79. [Google Scholar] [CrossRef]
- Reith, F.; Etschmann, B.; Grosse, C.; Moors, H.; Benotmane, M.A.; Monsieurs, P.; Grass, G.; Doonan, C.; Vogt, S.; Lai, B.; et al. Mechanisms of gold biomineralization in the bacterium Cupriavidus metallidurans. Proc. Natl. Acad. Sci. USA 2009, 106, 17757–17762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kredics, L.; Dóczi, I.; Antal, Z.; Manczinger, L. Effect of heavy metals on growth and extracellular enzyme activities of mycoparasitic Trichoderma strains. Bull. Environ. Contam. Toxicol. 2001, 66, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Sahu, A.; Mandal, A.; Thakur, J.; Manna, M.C.; Rao, A.S. Exploring bioaccumulation efficacy of Trichoderma viride: An alternative bioremediation of cadmium and lead. Natl. Acad. Sci. Lett. 2012, 35, 299–302. [Google Scholar] [CrossRef]
- Kacprzak, M.J.; Rosikon, K.; Fijalkowski, K.; Grobelak, A. The effect of Trichoderma on heavy metal mobility and uptake by Miscanthus giganteus, Salix sp., Phalaris arundinacea, and Panicum virgatum. Appl. Environ. Soil Sci. 2014, 2014, 506142. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, P.; Singh, P.C.; Mishra, A.; Chauhan, P.S.; Dwivedi, S.; Bais, R.T.; Tripathi, R.D. Trichoderma: A potential bioremediator for environmental clean up. Clean Technol. Environ. 2013, 15, 541–550. [Google Scholar] [CrossRef]
- Stebbing, A.R.D. The kinetics of growth control in a colonial hydroid. J. Mar. Biol. Assoc. UK 1981, 61, 35–63. [Google Scholar] [CrossRef]
- Fomina, M.; Gadd, G.M. Metal sorption by biomass of melanin-producing fungi grown in clay-containing medium. J. Chem. Technol. Biotechnol. 2003, 78, 23–34. [Google Scholar] [CrossRef]
- De Padua, J.C.; dela Cruz, T.E.E. Isolation and characterization of nickel-tolerant Trichoderma strains from marine and terrestrial environments. J. Fungi 2021, 7, 591. [Google Scholar] [CrossRef]
- Küpper, V.; Steiner, U.; Kortekamp, A. Trichoderma species isolated from grapevine with tolerance towards common copper fungicides used in viticulture for plant protection. Pest Manag. Sci. 2022, 78, 3266–3276. [Google Scholar] [CrossRef]
- Erukhimovitch, V.; Pavlov, V.; Talyshinsky, M.; Souprun, Y.; Huleihel, M. FTIR microscopy as a method for identification of bacterial and fungal infections. J. Pharm. Biomed. Anal. 2005, 37, 1105–1108. [Google Scholar] [CrossRef] [PubMed]
- Naumann, A. A novel procedure for strain classification of fungal mycelium by cluster and artificial neural network analysis of Fourier transform infrared (FTIR) spectra. Analyst 2009, 134, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Forfang, K.; Zimmermann, B.; Kosa, G.; Kohler, A.; Shapaval, V. FTIR spectroscopy for evaluation and monitoring of lipid extraction efficiency for oleaginous fungi. PLoS ONE 2017, 12, e0170611. [Google Scholar] [CrossRef] [Green Version]
- Szeghalmi, A.; Kaminskyj, S.; Gough, K.M. A synchrotron FTIR microspectroscopy investigation of fungal hyphae grown under optimal and stressed conditions. Anal. Bioanal. Chem. 2007, 387, 1779–1789. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Cai, P.; Chen, W.; Liang, W.; Hong, Z.; Huang, Q. Impact of cell wall structure on the behavior of bacterial cells in the binding of copper and cadmium. Colloids Surf. A Physicochem. Eng. Asp. 2009, 347, 50–55. [Google Scholar] [CrossRef]
- Pugazhendhi, A.; Ranganathan, K.; Kaliannan, T. Biosorptive removal of copper (II) by Bacillus cereus isolated from contaminated soil of electroplating industry in India. Water Air Soil Pollut. 2018, 229, 1–9. [Google Scholar] [CrossRef]
- Kumar, V.; Dwivedi, S.K. A review on accessible techniques for removal of hexavalent Chromium and divalent Nickel from industrial wastewater: Recent research and future outlook. J. Clean. Prod. 2021, 295, 126229. [Google Scholar] [CrossRef]
- Tu, C.; Liu, Y.; Wei, J.; Li, L.; Scheckel, K.G.; Luo, Y. Characterization and mechanism of copper biosorption by a highly copper-resistant fungal strain isolated from copper-polluted acidic orchard soil. Environ. Sci. Pollut. Res. 2018, 25, 24965–24974. [Google Scholar] [CrossRef]
- Sujatha, P.; Kalarani, V.; Kumar, B.N. Effective biosorption of nickel (II) from aqueous solutions using Trichoderma viride. J. Chem. 2013, 2013, 716098. [Google Scholar] [CrossRef] [Green Version]
- Waern, J.B.; Harris, H.H.; Lai, B.; Cai, Z.; Harding, M.M.; Dillon, C.T. Intracellular mapping of the distribution of metals derived from the antitumor metallocenes. J. Biol. Inorg. Chem. 2005, 10, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Cheow, Y.L.; Ng, S.L.; Ting, A.S.Y. Mechanisms for metal removal established via electron microscopy and spectroscopy: A case study on metal tolerant fungi Penicillium simplicissimum. J. Hazard. Mater. 2019, 362, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Isar, J.; Saran, S.; Saxena, R.K. Bioaccumulation of copper by Trichoderma viride. Bioresour. Technol. 2006, 97, 1018–1025. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, M.; Yap, C.K.; Abdullah, F.; Tan, S.G. Trichoderma atroviride as a bioremediator of Cu pollution: An in vitro study. Toxicol. Environ. Chem. 2009, 91, 1305–1314. [Google Scholar] [CrossRef]
- Palanivel, T.M.; Sivakumar, N.; Al-Ansari, A.; Victor, R. Bioremediation of copper by active cells of Pseudomonas stutzeri LA3 isolated from an abandoned copper mine soil. J. Environ. Manag. 2020, 253, 109706. [Google Scholar] [CrossRef]
- Prasad, G.; Kumar, V.; Dwivedi, S.K. Antifungal activity of some selected medicinal plants against Fusarium solani causing wilt and rot in Pearl millet. AJBS 2018, 13, 21–27. [Google Scholar] [CrossRef]
- Kumar, V.; Dwivedi, S.K. Hexavalent chromium stress response, reduction capability and bioremediation potential of Trichoderma spp. isolated from electroplating wastewater. Ecotoxicol. Environ. Saf. 2019, 185, 109734. [Google Scholar] [CrossRef]
- Raklami, A.; Meddich, A.; Oufdou, K.; Baslam, M. Plants—Microorganisms-based bioremediation for heavy metal cleanup: Recent developments, phytoremediation techniques, regulation mechanisms, and molecular responses. Int. J. Mol. Sci. 2022, 23, 5031. [Google Scholar] [CrossRef]
- Gonzalez Henao, S.; Ghneim-Herrera, T. Heavy metals in soils and the remediation potential of bacteria associated with the plant microbiome. Front. Environ. Sci. 2021, 9, 15. [Google Scholar] [CrossRef]
- Mushtaq, S.; Bareen, F.E.; Tayyeb, A.; Nazir, A. Autochthonous strains of Trichoderma isolated from tannery solid waste improve phytoextraction potential of heavy metals by sunflower. Int. J. Phytoremediat. 2022; 1–20, online ahead of print. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sample Number | Cd (mg/kg) | Cr (mg/kg) | Co (mg/kg) | Cu (mg/kg) | Pb (mg/kg) | Mn (mg/kg) | Ni (mg/kg) | Zn (mg/kg) | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Vineyard | 0.162 ± 0.012 | 0.019 ± 0.000 | 0.066 ± 0.002 | 19.684 ± 1.024 | 1.985 ± 0.083 | 18.268 ± 0.384 | 0.391 ± 0.038 | 5.026 ± 0.226 |
| 2 | Vineyard | 0.141 ± 0.007 | 0.019 ± 0.001 | 0.083 ± 0.003 | 16.718 ± 0.953 | 1.965 ± 0.100 | 20.122 ± 0.563 | 0.413 ± 0.032 | 4.348 ± 0.209 |
| 3 | Vineyard | 0.097 ± 0.008 | 0.018 ± 0.001 | 0.094 ± 0.002 | 11.818 ± 0.508 | 1.554 ± 0.071 | 22.970 ± 0.528 | 0.311 ± 0.026 | 2.294 ± 0.115 |
| 4 | Vineyard | 0.099 ± 0.005 | 0.017 ± 0.001 | 0.082 ± 0.002 | 12.466 ± 0.362 | 1.658 ± 0.066 | 21.914 ± 0.636 | 0.281 ± 0.019 | 2.166 ± 0.109 |
| 5 | Agricultural soil | 0.067 ± 0.002 | 0.018 ± 0.000 | 0.093 ± 0.003 | 1.507 ± 0.077 | 1.142 ± 0.061 | 30.554 ± 0.552 | 0.944 ± 0.072 | 1.023 ± 0.042 |
| 6 | Agricultural soil | 0.062 ± 0.003 | 0.018 ± 0.000 | 0.072 ± 0.002 | 1.087 ± 0.081 | 1.023 ± 0.052 | 26.736 ± 0.668 | 0.871 ± 0.081 | 0.994 ± 0.039 |
| 7 | Agricultural soil | 0.102 ± 0.004 | 0.142 ± 0.004 | 0.049 ± 0.001 | 3.552 ± 0.238 | 3.536 ± 0.163 | 8.842 ± 0.141 | 1.613 ± 0.147 | 1.268 ± 0.062 |
| 8 | Agricultural soil | 0.116 ± 0.005 | 0.095 ± 0.003 | 0.047 ± 0.002 | 3.550 ± 0.090 | 3.066 ± 0.123 | 9.428 ± 0.226 | 1.981 ± 0.184 | 1.315 ± 0.075 |
| 9 | Forest soil | 0.052 ± 0.002 | 0.024 ± 0.001 | 0.044 ± 0.002 | 1.837 ± 0.063 | 0.816 ± 0.026 | 10.228 ± 0.277 | 0.290 ± 0.016 | 0.667 ± 0.031 |
| 10 | Forest soil | 0.035 ± 0.003 | 0.026 ± 0.000 | 0.052 ± 0.001 | 1.472 ± 0.059 | 0.661 ± 0.036 | 9.158 ± 0.148 | 0.232 ± 0.014 | 0.471 ± 0.029 |
| 11 | Forest soil | 0.129 ± 0.006 | 0.237 ± 0.005 | 2.130 ± 0.062 | 1.114 ± 0.033 | 2.328 ± 0.109 | 54.606 ± 0.983 | 106.056 ± 8.062 | 2.722 ± 0.128 |
| 12 | Forest soil | 0.011 ± 0.001 | 0.491 ± 0.013 | 1.230 ± 0.038 | 0.625 ± 0.033 | 8.257 ± 0.330 | 24.912 ± 0.473 | 6.747 ± 0.614 | 3.001 ± 0.159 |
| 13 | Forest soil | 0.048 ± 0.002 | 0.305 ± 0.007 | 1.173 ± 0.028 | 0.556 ± 0.029 | 6.657 ± 0.280 | 13.488 ± 0.351 | 5.883 ± 0.565 | 2.383 ± 0.101 |
| 14 | Agricultural soil | 0.069 ± 0.005 | 0.041 ± 0.001 | 0.084 ± 0.004 | 1.696 ± 0.086 | 1.982 ± 0.097 | 21.766 ± 0.588 | 1.182 ± 0.050 | 0.637 ± 0.020 |
| 15 | Forest soil | 0.067 ± 0.003 | 0.061 ± 0.002 | 0.078 ± 0.002 | 2.066 ± 0.077 | 2.678 ± 0.088 | 24.022 ± 1.081 | 1.541 ± 0.149 | 0.621 ± 0.051 |
| 16 | Forest soil | 0.046 ± 0.004 | 0.018 ± 0.001 | 0.077 ± 0.003 | 7.326 ± 0.300 | 1.196 ± 0.049 | 24.020 ± 0.576 | 0.684 ± 0.065 | 5.584 ± 0.123 |
| 17 | Forest soil | 0.047 ± 0.003 | 0.018 ± 0.001 | 0.066 ± 0.003 | 5.478 ± 0.186 | 1.110 ± 0.057 | 20.438 ± 0.593 | 0.679 ± 0.054 | 4.390 ± 0.320 |
| 18 | Forest soil | 0.209 ± 0.015 | 0.026 ± 0.001 | 0.074 ± 0.003 | 1.560 ± 0.083 | 1.868 ± 0.077 | 26.102 ± 0.548 | 1.080 ± 0.077 | 0.729 ± 0.041 |
| 19 | Forest soil | 0.065 ± 0.004 | 0.359 ± 0.009 | 0.076 ± 0.002 | 1.595 ± 0.062 | 1.746 ± 0.079 | 28.582 ± 1.003 | 1.112 ± 0.082 | 0.774 ± 0.038 |
| 20 | Forest soil | 0.071 ± 0.005 | 0.366 ± 0.011 | 0.037 ± 0.001 | 1.023 ± 0.053 | 1.359 ± 0.053 | 13.722 ± 0.563 | 0.942 ± 0.071 | 1.558 ± 0.064 |
| 21 | Forest soil | 0.054 ± 0.004 | 0.018 ± 0.000 | 0.070 ± 0.002 | 0.962 ± 0.036 | 1.631 ± 0.078 | 21.184 ± 0.657 | 0.769 ± 0.066 | 1.188 ± 0.056 |
| 22 | Forest soil | 0.044 ± 0.002 | 0.800 ± 0.033 | 0.206 ± 0.005 | 3.406 ± 0.143 | 2.260 ± 0.081 | 36.716 ± 1.248 | 4.592 ± 0.445 | 0.811 ± 0.048 |
| 23 | Forest soil | 0.061 ± 0.004 | 0.078 ± 0.002 | 0.032 ± 0.001 | 4.328 ± 0.234 | 2.216 ± 0.112 | 12.824 ± 0.321 | 1.358 ± 0.126 | 5.518 ± 0.386 |
| MIN | 0.011 ± 0.001 | 0.017 ± 0.001 | 0.032 ± 0.001 | 0.556 ± 0.029 | 0.661 ± 0.036 | 8.842 ± 0.141 | 0.232 ± 0.014 | 0.471 ± 0.029 | |
| MAX | 0.209 ± 0.015 | 0.800 ± 0.033 | 2.130 ± 0.062 | 19.684 ± 1.024 | 8.257 ± 0.330 | 54.606 ± 0.983 | 106.056 ± 8.062 | 5.584 ± 0.226 |
| Copper mg/L | Time | SZMC20660 T. harzianum | SZMC20969 T. harzianum |
|---|---|---|---|
| Colony diameter (cm) | |||
| Control | 48 h | 7.78 ± 0.19 a | 6.64 ± 0.99 ab |
| 30 mg/L | 6.94 ± 0.08 a | 7.54 ± 0.18 a | |
| 60 mg/L | 5.88 ± 0.46 b | 6.04 ± 0.28 b | |
| 120 mg/L | 1.24 ± 0.55 c | 2.06 ± 0.08 c | |
| 240 mg/L | 0.00 ± 0.00 d | 0.00 ± 0.00 d | |
| Control | 96 h | 8.46 ± 0.05 a | 8.48 ± 0.05 a |
| 30 mg/L | 8.46 ± 0.08 a | 8.44 ± 0.08 a | |
| 60 mg/L | 8.45 ± 0.07 a | 8.47 ± 0.04 a | |
| 120 mg/L | 6.04 ± 0.04 c | 7.12 ± 0.50 b | |
| 240 mg/L | 1.02 ± 0.02 d | 1.01 ± 0.05 d | |
| Control | 144 h | 8.50 ± 0.07 a | 8.52 ± 0.04 a |
| 30 mg/L | 8.50 ± 0.07 a | 8.48 ± 0.05 a | |
| 60 mg/L | 8.49 ± 0.05 a | 8.50 ± 0.08 a | |
| 120 mg/L | 8.46 ± 0.05 a | 8.51 ± 0.05 a | |
| 240 mg/L | 1.78 ± 0.17 b | 1.50 ± 0.06 c | |
| Nickel mg/L | Time | SZMC20664 T. longibrachiatum | SZMC20669 T. longibrachiatum | SZMC20665 T. longibrachiatum |
|---|---|---|---|---|
| Colony diameter (cm) | ||||
| Control | 48 h | 7.60 ± 0.25 ab | 7.52 ± 0.19 ab | 7.50 ± 0.35 ab |
| 30 mg/L | 7.70 ± 0.2 ab | 7.74 ± 0.25 ab | 7.83 ± 0.29 a | |
| 60 mg/L | 6.80 ± 0.20 c | 7.12 ± 0.20 bc | 6.88 ± 0.2 c | |
| 120 mg/L | 1.82 ± 0.31 d | 1.62 ± 0.37 d | 1.86 ± 0.71 d | |
| 240 mg/L | 0.06 ± 0.13 e | 0.00 ± 0.00 e | 0.00 ± 0.00 e | |
| Control | 96 h | 7.96 ± 0.20 a | 8.02 ± 0.22 a | 8.06 ± 0.23 a |
| 30 mg/L | 7.92 ± 0.21 a | 7.98 ± 0.17 a | 8.04 ± 0.16 a | |
| 60 mg/L | 7.94 ± 0.19 a | 8.01 ± 0.18 a | 8.04 ± 0.11 a | |
| 120 mg/L | 5.44 ± 0.32 b | 5.46 ± 0.39 b | 5.94 ± 0.48 b | |
| 240 mg/L | 0.00 ± 0.00 c | 0.00 ± 0.00 c | 0.00 ± 0.00 c | |
| Control | 144 h | 8.12 ± 0.31 ab | 8.28 ± 0.24 a | 8.32 ± 0.13 a |
| 30 mg/L | 8.08 ± 0.22 ab | 8.24 ± 0.16 a | 8.28 ± 0.22 a | |
| 60 mg/L | 8.07 ± 0.31 ab | 8.26 ± 0.21 a | 8.30 ± 0.12 a | |
| 120 mg/L | 7.68 ± 0.25 b | 8.00 ± 0.25 ab | 7.10 ± 0.20 c | |
| 240 mg/L | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 0.00 ± 0.00 d | |
| Ni | Cu | ||||||
|---|---|---|---|---|---|---|---|
| Control (Untreated Cells) | T. long. 22664 | T. long. 22669 | T. long. 22665 | T. harz. 20660 | T. harz. 20969 | Functional Group | Macromolecule |
| 3288 | 3285 | 3281 | 3279 | 3278 | 3281 | O-H, N-H | various |
| 2924, 2854 | 2925, 2854 | 2924, 2854 | 2927, 2856 | 2924, 2854 | 2926, 2855 | C-H | lipids |
| 1744 | 1744 | 1744 | / | 1744 | 1744 | C=O | lipids |
| 1639 | 1639 | 1637 | 1637 | 1638 | 1638 | Amide I C=O, C-N | proteins |
| 1545 | 1548 | 1545 | 1545 | 1546 | 1545 | Amide II N-H, C-N | proteins |
| 1456, 1415, 1378, 1314 | /, 1416, 1375, 1314 | /, 1418, 1376, 1314 | 1451, /, 1381, 1313 | 1454, 1414, 1378, 1313 | 1454, 1414, 1378, 1313 | O-H, CH2, CH3 | lipids, proteins |
| 1235 | 1240 | 1239 | 1241 | 1237 | 1240 | PO2, C-O-C | phosphate compounds, polysaccharides |
| 1153 | 1152 | 1154 | 1152 | 1154 | 1153 | C-O-C | polysaccharides |
| 1077 | 1078 | 1077 | 1078 | 1076 | 1077 | PO2, C-O | phosphate compounds, polysaccharides |
| 1029 | 1027 | 1031 | 1025 | 1035 | 1032 | C-O, C-C | lipids |
| Sample | Ni (%) | Cu (%) | |
|---|---|---|---|
| Control | / | / | |
| T. longibrachiatum SZMC 22665 | 0.249 | / | |
| 60 mg/L Ni | T. longibrachiatum SZMC 22669 | 0.171 | / |
| T. longibrachiatum SZMC22664 | 0.103 | / | |
| T. harzianum SZMC 20660 | / | 0.021 | |
| 60 mg/L Cu | T. harzianum SZMC 20969 | / | 0.070 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Racić, G.; Vukelić, I.; Kordić, B.; Radić, D.; Lazović, M.; Nešić, L.; Panković, D. Screening of Native Trichoderma Species for Nickel and Copper Bioremediation Potential Determined by FTIR and XRF. Microorganisms 2023, 11, 815. https://doi.org/10.3390/microorganisms11030815
Racić G, Vukelić I, Kordić B, Radić D, Lazović M, Nešić L, Panković D. Screening of Native Trichoderma Species for Nickel and Copper Bioremediation Potential Determined by FTIR and XRF. Microorganisms. 2023; 11(3):815. https://doi.org/10.3390/microorganisms11030815
Chicago/Turabian StyleRacić, Gordana, Igor Vukelić, Branko Kordić, Danka Radić, Milana Lazović, Ljiljana Nešić, and Dejana Panković. 2023. "Screening of Native Trichoderma Species for Nickel and Copper Bioremediation Potential Determined by FTIR and XRF" Microorganisms 11, no. 3: 815. https://doi.org/10.3390/microorganisms11030815
APA StyleRacić, G., Vukelić, I., Kordić, B., Radić, D., Lazović, M., Nešić, L., & Panković, D. (2023). Screening of Native Trichoderma Species for Nickel and Copper Bioremediation Potential Determined by FTIR and XRF. Microorganisms, 11(3), 815. https://doi.org/10.3390/microorganisms11030815