
Diverse Repertoire and Relationship of Exopolysaccharide Genes in Cold-Adapted Acinetobacter sp. CUI-P1 Revealed by Comparative Genome Analysis

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Screening and Isolation of EPS-Producing Bacteria
2.3. Extraction and Purification of EPS
2.4. Cryoprotectant Activity of EPSs
2.5. Emulsification Stability
2.6. Metal Tolerance of EPS-Producing Bacteria
2.7. Physical Characterization of EPS
2.8. Chemical Characterization of EPS
2.9. Genomic DNA Extraction, Genome Sequence, and Analysis
2.10. Statistical Analysis
3. Results
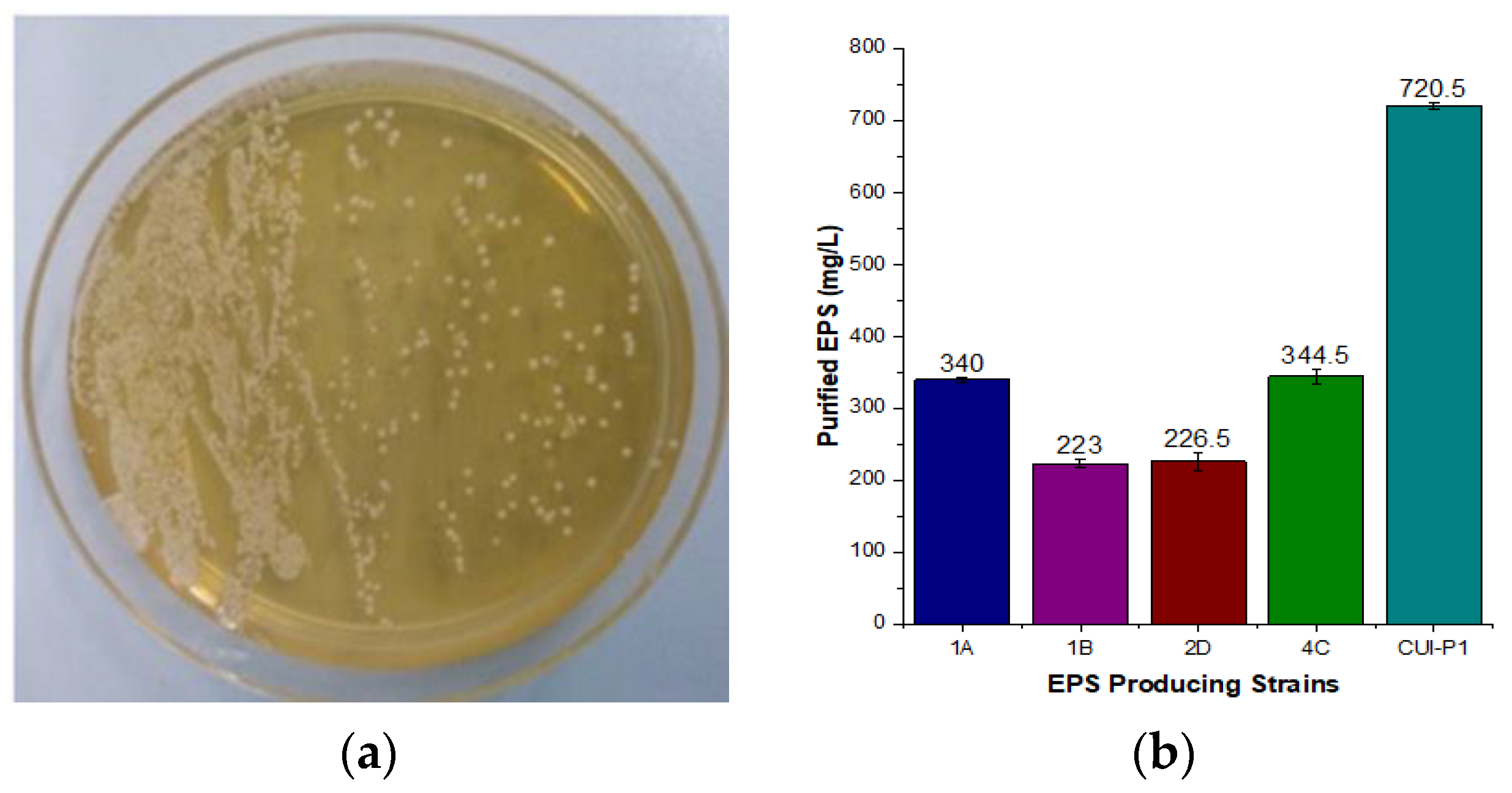
3.1. Screening of Isolated Strains for EPS Production
3.2. Identification of EPS-Producing Strains
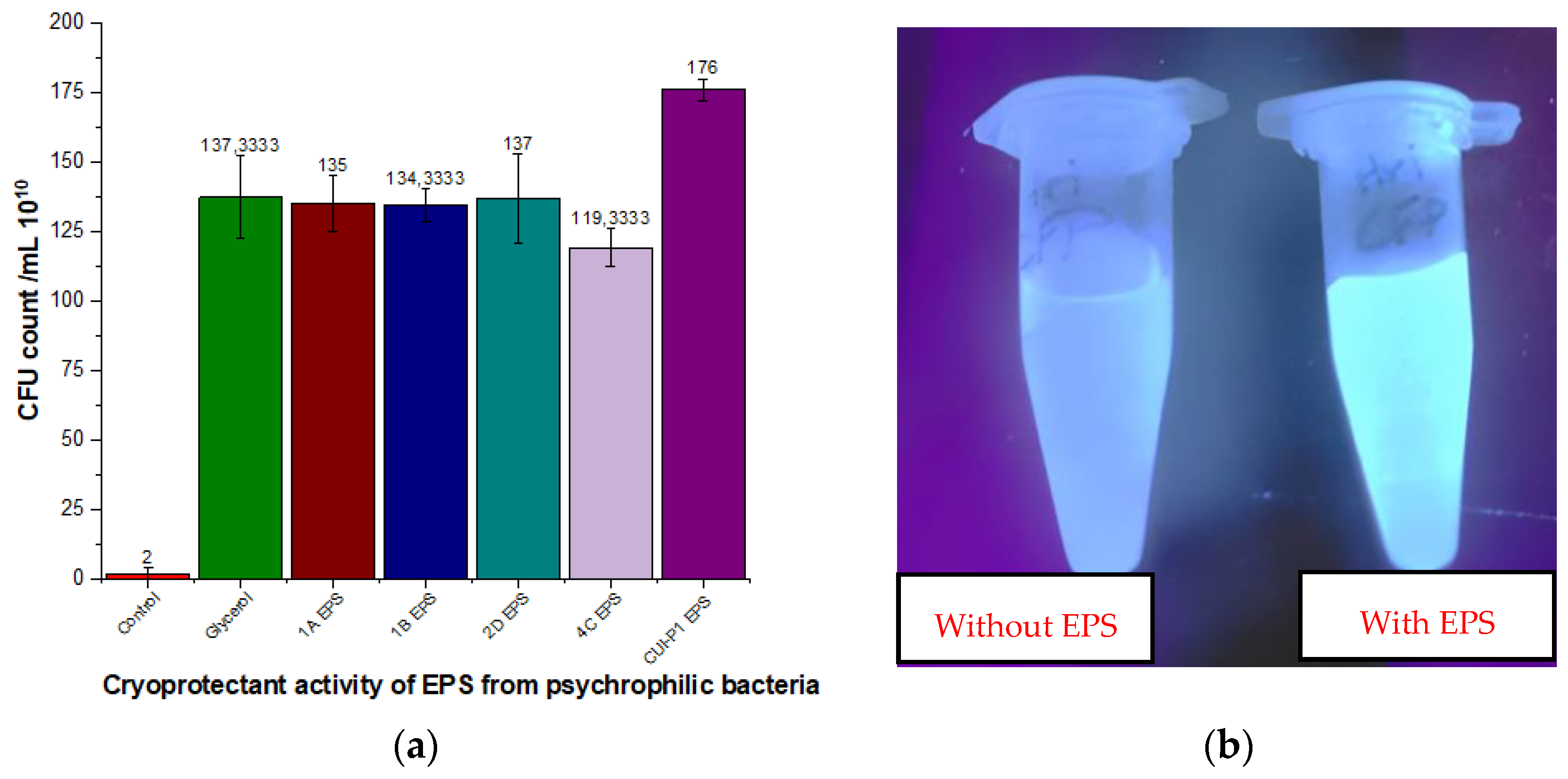
3.3. Cryoprotectant Role of Isolated EPS
3.4. Emulsification Stability
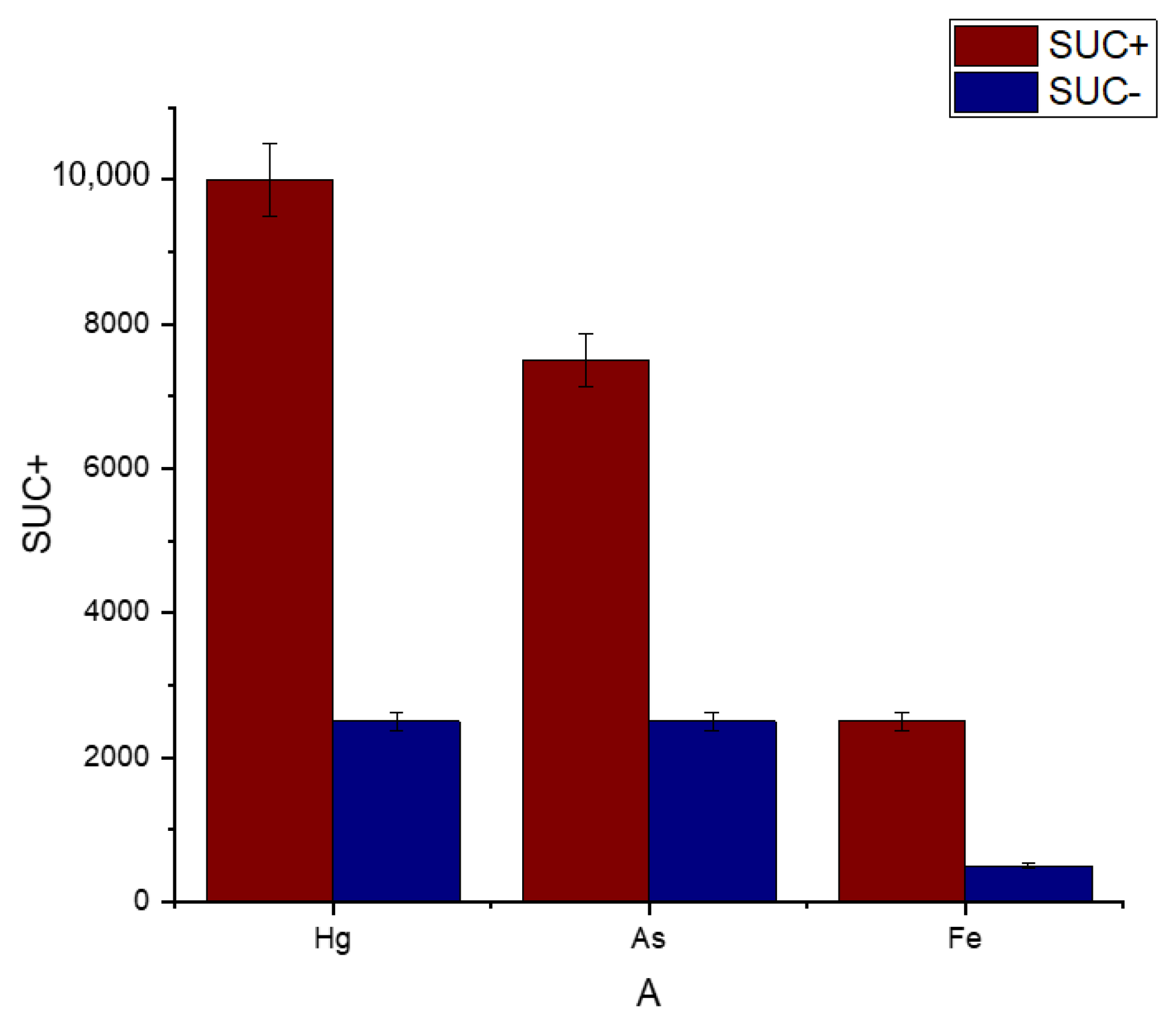
3.5. Metal Tolerance
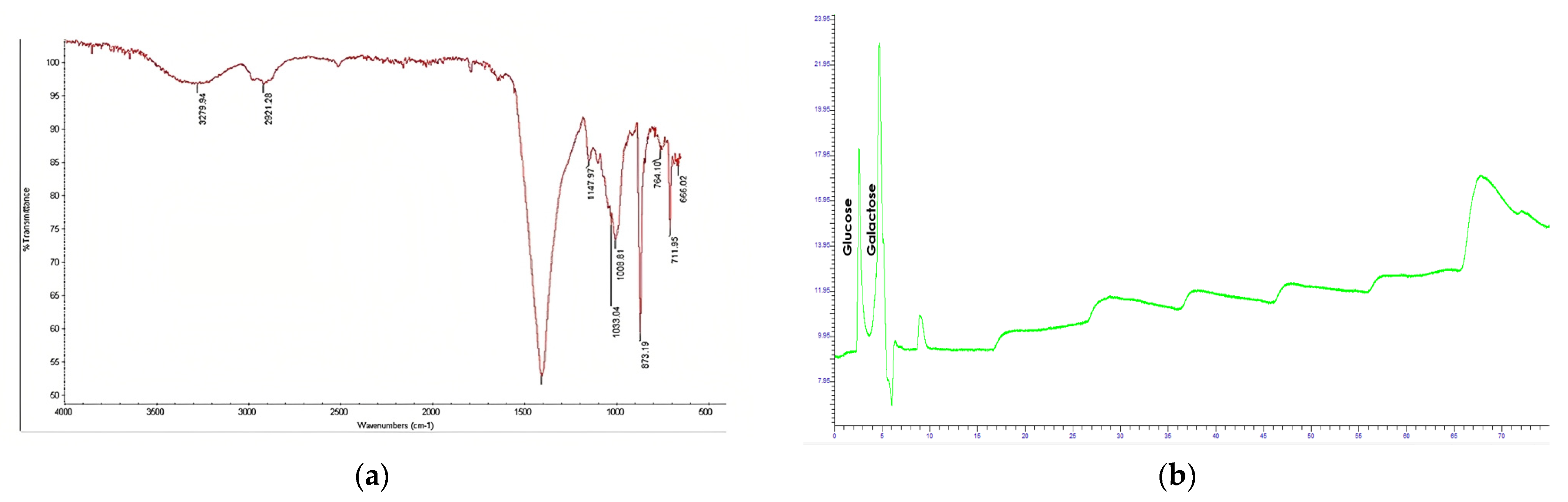
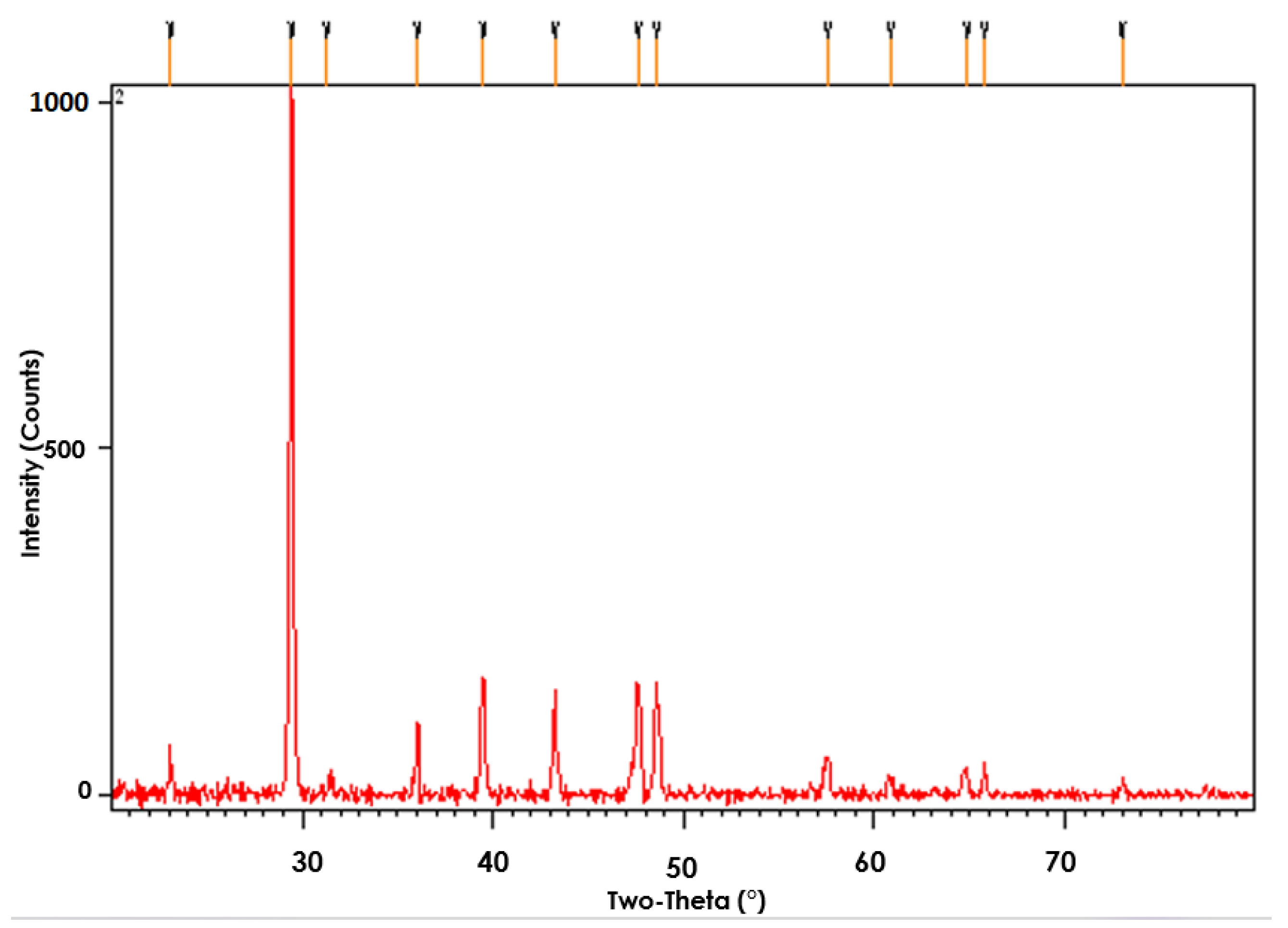
3.6. Chemical Characterization of EPS
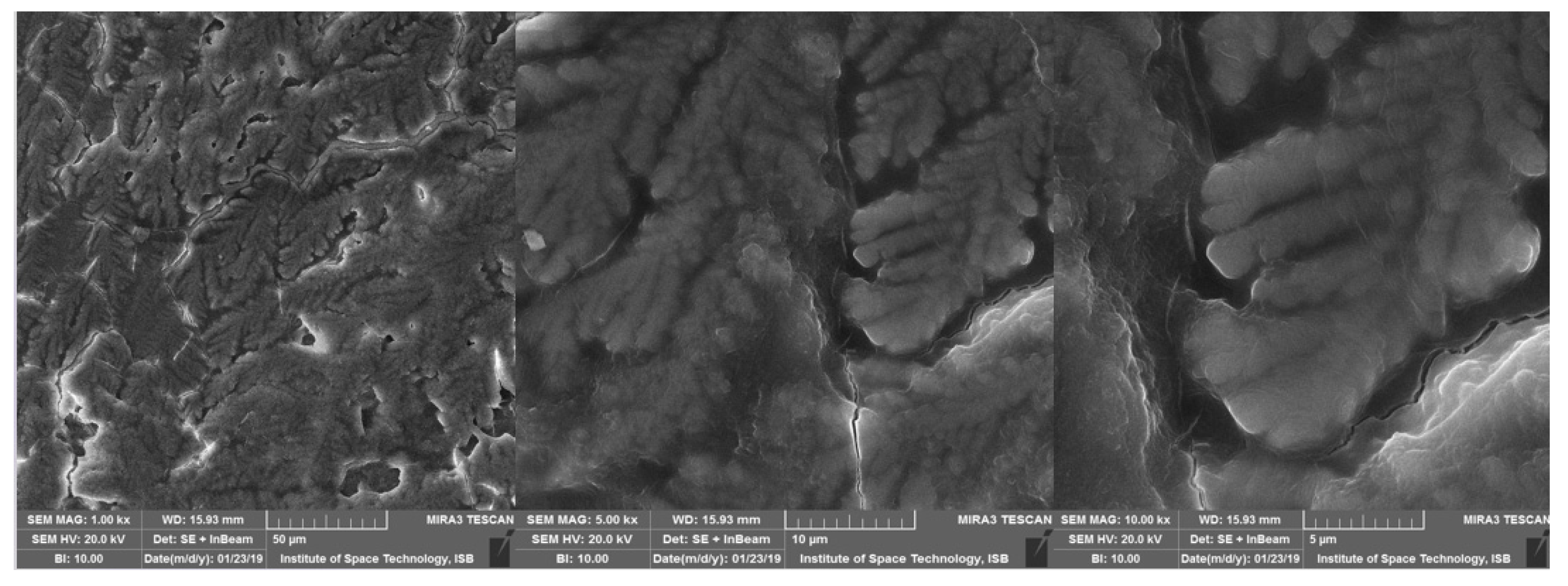
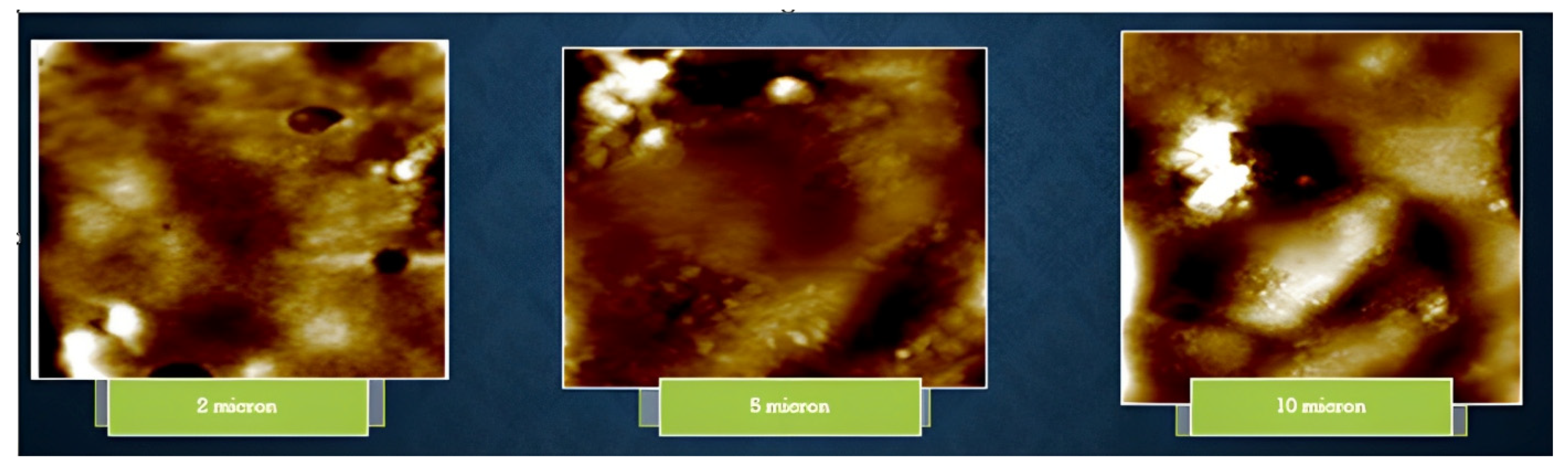
3.7. Physical Characterization of EPS
3.8. Genome Sequencing of Acinetobacter sp. CUI-P1 Provides Genetic Evidence for the EPS Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antunes, L.C.S.; Imperi, F.; Carattoli, A.; Visca, P. Deciphering the Multifactorial Nature of Acinetobacter baumannii Pathogenicity. PLoS ONE 2011, 6, e22674. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Saxena, A.K. Prospecting cold deserts of north western Himalayas for microbial diversity and plant growth promoting attributes. J. Biosci. Bioeng. 2015, 119, 683–693. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, B.-G.; Park, H.J.; Yim, J.H. Cryoprotective properties and preliminary characterization of exopolysaccharide (P-Arcpo 15) produced by the Arctic bacterium Pseudoalteromonas elyakovii Arcpo 15. Prep. Biochem. Biotechnol. 2016, 46, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Carrión, O.; Delgado, L.; Mercade, E. New emulsifying and cryoprotective exopolysaccharide from Antarctic Pseudomonas sp. ID1. Carbohydr. Polym. 2015, 117, 1028–1034. [Google Scholar] [CrossRef]
- Mojib, N.; Farhoomand, A.; Andersen, D.T.; Bej, A.K. UV and cold tolerance of a pigment-producing Antarctic Janthinobacterium sp. Ant5-2. Extremophiles 2013, 17, 367–378. [Google Scholar] [CrossRef]
- Wang, J.; Salem, D.R.; Sani, R.K. Extremophilic exopolysaccharides: A review and new perspectives on engineering strategies and applications. Carbohydr. Polym. 2018, 205, 8–26. [Google Scholar] [CrossRef] [PubMed]
- Schlüter, O.; Fröhling, A. Encyclopedia of Food Microbiology; Elsevier: Oxford, UK, 2014. [Google Scholar]
- Rahal, J.J. Novel Antibiotic Combinations against Infections with Almost Completely Resistant Pseudomonas aeruginosa and Acinetobacter Species. Clin. Infect. Dis. 2006, 43 (Suppl. S2), S95–S99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangiagalli, M.; Bar-Dolev, M.; Tedesco, P.; Natalello, A.; Kaleda, A.; Brocca, S.; Miceli, C.; Braslavsky, I.; Lotti, M. Cryo-protective effect of an ice-binding protein derived from Antarctic bacteria. FEBS J. 2017, 284, 163–177. [Google Scholar] [CrossRef] [Green Version]
- Sartorio, M.G.; Cortez, N.; González, J.M. Structure and functional properties of the cold-Adapted catalase from Acinetobacter sp. Ver3 native to the Atacama plateau in northern Argentina. Acta Crystallogr. Sect. D Struct. Biol. 2021, 77, 369–379. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Xie, J. Response to Cold Adaption in Acinetobacter johnsonii XY27 from Spoiled Bigeye Tuna (Thunnus obesus): Membrane Protein Composition and Protein Biomarker Identification by Proteomics. J. Agric. Food Chem. 2022, 70, 10000–10010. [Google Scholar] [CrossRef]
- Casillo, A.; Parrilli, E.; Sannino, F.; Mitchell, D.E.; Gibson, M.I.; Marino, G.; Lanzetta, R.; Parrilli, M.; Cosconati, S.; Novellino, E.; et al. Structure-activity relationship of the exopolysaccharide from a psychrophilic bacterium: A strategy for cryoprotection. Carbohydr. Polym. 2017, 156, 364–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rashid, F.; Ahmed, Z.; Hussain, S.; Huang, J.-Y.; Ahmad, A. Linum usitatissimum L. seeds: Flax gum extraction, physicochemical and functional characterization. Carbohydr. Polym. 2019, 215, 29–38. [Google Scholar] [CrossRef]
- Pessôa, M.G.; Vespermann, K.A.; Paulino, B.N.; Barcelos, M.C.; Pastore, G.M.; Molina, G. Newly isolated microorganisms with potential application in biotechnology. Biotechnol. Adv. 2019, 37, 319–339. [Google Scholar] [CrossRef] [PubMed]
- Afreen, A.; Ahmed, Z.; Khalid, N.; Ferheen, I.; Ahmed, I. Optimization and cholesterol-lowering activity of exopolysaccharide from Lactiplantibacillus paraplantarum NCCP 962. Appl. Microbiol. Biotechnol. 2023, 107, 1189–1204. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, J.; Yang, Z. Immunomodulatory and antitumor activities of the exopolysaccharide produced by potential probiotic Lactobacillus plantarum YW11 in a HT-29 tumor-burdened nude mouse model. Food Sci. Technol. 2022, 42. [Google Scholar] [CrossRef]
- Poli, A.; Di Donato, P.; Tommonaro, G.; Abbamondi, G.R.; Finore, I.; Nicolaus, B. Exopolysaccharide-Producing Microorganisms from Extreme Areas: Chemistry and Application. In Extremophiles in Eurasian Ecosystems: Ecology, Diversity, and Applications; Springer: Berlin/Heidelberg, Germany, 2018; pp. 405–433. [Google Scholar]
- Schmid, J.; Rühmann, B.; Sieber, V.; Romero-Jiménez, L.; Sanjuán, J.; Pérez-Mendoza, D. Screening of c-di-GMP-Regulated Exopolysaccharides in Host Interacting Bacteria. In Host-Pathogen Interactions; Springer: Berlin/Heidelberg, Germany, 2018; pp. 263–275. [Google Scholar]
- Naik, M.M.; Dubey, S. Lead-and mercury-resistant marine bacteria and their application in lead and mercury bioremediation. In Marine Pollution and Microbial Remediation; Springer: Berlin/Heidelberg, Germany, 2017; pp. 29–40. [Google Scholar]
- Ali, P.; Chen, F.; Hassan, F.; Sosa, A.; Khan, S.; Badshah, M.; Shah, A.A. Bacterial community characterization of Batura Glacier in the Karakoram Range of Pakistan. Int. Microbiol. 2021, 24, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ahmed, Z.; Feng, W.; Li, C.; Song, S. Physicochemical properties of exopolysaccharide produced by Lactobacillus kefiranofaciens ZW3 isolated from Tibet kefir. Int. J. Biol. Macromol. 2008, 43, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Idrees, M.; Thangavelu, K.; Sikaroodi, M.; Smith, C.; Sivaraman, J.; Gillevet, P.; Bokhari, H. Novel fluorescent protein from Hydnophora rigida possesses green emission. Biochem. Biophys. Res. Commun. 2014, 448, 33–38. [Google Scholar] [CrossRef]
- Castellane, T.C.L.; Otoboni, A.M.M.B.; Lemos, E.G.D.M. Characterization of Exopolysaccharides Produced by Rhizobia Species. Rev. Bras. De Ciência Do Solo 2015, 39, 1566–1575. [Google Scholar] [CrossRef] [Green Version]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, J.D.; Nielsen, H.; von Heijne, G.; Brunak, S. Improved Prediction of Signal Peptides: SignalP 3.0. J. Mol. Biol. 2004, 340, 783–795. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]
- Alikhan, N.-F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Stothard, P.; Wishart, D.S. Circular genome visualization and exploration using CGView. Bioinformatics 2005, 21, 537–539. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Ai, L.; Cui, S.W. Fourier Transform Infrared Spectroscopy (FTIR) for Carbohydrate Analysis In Methodology for Structural Analysis of Polysaccharides; Springer: Berlin/Heidelberg, Germany, 2018; pp. 69–71. [Google Scholar]
- Abid, Y.; Casillo, A.; Gharsallah, H.; Joulak, I.; Lanzetta, R.; Corsaro, M.M.; Attia, H.; Azabou, S. Production and structural characterization of exopolysaccharides from newly isolated probiotic lactic acid bacteria. Int. J. Biol. Macromol. 2018, 108, 719–728. [Google Scholar] [CrossRef]
- Caruso, C.; Rizzo, C.; Mangano, S.; Poli, A.; Di Donato, P.; Nicolaus, B.; Di Marco, G.; Michaud, L.; Lo Giudice, A. Extracellular polymeric substances with metal adsorption capacity produced by Pseudoalteromonas sp. MER144 from Antarctic seawater. Environ. Sci. Pollut. Res. 2018, 25, 4667–4677. [Google Scholar] [CrossRef]
- Shang, N.; Xu, R.; Li, P. Structure characterization of an exopolysaccharide produced by Bifidobacterium animalis RH. Carbohydr. Polym. 2013, 91, 128–134. [Google Scholar] [CrossRef]
- Razmkhah, S.; Razavi, S.M.A.; Mohammadifar, M.A.; Koocheki, A.; Ale, M.T. Stepwise extraction of Lepidium sativum seed gum: Physicochemical characterization and functional properties. Int. J. Biol. Macromol. 2016, 88, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Rani, R.P.; Anandharaj, M.; Sabhapathy, P.; Ravindran, A.D. Physiochemical and biological characterization of novel exopolysaccharide produced by Bacillus tequilensis FR9 isolated from chicken. Int. J. Biol. Macromol. 2017, 96, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Schmid, J.; Heider, D.; Wendel, N.J.; Sperl, N.; Sieber, V. Bacterial Glycosyltransferases: Challenges and Opportunities of a Highly Diverse Enzyme Class Toward Tailoring Natural Products. Front. Microbiol. 2016, 7, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mettu, R.; Chen, C.-Y.; Wu, C.-Y. Synthetic carbohydrate-based vaccines: Challenges and opportunities. J. Biomed. Sci. 2020, 27, 9. [Google Scholar] [PubMed] [Green Version]
- Poli, A.; Anzelmo, G.; Nicolaus, B. Bacterial Exopolysaccharides from Extreme Marine Habitats: Production, Characterization and Biological Activities. Mar. Drugs 2010, 8, 1779–1802. [Google Scholar] [CrossRef]
- Soltaninejad, A.; Jazini, M.; Karimi, K. Biorefinery for efficient xanthan gum, ethanol, and biogas production from potato crop residues. Biomass Bioenergy 2022, 158, 106354. [Google Scholar] [CrossRef]
- Amao, J.A.; Omojasola, P.F.; Barooah, M. Isolation and characterization of some exopolysaccharide producing bacteria from cassava peel heaps. Sci. Afr. 2019, 4, e00093. [Google Scholar]
- Ye, S.; Zhang, J.; Liu, Z.; Zhang, Y.; Li, J.; Li, Y.O. Biosynthesis of selenium rich exopolysaccharide (Se-EPS) by Pseudomonas PT-8 and characterization of its antioxidant activities. Carbohydr. Polym. 2016, 142, 230–239. [Google Scholar]
- Vidhyalakshmi, R.; Nachiyar, C.V.; Kumar, G.N.; Sunkar, S.; Badsha, I. Production, characterization and emulsifying property of exopolysaccharide produced by marine isolate of Pseudomonas fluorescens. Biocatal. Agric. Biotechnol. 2018, 16, 320–325. [Google Scholar] [CrossRef]
- Sandhya, V.; Ali, S.Z. The production of exopolysaccharide by Pseudomonas putida GAP-P45 under various abiotic stress conditions and its role in soil aggregation. Microbiology 2015, 84, 512–519. [Google Scholar]
- Korcz, E.; Varga, L. Exopolysaccharides from lactic acid bacteria: Techno-functional application in the food industry. Trends Food Sci. Technol. 2021, 110, 375–384. [Google Scholar] [CrossRef]
- Zannini, E.; Waters, D.M.; Coffey, A.; Arendt, E.K. Production, properties, and industrial food application of lactic acid bacteria-derived exopolysaccharides. Appl. Microbiol. Biotechnol. 2015, 100, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Chrismas, N.A.M.; Barker, G.; Anesio, A.M.; Sánchez-Baracaldo, P. Genomic mechanisms for cold tolerance and production of exopolysaccharides in the Arctic cyanobacterium Phormidesmis priestleyi BC1401. BMC Genom. 2016, 17, 533. [Google Scholar] [CrossRef] [Green Version]
- van Kranenburg, R.; Marugg, J.D.; Van Swam, I.I.; Willem, N.J.; De Vos, W.M. Molecular characterization of the plasmid-encoded eps gene cluster essential for exopolysaccharide biosynthesis in Lactococcus lactis. Mol. Microbiol. 1997, 24, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Boels, I.C.; van Kranenburg, R.; Kanning, M.W.; Chong, B.F.; de Vos, W.M.; Kleerebezem, M. Increased Exopolysaccharide Production in Lactococcus lactis due to Increased Levels of Expression of the NIZO B40 epsGene Cluster. Appl. Environ. Microbiol. 2003, 69, 5029–5031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soumya, M.; Nampoothiri, K.M. An overview of functional genomics and relevance of glycosyltransferases in exopolysaccharide production by lactic acid bacteria. Int. J. Biol. Macromol. 2021, 184, 1014–1025. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, C.; Liu, P.; Ahmed, Z.; Xiao, P.; Bai, X. Physical characterization of exopolysaccharide produced by Lactobacillus plantarum KF5 isolated from Tibet Kefir. Carbohydr. Polym. 2010, 82, 895–903. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Source | Biochemical Identification | EPS Production |
|---|---|---|---|
| CUI-P1 | Pissan Glacier | Acinetobacter sp. | Yes |
| CUI-1A | Passu Glacier | Exiguobacterium indicum | Yes |
| CUI-1B | Passu Glacier | Hymenobacter sp. | Yes |
| CUI-1C | Passu Glacier | Taxeobacter sp. | No |
| CUI-1D | Passu Glacier | Serratia sp. | No |
| CUI-2A | Passu Glacier | Enterobacter sp. | No |
| CUI-2B | Passu Glacier | Aeromonas hydrophila | No |
| CUI-2C | Passu Glacier | Enterococcus spp. | No |
| CUI-2D | Passu Glacier | Staphylococcus lantus | Yes |
| CUI-2E | Passu Glacier | Acinetobacter spp. | No |
| CUI-2F | Passu Glacier | Lactobacillus spp. | No |
| CUI-2G | Passu Glacier | Aeromonas hydrophila | No |
| CUI-3A | Passu Glacier | Vibrio cholerae | No |
| CUI-4A | Passu Glacier | Sphingomonas sp. | Yes |
| CUI-4B | Passu Glacier | Pseudomonas gessardii | No |
| CUI-4C | Passu Glacier | Bacillus subtilis | No |
| CUI-4D | Passu Glacier | Acinetobacter baumannii | No |
| CUI-5A | Pissan Glacier | Flavobacterium spp. | No |
| CUI-5B | Pissan Glacier | Pseudomonas sp. | No |
| CUI-5C | Pissan Glacier | Pseudomonas sp. | No |
| CUI-7A | Pissan Glacier | Aeromonas hydrophila | No |
| CUI-7B | Pissan Glacier | Serratia sp. | No |
| CUI-8C | Pissan Glacier | Pseudomonas aeruginosa | No |
| CUI-9A | Pissan Glacier | Serratia marcescens | No |
| CUI-9C | Pissan Glacier | Aeromonas sp. | No |
| Emulsifier | Incubation Time (min) | Optical Density A540nm | Emulsifying Activity % |
|---|---|---|---|
| Standard | 0 | 0.17 | 100 |
| 30 | 0.069 | 40 | |
| 60 | 0.032 | 18 | |
| 90 | 0.019 | 11 | |
| EPS | 0 | 0.17 | 100 |
| 30 | 0.166 | 97.6 | |
| 60 | 0.162 | 95 | |
| 90 | 0.141 | 87 |
| Name of Strain | Acinetobacter Strain CUI-P1 | Acinetobacter baumannii ATCC 17978 |
|---|---|---|
| Genomic Size (Mb) | 19.5 | 4.09 |
| GC Content (%) | 42.9 | 38.9 |
| Number of Genes | 9718 | 3575 |
| Number of RNAs | 166 | 11 |
| Contigs | 199 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferheen, I.; Ahmed, Z.; Alonazi, W.B.; Pessina, A.; Ibrahim, M.; Pucciarelli, S.; Bokhari, H. Diverse Repertoire and Relationship of Exopolysaccharide Genes in Cold-Adapted Acinetobacter sp. CUI-P1 Revealed by Comparative Genome Analysis. Microorganisms 2023, 11, 885. https://doi.org/10.3390/microorganisms11040885
Ferheen I, Ahmed Z, Alonazi WB, Pessina A, Ibrahim M, Pucciarelli S, Bokhari H. Diverse Repertoire and Relationship of Exopolysaccharide Genes in Cold-Adapted Acinetobacter sp. CUI-P1 Revealed by Comparative Genome Analysis. Microorganisms. 2023; 11(4):885. https://doi.org/10.3390/microorganisms11040885
Chicago/Turabian StyleFerheen, Ifra, Zaheer Ahmed, Wadi B. Alonazi, Alex Pessina, Muhammad Ibrahim, Sandra Pucciarelli, and Habib Bokhari. 2023. "Diverse Repertoire and Relationship of Exopolysaccharide Genes in Cold-Adapted Acinetobacter sp. CUI-P1 Revealed by Comparative Genome Analysis" Microorganisms 11, no. 4: 885. https://doi.org/10.3390/microorganisms11040885
APA StyleFerheen, I., Ahmed, Z., Alonazi, W. B., Pessina, A., Ibrahim, M., Pucciarelli, S., & Bokhari, H. (2023). Diverse Repertoire and Relationship of Exopolysaccharide Genes in Cold-Adapted Acinetobacter sp. CUI-P1 Revealed by Comparative Genome Analysis. Microorganisms, 11(4), 885. https://doi.org/10.3390/microorganisms11040885