

Novel Genetic Lineages of Rickettsia helvetica Associated with Ixodes apronophorus and Ixodes trianguliceps Ticks
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
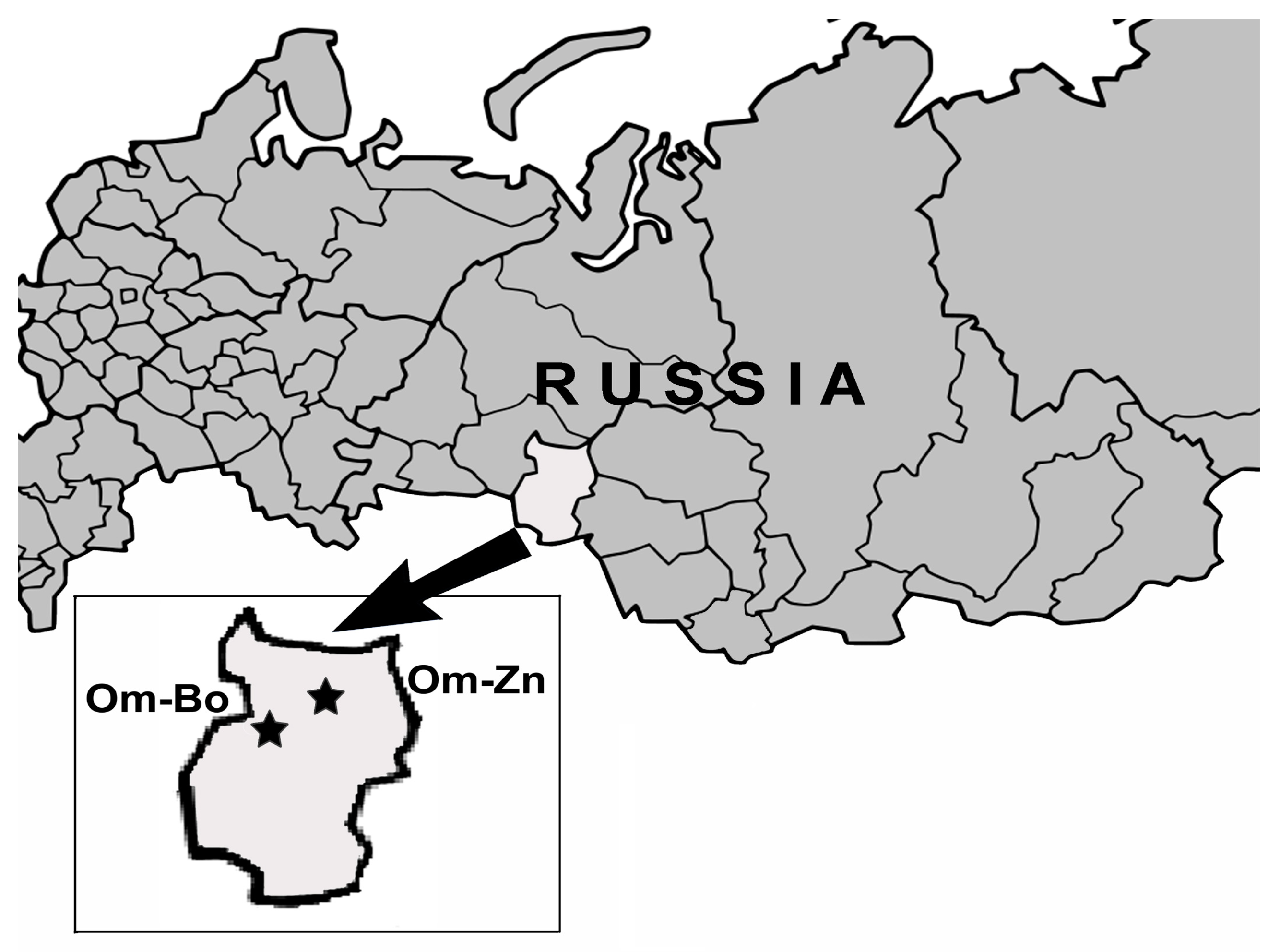
2.1. Sampling
2.2. Tick Metamorphosis
2.3. DNA Extraction
2.4. Detection and Genotyping of Rickettsia spp.
2.5. Sequencing and Phylogenetic Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amplified Locus | Organism | Reaction | Primer Name | Primer Sequences 5′-3′ | T# (°C) | References |
|---|---|---|---|---|---|---|
| 16S rRNA | Rickettsia spp. | Primary | 16S1 | gacgggtgagtaacacgtggg | 56 | [22] |
| gene | 16S2 | gtcttttagggatttgctccac | ||||
| Nested | 16S3 | gatggatgagcccgcgtcag | 60 | |||
| 16S4 | gcatctctgcgatccgcgac | |||||
| gltA gene | Rickettsia spp. | Primary | glt1 | gattgctttacttacgaccc | 52 | [22] |
| glt2 | tgcatttctttccattgtgc | |||||
| Nested | glt3 | tatagacggtgataaaggaatc | 53 | |||
| glt4 | cagaactaccgatttctttaagc | |||||
| ompA gene | Rickettsia spp. | Conventional | Rr190.70p | atggcgaatatttctccaaaa | 55 | [23] |
| fragment I | 190-701 | gttccgttaatggcagcatct | ||||
| ompA gene | Rickettsia spp. | Primary | Afnw_1 | ggcacaaatactttaacattacc | 52 | [24] |
| 190-6808 | cacgaactttcacactacc | [23] | ||||
| Nested | Afnw_3 | aagcctactcctaaagagaatg | 53 | [24] | ||
| Afnw_4 | cgacagtctctagtgccg | |||||
| Rickettsia spp. | Primary | A1 | taacattacaagctggaggaagcc | 58 | [22] | |
| A2 | ttcagagcctgaccaccgg | |||||
| Nested | A5 | caagtgctggtgatgttacta | 56 | |||
| A6 | tagttacatttcctgcacctac | |||||
| R. helvetica | Primary | Afn1_helv | gtaatactagcatcaccgaaatcc | 55 | This study | |
| 190-6808 | cacgaactttcacactacc | [23] | ||||
| Nested | 190-5125 | gcggttactttagccaaagg | 54 | |||
| Afnw_4 | cgacagtctctagtgccg | [24] | ||||
| ompA gene | “Ca. R. tarasevichiae” | Conventional | 190-5125 | gcggttactttagccaaagg | 56 | [23] |
| fragment IV | 190-6013m * | gcatcttytgcgttgyattac | ||||
| ompB gene | Rickettsia spp. | Primary | M59 F | ccgcagggttggtaactgc | 55 | [25] |
| 120-1497m * | cctatatcgccggtaattgtagc | |||||
| Nested | BR1 | gttactaatggatttattcaagt | 53 | [22] | ||
| BR2 | gcataaacttgtccagcgat | |||||
| Rickettsia spp. | Primary | B2f_5 | taaacttgctgacggtacag | 56 | [24] | |
| B2f_2 | cgattatgccgttatcgcttccaag | |||||
| Nested | B2f_3 | gtagcctaacaaatgctcaaac | 52 | |||
| 120-2399 | cttgtttgtttaatgttacggt | [25] | ||||
| R. helvetica | Primary | B2f_3 | gtagcctaacaaatgctcaaac | 52 | ||
| B2f_2 | cgattatgccgttatcgcttccaag | |||||
| Nested | B2f_1helv | cagtacaattcgctcacaacac | 55 | This study | ||
| 120-2399 | cttgtttgtttaatgttacggt | |||||
| Rickettsia spp. | Primary | B1 | atatgcaggtatcggtact | 56 | [22] | |
| B2 | ccatataccgtaagctacat | |||||
| Nested | B3 | gcaggtatcggtactataaac | 56 | |||
| B4 | aatttacgaaacgattacttccgg | |||||
| Rickettsia spp. | Primary | 120-3462 | ccacaggaactacaaccatt | 52 | [25] | |
| 120-4879m * | tagaagtttacacggacttttagag | |||||
| Nested | B3f_3f | gctggacctgaagctggagc | 55 | [24] | ||
| 120-4879m * | tagaagtttacacggacttttagag | [25] | ||||
| sca4 gene | Rickettsia spp. | Primary | D1f | atgagtaaagacggtaacct | 52 | [26] |
| D1876rm * | tagtttgttccgccgtaatc | |||||
| Nested | sc1f_3 | gatgtaggtgatgaactctg | 52 | [24] | ||
| D1390r | cttgcttttcagcaatatcac | [26] | ||||
| Primary | sc4-1 | atgtctctgaattaagcaatgc | 52 | [22] | ||
| Rj2837r | cctgatactacccttacatc | [27] | ||||
| Nested | sc4-5 | ccggcacaacaacaattgatg | 50 | [22] | ||
| sc4-6 | cctttaccagctcatctactt | |||||
| Primary | sc4-3 | aattattaggctctgtattaaaga | 52 | [22] | ||
| D3069r | tcagcgttgtggaggggaag | [26] | ||||
| Nested | sc4-5 | ccggcacaacaacaattgatg | 52 | [22] | ||
| sc4-7 | ctctcttttaataggtgttgatt | [24] | ||||
| htrA gene | Rickettsia spp. | Conventional | 17k-5 | gctttacaaaattctaaaaaccatata | 55 | [28] |
| 17k-3 | tgtctatcaattcacaacttgcc | |||||
| 23S-5S IGS | Rickettsia spp. | Conventional | RCK/23-5-F | gataggtcrgrtgtggaagca | 55 | [29] |
| RCK/23-5-R | tcgggaygggatcgtgtgtttc | |||||
| groESL | Rickettsia spp. | Primary | Ric-ESL-F1 | ggtaaatgggcaggyaccgaa | 60 | [30] |
| operon | Ric-ESL-R1 | gaagcaacrgaagcagcatctt | ||||
| Nested | Ric-ESL-F2 | atcgttatgaaagaaagcgayg | 58 | |||
| Ric-ESL-R2 | agwgcagtacgcactactttagc |
2.6. Statistical Analysis
2.7. Nucleotide Sequence Accession Numbers
3. Results
3.1. Sampling
3.2. Detection of Rickettsia spp. in Non-Molted Ticks
3.3. Detection of Rickettsia spp. in Molted Ixodes spp. Ticks
3.4. Genotyping of Rickettsia raoultii and Candidate Species
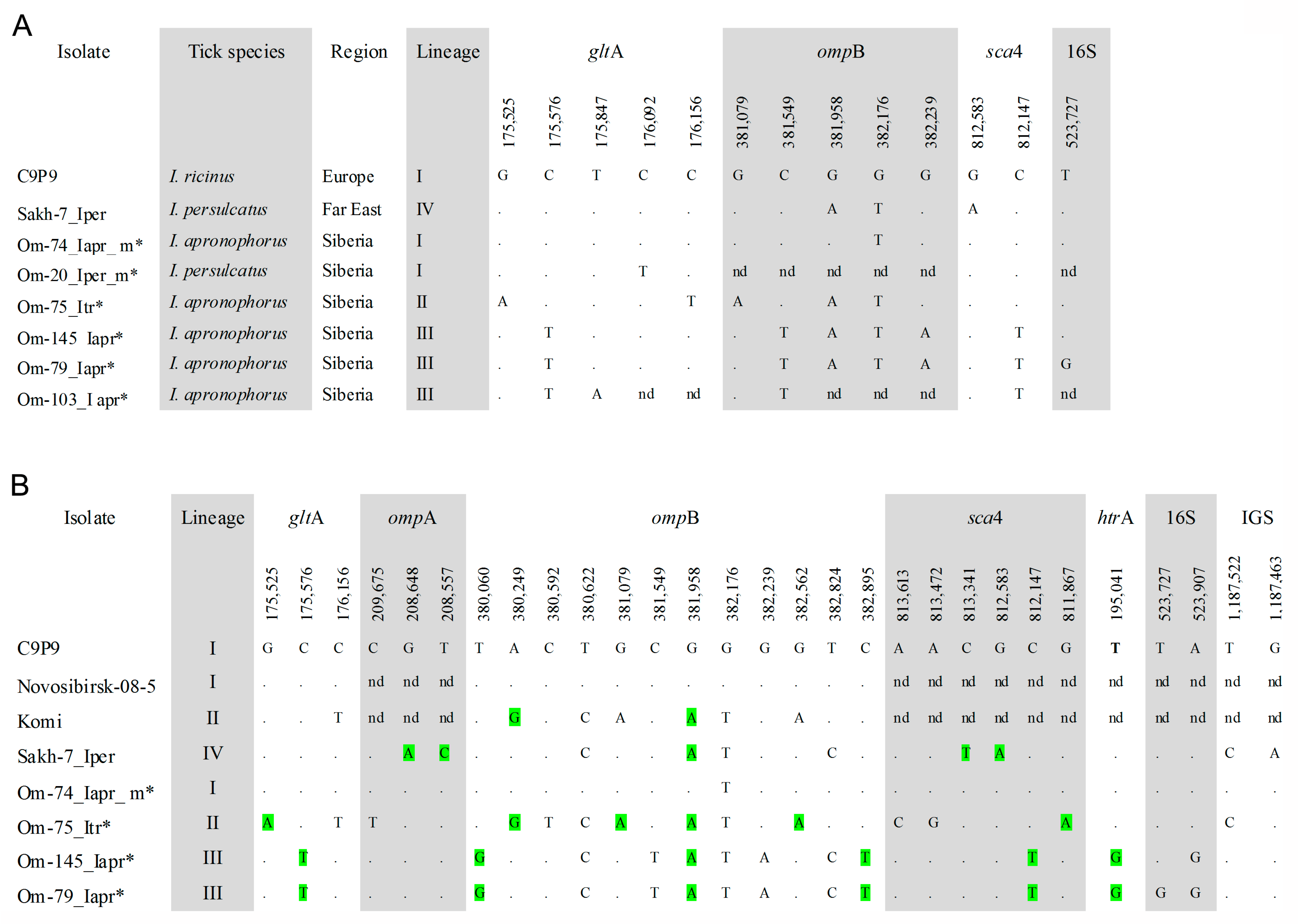
3.5. Genotyping of R. helvetica
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eisen, L. Vector competence studies with hard ticks and Borrelia burgdorferi sensu lato spirochetes: A review. Ticks Tick Borne Dis. 2020, 11, 101359. [Google Scholar] [CrossRef] [PubMed]
- Hoornstra, D.; Azagi, T.; van Eck, J.A.; Wagemakers, A.; Koetsveld, J.; Spijker, R.; Platonov, A.E.; Sprong, H.; Hovius, J.W. Prevalence and clinical manifestation of Borrelia miyamotoi in Ixodes ticks and humans in the northern hemisphere: A systematic review and meta-analysis. Lancet. Microbe 2022, 3, e772–e786. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [PubMed]
- Rar, V.; Golovljova, I. Anaplasma, Ehrlichia, and “Candidatus Neoehrlichia” bacteria: Pathogenicity, biodiversity, and molecular genetic characteristics, a review. Infect. Genet. Evol. 2011, 11, 1842–1861. [Google Scholar] [CrossRef] [PubMed]
- Balashov, Y.S. Ixodid Ticks—Parasites and Vectors of Diseases; Nauka: Sankt-Peterburg, Russia, 1998. (In Russian) [Google Scholar]
- Yakimenko, V.V.; Malkova, M.G.; Shpynov, S.N. Ixodid Ticks of the Western Siberia; Omskiy Nauchnyi Vestnik: Omsk, Russia, 2013; 240p. (In Russian) [Google Scholar]
- Karimov, A.V.; KoralloVinarskaya, N.P.; Kuzmenko, Y.F.; Vinarski, M.V. Ixodes apronophorus Schulze (Acari: Ixodida: Ixodidae): Distribution, abundance, and diversity of its mammal hosts in West Siberia (Results of a 54-year long surveillance). Diversity 2022, 14, 702. [Google Scholar] [CrossRef]
- Rar, V.; Yakimenko, V.; Tikunov, A.; Vinarskaya, N.; Tancev, A.; Babkin, I.; Epikhina, T.; Tikunova, N. Genetic and morphological characterization of Ixodes apronophorus from Western Siberia, Russia. Ticks Tick Borne Dis. 2020, 11, 101284. [Google Scholar] [CrossRef]
- Filippova, N.A. Ixodid ticks of the subfamily Ixodinae. In The Fauna of the USSR. Arachnida; Nauka Publishing House: Leningrad, Russia, 1977; New Ser 4. (In Russian) [Google Scholar]
- Korenberg, E.I.; Kovalevskii, Y.V.; Gorelova, N.B.; Nefedova, V.V. Comparative analysis of the roles of Ixodes persulcatus and I. trianguliceps ticks in natural foci of ixodid tick-borne borrelioses in the Middle Urals, Russia. Ticks Tick Borne Dis. 2015, 6, 316–321. [Google Scholar] [CrossRef]
- Rar, V.; Livanova, N.; Tkachev, S.; Kaverina, G.; Tikunov, A.; Sabitova, Y.; Igolkina, Y.; Panov, V.; Livanov, S.; Fomenko, N.; et al. Detection and genetic characterization of a wide range of infectious agents in Ixodes pavlovskyi ticks in Western Siberia, Russia. Parasit. Vectors 2017, 10, 258. [Google Scholar] [CrossRef]
- Rar, V.; Yakimenko, V.; Tikunov, A.; Makenov, M.; Epikhina, T.; Tancev, A.; Tikunova, N. Genetic variability of Anaplasmataceae circulating in small mammals and ticks in an Ixodes persulcatus/Ixodes trianguliceps sympatric area in Russian Siberia. Ticks Tick Borne Dis. 2020, 11, 101499. [Google Scholar] [CrossRef]
- Sabitova, Y.; Rar, V.; Tikunov, A.; Yakimenko, V.; Korallo-Vinarskaya, N.; Livanova, N.; Tikunova, N. Detection and genetic characterization of a putative novel Borrelia genospecies in Ixodes apronophorus/Ixodes persulcatus/Ixodes trianguliceps sympatric areas in Western Siberia. Ticks Tick Borne Dis. 2023, 14, 102075. [Google Scholar] [CrossRef]
- Igolkina, Y.P.; Rar, V.A.; Yakimenko, V.V.; Malkova, M.G.; Tancev, A.K.; Tikunov, A.Y.; Epikhina, T.I.; Tikunova, N.V. Genetic variability of Rickettsia spp. in Ixodes persulcatus/Ixodes trianguliceps sympatric areas from Western Siberia, Russia: Identification of a new Candidatus Rickettsia species. Infect. Genet. Evol. 2015, 34, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Igolkina, Y.; Bondarenko, E.; Rar, V.; Epikhina, T.; Vysochina, N.; Pukhovskaya, N.; Tikunov, A.; Ivanov, L.; Golovljova, I.; Ivanov, M.; et al. Genetic variability of Rickettsia spp. in Ixodes persulcatus ticks from continental and island areas of the Russian Far East. Ticks Tick Borne Dis. 2016, 7, 1284–1289. [Google Scholar] [CrossRef]
- Shpynov, S.; Fournier, P.E.; Rudakov, N.; Tarasevich, I.; Raoult, D. Detection of members of the genera Rickettsia, Anaplasma, and Ehrlichia in ticks collected in the Asiatic part of Russia. Ann. N. Y. Acad. Sci. 2006, 1078, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Vikentjeva, M.; Geller, J.; Remm, J.; Golovljova, I. Rickettsia spp. in rodent-attached ticks in Estonia and first evidence of spotted fever group Rickettsia species Candidatus Rickettsia uralica in Europe. Parasit. Vectors 2021, 14, 65. [Google Scholar] [CrossRef]
- Nowak-Chmura, M.; Siuda, K. Ticks of Poland. Review of contemporary issues and latest research. Ann. Parasitol. 2012, 58, 125–155. [Google Scholar] [PubMed]
- Stanko, M.; Derdáková, M.; Špitalská, E.; Kazimírová, M. Ticks and their epidemiological role in Slovakia: From the past till present. Biologia 2022, 77, 1575–1610. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Pfäffle, M.P.; Petney, T.N. Genus Ixodes Latreille, 1795. In Ticks of Europe and North Africa. A Guide to Species Identification; Estrada-Pena, E., Mihalca, A.D., Petney, T.N., Eds.; Springer: Cham, Switzerland, 2017; pp. 79–91. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Igolkina, Y.; Rar, V.; Vysochina, N.; Ivanov, L.; Tikunov, A.; Pukhovskaya, N.; Epikhina, T.; Golovljova, I.; Tikunova, N. Genetic variability of Rickettsia spp. in Dermacentor and Haemaphysalis ticks from the Russian Far East. Ticks Tick Borne Dis. 2018, 9, 1594–1603. [Google Scholar] [CrossRef]
- Fournier, P.E.; Roux, V.; Raoult, D. Phylogenetic analysis of spotted fever group rickettsiae by study of the outer surface protein rOmpA. Int. J. Syst. Bacteriol. 1998, 48, 839–849. [Google Scholar] [CrossRef]
- Igolkina, Y.; Rar, V.; Yakimenko, V.; Tikunov, A.; Tikunova, N. “Candidatus Rickettsia uralica” and “Candidatus Rickettsia thierseensis” are genetic variants of one species. Ticks Tick Borne Dis. 2022, 13, 101933. [Google Scholar] [CrossRef]
- Roux, V.; Raoult, D. Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol. 2000, 50, 1449–1455. [Google Scholar] [CrossRef] [PubMed]
- Sekeyova, Z.; Roux, V.; Raoult, D. Phylogeny of Rickettsia spp. inferred by comparing sequences of “gene D”, which encodes an intracytoplasmic protein. Int. J. Syst. Evol. Microbiol. 2001, 51, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Inokuma, H. Identification of spotted fever group Rickettsia species by polymerase chain reaction-restriction fragment length polymorphism analysis of the sca4 gene. Vector Borne Zoonotic Dis. 2009, 9, 747–749. [Google Scholar] [CrossRef]
- Labruna, M.B.; McBride, J.W.; Bouyer, D.H.; Camargo, L.M.A.; Camargo, E.P.; Walker, D.H. Molecular evidence for a spotted fever group Rickettsia species in the tick Amblyomma longirostre in Brazil. J. Med. Entomol. 2004, 41, 533–537. [Google Scholar] [CrossRef]
- Jado, I.; Escudero, R.; Gil, H.; Jimenez-Alonso, M.I.; Sousa, R.; Garcia-Perez, A.L.; Rodriguez-Vargas, M.; Lobo, B.; Anda, P. Molecular method for identification of Rickettsia species in clinical and environmental samples. J. Clin. Microbiol. 2006, 44, 4572–4576. [Google Scholar] [CrossRef]
- Shao, J.W.; Zhang, X.L.; Li, W.J.; Huang, H.L.; Yan, J. Distribution and molecular characterization of rickettsiae in ticks in Harbin area of Northeastern China. PLoS Negl. Trop. Dis. 2020, 14, e0008342. [Google Scholar] [CrossRef] [PubMed]
- Kartashov, M.Y.; Glushkova, L.I.; Mikryukova, T.P.; Korabelnikov, I.V.; Egorova, Y.I.; Tupota, N.L.; Protopopova, E.V.; Konovalova, S.N.; Ternovoi, V.A.; Loktev, V.B. Detection of Rickettsia helvetica and Candidatus R. tarasevichiae DNA in Ixodes persulcatus ticks collected in Northeastern European Russia (Komi Republic). Ticks Tick Borne Dis. 2017, 8, 588–592. [Google Scholar] [CrossRef]
- Katargina, O.; Geller, J.; Ivanova, A.; Värv, K.; Tefanova, V.; Vene, S.; Lundkvist, Å.; Golovljova, I. Detection and identification of Rickettsia species in Ixodes tick populations from Estonia. Ticks Tick Borne Dis. 2015, 6, 689–694. [Google Scholar] [CrossRef]
- Balážová, A.; Földvári, G.; Bilbija, B.; Nosková, E.; Široký, P. High prevalence and low diversity of Rickettsia in Dermacentor reticulatus ticks, Central Europe. Emerg. Infect. Dis. 2022, 28, 893–895. [Google Scholar] [CrossRef]
- Cheng, C.; Fu, W.; Ju, W.; Yang, L.; Xu, N.; Wang, Y.M.; Li, H.; Wang, Y.L.; Hu, M.X.; Wen, J.; et al. Diversity of spotted fever group Rickettsia infection in hard ticks from Suifenhe, Chinese-Russian border. Ticks Tick Borne Dis. 2016, 7, 715–719. [Google Scholar] [CrossRef]
- Silaghi, C.; Gilles, J.; Höhle, M.; Pradel, I.; Just, F.T.; Fingerle, V.; Küchenhoff, H.; Pfister, K. Prevalence of spotted fever group rickettsiae in Ixodes ricinus (Acari: Ixodidae) in southern Germany. J. Med. Entomol. 2008, 45, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Schötta, A.M.; Wijnveld, M.; Höss, D.; Stanek, G.; Stockinger, H.; Markowicz, M. Identification and characterization of “Candidatus Rickettsia thierseensis”, a novel spotted fever group Rickettsia species detected in Austria. Microorganisms 2020, 28, 1670. [Google Scholar] [CrossRef] [PubMed]
- Jia, N.; Zheng, Y.C.; Jiang, J.F.; Ma, L.; Cao, W.C. Human infection with Candidatus Rickettsia tarasevichiae. N. Engl. J. Med. 2013, 369, 1178–1180. [Google Scholar] [CrossRef]
- Liu, W.; Li, H.; Lu, Q.B.; Cui, N.; Yang, Z.D.; Hu, J.G.; Fan, Y.D.; Guo, C.T.; Li, X.K.; Wang, Y.W.; et al. Candidatus Rickettsia tarasevichiae infection in Eastern Central China: A case series. Ann. Intern. Med. 2016, 164, 641–648. [Google Scholar] [CrossRef]
- Rudakov, N.; Samoylenko, I.; Shtrek, S.; Igolkina, Y.; Rar, V.; Zhirakovskaia, E.; Tkachev, S.; Kostrykina, T.; Blokhina, I.; Lentz, P.; et al. A fatal case of tick-borne rickettsiosis caused by mixed Rickettsia sibirica subsp. sibirica and “Candidatus Rickettsia tarasevichiae” infection in Russia. Ticks Tick Borne Dis. 2019, 10, 101278. [Google Scholar] [CrossRef] [PubMed]
- Igolkina, Y.; Rar, V.; Krasnova, E.; Filimonova, E.; Tikunov, A.; Epikhina, T.; Tikunova, N. Occurrence and clinical manifestations of tick-borne rickettsioses in Western Siberia: First Russian cases of Rickettsia aeschlimannii and Rickettsia slovaca infections. Ticks Tick Borne Dis. 2022, 13, 101927. [Google Scholar] [CrossRef]
- Shpynov, S.; Fournier, P.E.; Rudakov, N.; Raoult, D. “Candidatus Rickettsia tarasevichiae” in Ixodes persulcatus ticks collected in Russia. Ann. N. Y. Acad Sci. 2003, 990, 162–172. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, Q.; Zhang, X.; Li, Z.; Wang, Z.; Song, M.; Wei, F.; Wang, S.; Liu, Q. Characterization of rickettsiae in ticks in northeastern China. Parasit. Vectors 2016, 13, 498. [Google Scholar] [CrossRef]
- Igolkina, Y.; Nikitin, A.; Verzhutskaya, Y.; Gordeyko, N.; Tikunov, A.; Epikhina, T.; Tikunova, N.; Rar, V. Multilocus genetic analysis indicates taxonomic status of “Candidatus Rickettsia mendelii” as a separate basal group. Ticks Tick Borne Dis. 2023, 14, 102104. [Google Scholar] [CrossRef]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Spitalská, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef]
- Hensley, J.R.; Zambrano, M.L.; Williams-Newkirk, A.J.; Dasch, G.A. Detection of Rickettsia species, and Coxiella-like and Francisella-like endosymbionts in Amblyomma americanum and Amblyomma maculatum from a shared field site in Georgia, United States of America. Vector Borne Zoonotic Dis. 2021, 21, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Moraes-Filho, J.; Costa, F.B.; Gerardi, M.; Soares, H.S.; Labruna, M.B. Rickettsia rickettsii co-feeding transmission among Amblyomma aureolatum ticks. Emerg. Infect. Dis. 2018, 24, 2041–2048. [Google Scholar] [CrossRef] [PubMed]
- Silaghi, C.; Hamel, D.; Thiel, C.; Pfister, K.; Pfeffer, M. Spotted fever group rickettsiae in ticks, Germany. Emerg. Infect. Dis. 2011, 17, 890–892. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, K. Septicaemia with Rickettsia helvetica in a patient with acute febrile illness, rash and myasthenia. J. Infect. 2009, 58, 79–82. [Google Scholar] [CrossRef] [PubMed]





| Site/Tested Ticks | Tick Species | Tick Stage | No. of Ticks | No. (%) of Ticks Containing DNA of | ||||
|---|---|---|---|---|---|---|---|---|
| All Rickettsiae | R.helv | R.tar | R.ural | Mixed Infection | ||||
| Om-Bo/ | I.apr | Larvae | 47 | 34 | 33 | 1 | 0 | 0 |
| non-molted | Nymphs | 5 | 4 | 4 | 0 | 0 | 0 | |
| ticks | Females | 9 | 7 | 6 | 0 | 0 | 1/R.tar + R.helv | |
| Males | 1 | 1 | 1 | 0 | 0 | 0 | ||
| All stages | 62 | 46 (74.2) | 44 (71.0) | 1 (1.6) | 0 | 1 (1.6) | ||
| I.pers | Larvae | 55 | 46 | 0 | 45 | 0 | 1/R.tar + R.raol | |
| Nymphs | 4 | 3 | 0 | 3 | 0 | 0 | ||
| All stages | 59 | 49 (83.1) | 0 | 48 (81.4) | 0 | 1 (1.7) | ||
| I.tr | Larvae | 17 | 10 | 9 | 0 | 1 | 0 | |
| Nymphs | 4 | 4 | 0 | 0 | 4 | 0 | ||
| Females | 3 | 2 | 0 | 0 | 2 | 0 | ||
| All stages | 24 | 16 (66.7) | 9 (37.5) | 0 | 7 (29.2) | 0 | ||
| Om-Bo/ | I.apr | Nymphs | 2 | 1 | 1 | 0 | 0 | 0 |
| molted | Females | 1 | 1 | 1 | 0 | 0 | 0 | |
| ticks | Males | 2 | 1 | 1 | 0 | 0 | 0 | |
| All stages | 5 | 3 (60) | 3 (60) | 0 | 0 | 0 | ||
| I.pers | Nymphs | 3 | 3 | 0 | 2 | 1 | 0 | |
| Females | 7 | 5 | 0 | 5 | 0 | 0 | ||
| Males | 4 | 1 | 0 | 1 | 0 | 0 | ||
| All stages | 14 | 9 (64.3) | 0 | 8 (57.1) | 1 (7.1) | 0 | ||
| I.tr | Nymphs | 1 | 1 | 0 | 0 | 1 | 0 | |
| All stages | 1 | 1 | 0 | 0 | 1 | 0 | ||
| Om-Zn/ | I.apr | Nymphs | 4 | 3 | 3 | 0 | 0 | 0 |
| molted | Males | 1 | 1 | 1 | 0 | 0 | 0 | |
| ticks | All stages | 5 | 4 (80) | 4 (80) | 0 | 0 | 0 | |
| I.pers | Nymphs | 23 | 23 | 0 | 22 | 0 | 1/R.tar + R.sp | |
| Females | 36 | 29 | 0 | 28 | 0 | 1/R.tar + R.helv | ||
| Males | 28 | 22 | 0 | 22 | 0 | 0 | ||
| All stages | 87 | 74 | 0 | 72 (82.8) | 0 | 2 (2.3) | ||
| I.tr | Nymphs | 10 | 0 | 0 | 0 | 0 | 0 | |
| Females | 5 | 2 | 0 | 1 | 1 | 0 | ||
| Males | 8 | 2 | 0 | 1 | 1 | 0 | ||
| All stages | 23 | 4 (17.4) | 0 | 2 (8.7) | 2 (8.7) | 0 | ||
| Total | I.apr | All stages | 72 | 53 (73.6) | 51 (70.8) | 1 (1.4) | 0 | 1 (1.4) |
| I.pers | All stages | 160 | 132 (82.5) | 0 | 128 (80.0) | 1 (0.6) | 3 (1.9) | |
| I.tr | All stages | 48 | 21 (43.8) | 9 (18.8) | 2 (4.2) | 10 (20.8) | 0 | |
| Sites | Tick Species | Tested Ticks | No of Tested Ticks | No of R. helvetica Positive Ticks | No of R. helvetica Samples Belonging to | ||
|---|---|---|---|---|---|---|---|
| Lineage I | Lineage II | Lineage III | |||||
| Om-Bo | I. apronophorus | non-molted | 62 | 45 | 0 | 0 | 45 |
| molted | 5 | 3 | 0 | 1 | 2 | ||
| subtotal | 67 | 48 | 0 | 1 | 47 | ||
| I. persulcatus | non-molted | 59 | 0 | 0 | 0 | 0 | |
| molted | 14 | 0 | 0 | 0 | 0 | ||
| subtotal | 73 | 0 | 0 | 0 | 0 | ||
| I. trianguliceps | non-molted | 24 | 9 | 0 | 9 | 0 | |
| molted | 1 | 0 | 0 | 0 | 0 | ||
| subtotal | 25 | 9 | 0 | 9 | 0 | ||
| Om-Zn | I. apronophorus | molted | 5 | 4 | 3 | 0 | 1 |
| I. persulcatus | molted | 87 | 1 | 1 | 0 | 0 | |
| I. trianguliceps | molted | 23 | 0 | 0 | 0 | 0 | |
| Both | I. apronophorus | total | 72 | 52 | 3 | 1 | 48 |
| sites | I. persulcatus | total | 160 | 1 | 1 | 0 | 0 |
| I. trianguliceps | total | 48 | 9 | 0 | 9 | 0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Igolkina, Y.; Yakimenko, V.; Tikunov, A.; Epikhina, T.; Tancev, A.; Tikunova, N.; Rar, V. Novel Genetic Lineages of Rickettsia helvetica Associated with Ixodes apronophorus and Ixodes trianguliceps Ticks. Microorganisms 2023, 11, 1215. https://doi.org/10.3390/microorganisms11051215
Igolkina Y, Yakimenko V, Tikunov A, Epikhina T, Tancev A, Tikunova N, Rar V. Novel Genetic Lineages of Rickettsia helvetica Associated with Ixodes apronophorus and Ixodes trianguliceps Ticks. Microorganisms. 2023; 11(5):1215. https://doi.org/10.3390/microorganisms11051215
Chicago/Turabian StyleIgolkina, Yana, Valeriy Yakimenko, Artem Tikunov, Tamara Epikhina, Aleksey Tancev, Nina Tikunova, and Vera Rar. 2023. "Novel Genetic Lineages of Rickettsia helvetica Associated with Ixodes apronophorus and Ixodes trianguliceps Ticks" Microorganisms 11, no. 5: 1215. https://doi.org/10.3390/microorganisms11051215
APA StyleIgolkina, Y., Yakimenko, V., Tikunov, A., Epikhina, T., Tancev, A., Tikunova, N., & Rar, V. (2023). Novel Genetic Lineages of Rickettsia helvetica Associated with Ixodes apronophorus and Ixodes trianguliceps Ticks. Microorganisms, 11(5), 1215. https://doi.org/10.3390/microorganisms11051215