
Two Bacillus spp. Strains Improve the Structure and Diversity of the Rhizosphere Soil Microbial Community of Lilium brownii var. viridulum
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains
2.2. Experimental Design
2.3. Rhizosphere Soil Sample Collection
2.4. Determination of Rhizosphere Soil Physicochemical Properties
2.5. DNA Extraction, PCR Amplification, and High-Throughput Sequencing
2.6. Data Analysis
3. Results and Discussion
3.1. Biocontrol Activity of Bacillus BF1 and Y37 against Fusarium Wilt of Lily
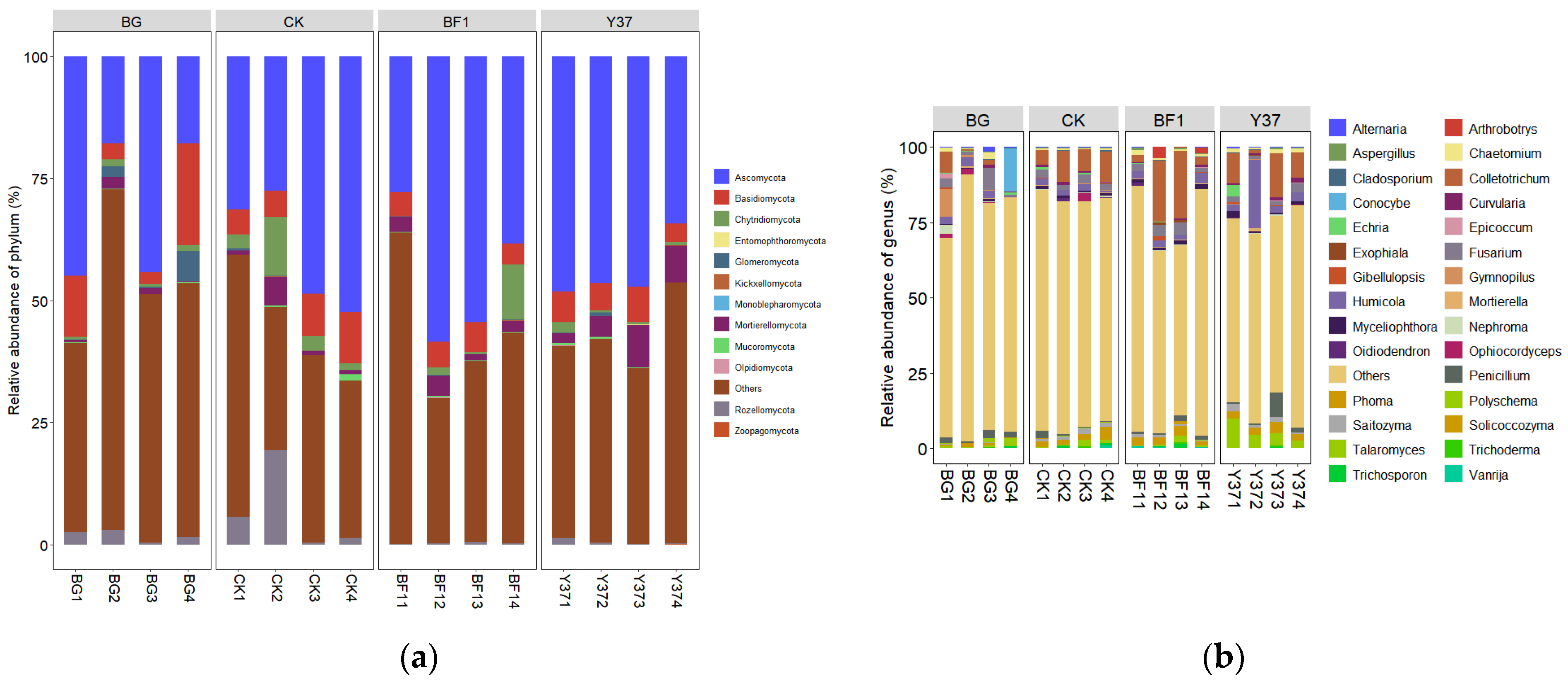
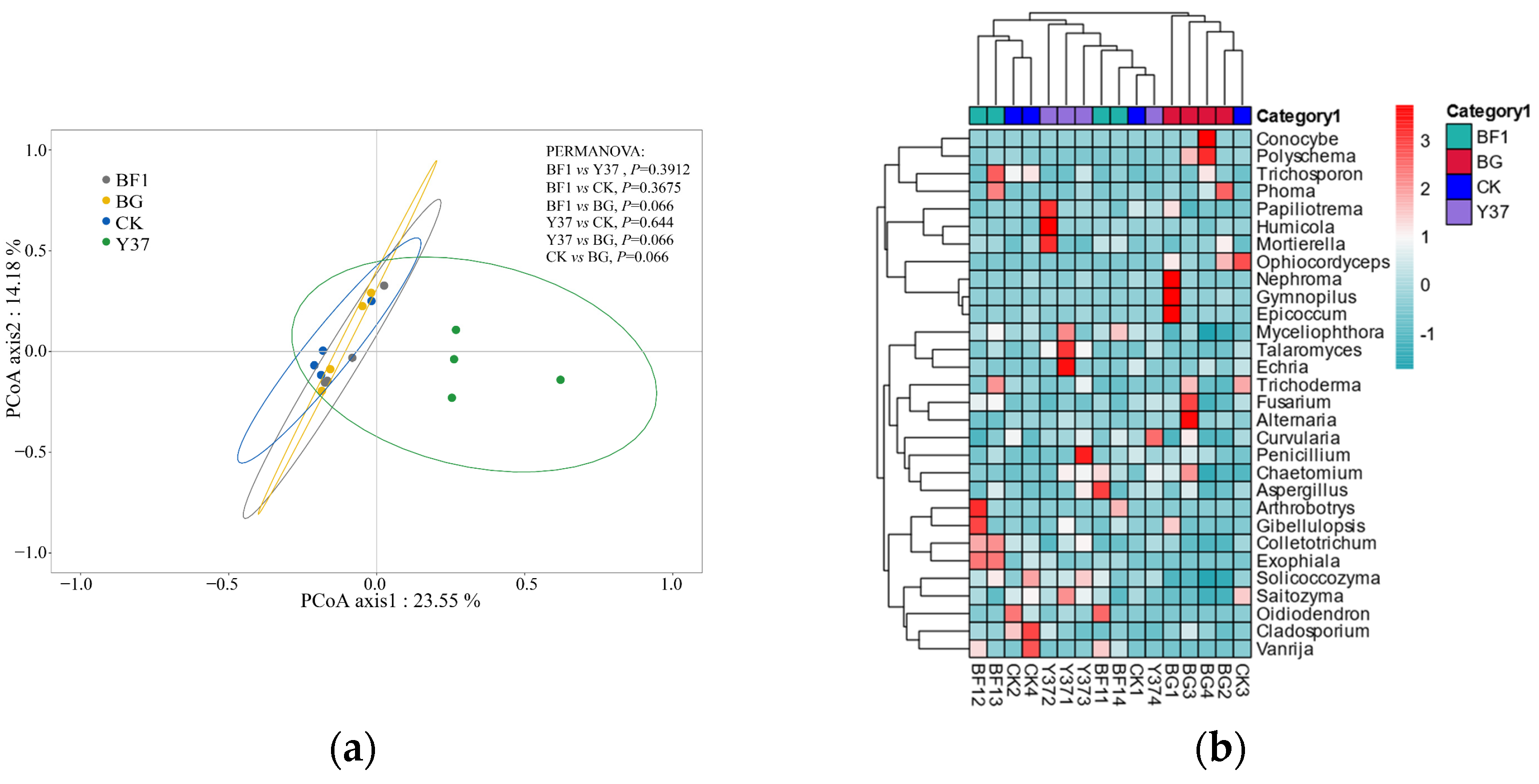
3.2. Effects of Bacillus BF1 and Y37 on Fungal Community Structure and Diversity in the Rhizosphere Soil
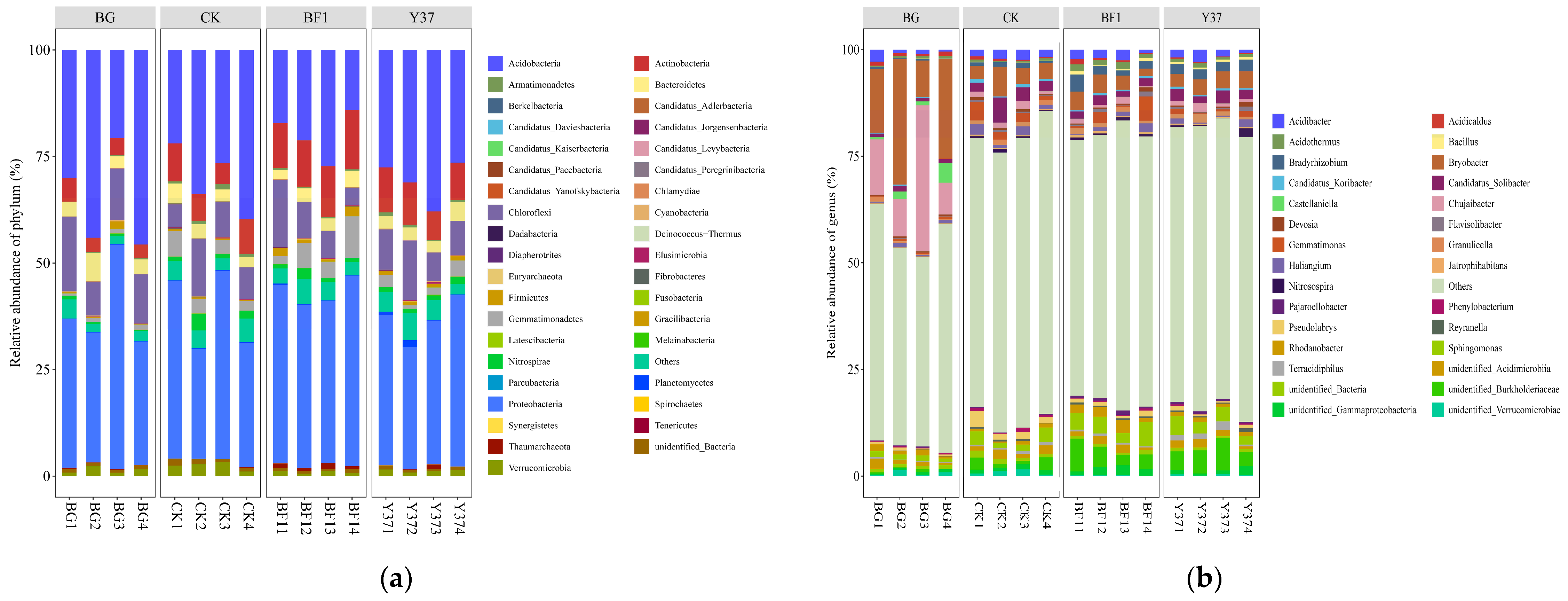
3.3. Effects of Bacillus BF1 and Y37 on Bacterial Community Structure and Diversity in the Rhizosphere Soil
3.4. Effects of Bacillus BF1 and Y37 on Soil Physicochemical Properties
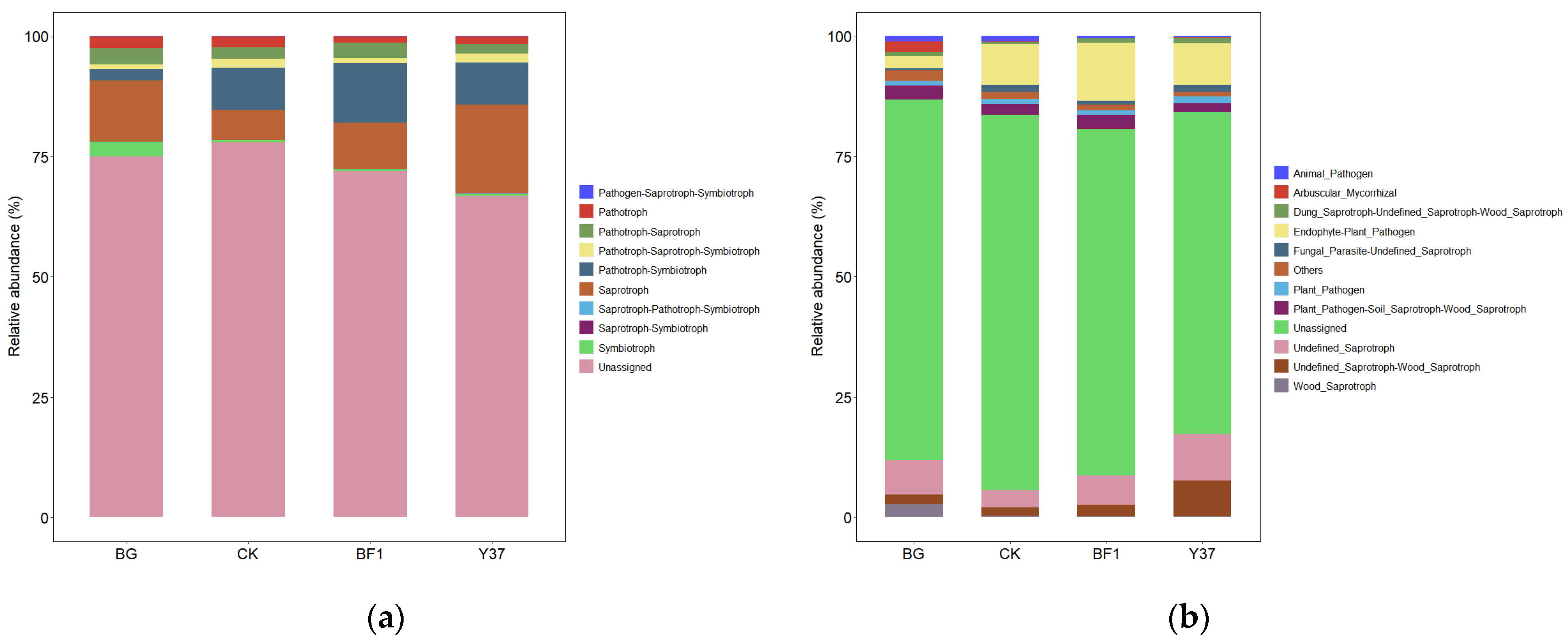
3.5. Predictive Functional Profiling of the Rhizosphere Soil Microbial Community
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munafo, J.P., Jr.; Gianfagna, T.J. Chemistry and biological activity of steroidal glycosides from the Lilium genus. Nat. Prod. Rep. 2015, 32, 454–477. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-C.; Liu, Y.-J.; He, G.-R.; Cao, Y.-W.; Bi, M.-M.; Song, M.; Yang, P.-P.; Xu, L.-F.; Ming, J. Comprehensive Analysis of Secondary Metabolites in the Extracts from Different Lily Bulbs and Their Antioxidant Ability. Antioxidants 2021, 10, 1634. [Google Scholar] [CrossRef] [PubMed]
- Akram, W.; Ahmad, A.; Yasin, N.A.; Anjum, T.; Ali, B.; Fatima, S.; Ahmed, S.; Simirgiotis, M.J.; Li, G. Mechanical strengthening and metabolic re-modulations are involved in protection against Fusarium wilt of tomato by B. subtilis IAGS174. J. Plant Interact. 2021, 16, 411–421. [Google Scholar] [CrossRef]
- Xu, W.; Yang, Q.; Yang, F.; Xie, X.; Goodwin, P.H.; Deng, X.; Tian, B.; Yang, L. Evaluation and Genome Analysis of Bacillus subtilis YB-04 as a Potential Biocontrol Agent Against Fusarium Wilt and Growth Promotion Agent of Cucumber. Front. Microbiol. 2022, 13, 885430. [Google Scholar] [CrossRef]
- Ingram, R.J.; Levy, F. Identity and symptomatology of a newly described lily leaf spot disease (Pseudocercosporella inconspicua) of Gray’s lily (Lilium grayi). Can. J. Plant Pathol. 2020, 42, 499–507. [Google Scholar] [CrossRef]
- Kaminska, M.; Dziekanowska, D. Molecular evidence for the presence of aster yellows-related phytoplasmas in lilies with leaf scorch and flower virescence. J. Phytopathol. 2002, 150, 90–93. [Google Scholar] [CrossRef]
- Kaur, H.; Garg, H. Pesticides: Environmental Impacts and Management Strategies; Intech: London, UK, 2014. [Google Scholar]
- Posada, L.F.; Ramírez, M.; Ochoa-Gómez, N.; Cuellar-Gaviria, T.Z.; Argel-Roldan, L.E.; Ramírez, C.A.; Villegas-Escobar, V. Bioprospecting of aerobic endospore-forming bacteria with biotechnological potential for growth promotion of banana plants. Sci. Hortic. 2016, 212, 81–90. [Google Scholar] [CrossRef]
- Liu, Z.; Xiao, J.; Zhang, X.; Dou, S.; Gao, T.; Wang, D.; Zhang, D. Influence of Bacillus subtilis strain Z-14 on microbial communities of wheat rhizospheric soil infested with Gaeumannomyces graminis var. tritici. Front. Microbiol. 2022, 13, 923242. [Google Scholar] [CrossRef]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef]
- Wang, M.; Xing, Y.; Wang, J.; Xu, Y.; Wang, G. The role of the chi1 gene from the endophytic bacteria Serratia proteamaculans 336x in the biological control of wheat take-all. Can. J. Microbiol. 2014, 60, 533–540. [Google Scholar] [CrossRef]
- Tian, L.; Shi, S.; Ji, L.; Nasir, F.; Ma, L.; Tian, C. Effect of the biocontrol bacterium Bacillus amyloliquefaciens on the rhizosphere in ginseng plantings. Int. Microbiol. 2018, 21, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Win, K.T.; Okazaki, K.; Ohkama-Ohtsu, N.; Yokoyama, T.; Ohwaki, Y. Short-term effects of biochar and Bacillus pumilus TUAT-1 on the growth of forage rice and its associated soil microbial community and soil properties. Biol. Fertil. Soils 2020, 56, 481–497. [Google Scholar] [CrossRef]
- James, D.B. Soil community feedback and the coexistence of competitors: Conceptual frameworks and empirical tests. New Phytol. 2003, 157, 465–473. [Google Scholar] [CrossRef]
- Palmieri, D.; Vitullo, D.; De Curtis, F.; Lima, G. A microbial consortium in the rhizosphere as a new biocontrol approach against fusarium decline of chickpea. Plant Soil 2016, 412, 425–439. [Google Scholar] [CrossRef]
- Holl, F.B.; Chanway, C.P. Rhizosphere colonization and seedling growth promotion of lodgepole pine byBacillus polymyxa. Can. J. Microbiol. 1992, 38, 303–308. [Google Scholar] [CrossRef]
- Hatzinger, P.B.; Alexander, M. Relationship between the number of bacteria added to soil or seeds and their abundance and distribution in the rhizosphere of alfalfa. Plant Soil 1994, 158, 211–222. [Google Scholar] [CrossRef]
- Shi, L.; Liu, N.; Liu, G.; Fang, J. Bacterial Community Structure and Dynamic Changes in Different Functional Areas of a Piggery Wastewater Treatment System. Microorganisms 2021, 9, 2134. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis; SSSA Book Series; John Wiley & Sons, Inc.: New York, NY, USA, 1996; pp. 961–1010. [Google Scholar]
- Sun, S.; Zhang, J.; Cai, C.; Cai, Z.; Li, X.; Wang, R. Coupling of non-point source pollution and soil characteristics covered by Phyllostachys edulis stands in hilly water source area. J. Environ. Manag. 2020, 268, 110657. [Google Scholar] [CrossRef]
- Pahlich, E.; Gerlitz, C. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemistry 1980, 19, 11–13. [Google Scholar] [CrossRef]
- Jiao, S.; Liu, Z.S.; Lin, Y.B.; Yang, J.; Chen, W.M.; Wei, G.H. Bacterial communities in oil contaminated soils: Biogeography and co-occurrence patterns. Soil Biol. Biochem. 2016, 98, 64–73. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Kechin, A.; Boyarskikh, U.; Kel, A.; Filipenko, M. cutPrimers: A New Tool for Accurate Cutting of Primers from Reads of Targeted Next Generation Sequencing. J. Comput. Biol. 2017, 24, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Allard-Massicotte, R.; Tessier, L.; Lecuyer, F.; Lakshmanan, V.; Lucier, J.-F.; Garneau, D.; Caudwell, L.; Vlamakis, H.; Bais, H.P.; Beauregard, P.B. Bacillus subtilis Early Colonization of Arabidopsis thaliana Roots Involves Multiple Chemotaxis Receptors. mBio 2016, 7, e01664-16. [Google Scholar] [CrossRef]
- Jiao, R.; Munir, S.; He, P.F.; Yang, H.W.; Wu, Y.X.; Wang, J.W.; He, P.B.; Cai, Y.Z.; Wang, G.; He, Y.Q. Biocontrol potential of the endophytic Bacillus amyloliquefaciens YN201732 against tobacco powdery mildew and its growth promotion. Biol. Control 2020, 143, 104160. [Google Scholar] [CrossRef]
- Cascioferro, S.; Cusimano, M.G.; Schillaci, D. Antiadhesion agents against Gram-positive pathogens. Future Microbiol. 2014, 9, 1209–1220. [Google Scholar] [CrossRef]
- Maung, C.E.H.; Choi, T.G.; Nam, H.H.; Kim, K.Y. Role of Bacillus amyloliquefaciens Y1 in the control of Fusarium wilt disease and growth promotion of tomato. Biocontrol Sci. Technol. 2017, 27, 1400–1415. [Google Scholar] [CrossRef]
- Zhu, J.; Tan, T.; Shen, A.; Yang, X.; Yu, Y.; Gao, C.; Li, Z.; Yi, C.; Chen, J.; Guo, L.; et al. Biocontrol potential of Bacillus subtilis IBFCBF-4 against Fusarium wilt of watermelon. J. Plant Pathol. 2020, 102, 433–441. [Google Scholar] [CrossRef]
- Yu, X.; Ai, C.; Xin, L.; Zhou, G. The siderophore-producing bacterium, Bacillus subtilis CAS15, has a biocontrol effect on Fusarium wilt and promotes the growth of pepper. Eur. J. Soil Biol. 2011, 47, 138–145. [Google Scholar] [CrossRef]
- Tan, T.; Zhu, J.; Shen, A.; Li, J.; Yu, Y.; Zhang, M.; Zhao, M.; Li, Z.; Chen, J.; Gao, C.; et al. Isolation and identification of a Bacillus subtilis HZ-72 exhibiting biocontrol activity against flax seedling blight. Eur. J. Plant Pathol. 2018, 153, 825–836. [Google Scholar] [CrossRef]
- Kroeber, M.; Wibberg, D.; Grosch, R.; Eikmeyer, F.; Verwaaijen, B.; Chowdhury, S.P.; Hartmann, A.; Puehler, A.; Schlueter, A. Effect of the strain Bacillus amyloliquefaciens FZB42 on the microbial community in the rhizosphere of lettuce under field conditions analyzed by whole rnetagenome sequencing. Front. Microbiol. 2014, 5, 252. [Google Scholar] [CrossRef]
- Araujo, R.; Dunlap, C.; Franco, C.M.M. Analogous wheat root rhizosphere microbial successions in field and greenhouse trials in the presence of biocontrol agents Paenibacillus peoriae SP9 and Streptomyces fulvissimus FU14. Mol. Plant Pathol. 2020, 21, 622–635. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.E.; Mejia, L.C.; Kyllo, D.; Rojas, E.I.; Maynard, Z.; Robbins, N.; Herre, E.A. Fungal endophytes limit pathogen damage in a tropical tree. Proc. Natl. Acad. Sci. USA 2003, 100, 15649–15654. [Google Scholar] [CrossRef]
- Herre, E.A.; Mejía, L.C.; Kyllo, D.A.; Rojas, E.; Maynard, Z.; Butler, A.; Van Bael, S.A. Ecological implications of anti-pathogen effects of tropical fungal endophytes and mycorrhizae. Ecology 2007, 88, 550–558. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; Mohamed, S.G.A.; Altyar, A.E.; Mohamed, G.A. Natural Products of the Fungal Genus Humicola: Diversity, Biological Activity, and Industrial Importance. Curr. Microbiol. 2021, 78, 2488–2509. [Google Scholar] [CrossRef]
- Chavez, R.; Bull, P.; Eyzaguirre, J. The xylanolytic enzyme system from the genus Penicillium. J. Biotechnol. 2006, 123, 413–433. [Google Scholar] [CrossRef]
- Zhai, M.M.; Li, J.E.; Jiang, C.X.; Shi, Y.P.; Di, D.L.; Crews, P.; Wu, Q.X. The Bioactive Secondary Metabolites from Talaromyces species. Nat. Prod. Bioprospect. 2016, 6, 1–24. [Google Scholar] [CrossRef]
- Han, L.; Wang, Z.; Li, N.; Wang, Y.; Feng, J.; Zhang, X. Bacillus amyloliquefaciens B1408 suppresses Fusarium wilt in cucumber by regulating the rhizosphere microbial community. Appl. Soil Ecol. 2019, 136, 55–66. [Google Scholar] [CrossRef]
- You, C.; Zhang, C.; Kong, F.; Feng, C.; Wang, J. Comparison of the effects of biocontrol agent Bacillus subtilis and fungicide metalaxyl–mancozeb on bacterial communities in tobacco rhizospheric soil. Ecol. Eng. 2016, 91, 119–125. [Google Scholar] [CrossRef]
- Zhao, W.; Guo, Q.; Li, S.; Lu, X.; Dong, L.; Wang, P.; Zhang, X.; Su, Z.; Ma, P. Application of Bacillus subtilis NCD-2 can suppress cotton verticillium wilt and its effect on abundant and rare microbial communities in rhizosphere. Biol. Control 2022, 165, 104812. [Google Scholar] [CrossRef]
- Chen, W.; Wu, Z.; Liu, C.; Zhang, Z.; Liu, X. Biochar combined with Bacillus subtilis SL-44 as an eco-friendly strategy to improve soil fertility, reduce Fusarium wilt, and promote radish growth. Ecotoxicol. Environ. Saf. 2023, 251, 114509. [Google Scholar] [CrossRef] [PubMed]
- Sennett, L.B.; Goyer, C.; Burton, D.L.; Zebarth, B.J.; Whitney, S. Chemical fumigation and biofumigation alter soil bacterial community diversity and composition. FEMS Microbiol. Ecol. 2022, 98, fiac026. [Google Scholar] [CrossRef]
- Chen, R.; Jiang, W.; Xu, S.; Fan, H.; Chen, X.; Shen, X.; Yin, C.; Mao, Z. An emerging chemical fumigant: Two-sided effects of dazomet on soil microbial environment and plant response. Environ. Sci. Pollut. Res. 2022, 29, 3022–3036. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, T.; Zheng, Z.; Chen, H.Y.H. Soil aggregate-associated bacterial metabolic activity and community structure in different aged tea plantations. Sci. Total Environ. 2019, 654, 1023–1032. [Google Scholar] [CrossRef]
- Wan, T.; Zhao, H.; Wang, W. Effect of biocontrol agent Bacillus amyloliquefaciens SN16-1 and plant pathogen Fusarium oxysporum on tomato rhizosphere bacterial community composition. Biol. Control 2017, 112, 1–9. [Google Scholar] [CrossRef]
- Tanwar, A.; Aggarwal, A.; Parkash, V. Effect of bioinoculants and superphosphate fertilizer on the growth and yield of broccoli (Brassica oleracea L. var. italica Plenck). N. Z. J. Crop Hortic. Sci. 2014, 42, 288–302. [Google Scholar] [CrossRef]
- Yildirim, E.; Karlidag, H.; Turan, M.; Dursun, A.; Goktepe, F. Growth, Nutrient Uptake, and Yield Promotion of Broccoli by Plant Growth Promoting Rhizobacteria with Manure. Hortscience 2011, 46, 932–936. [Google Scholar] [CrossRef]
- Hu, Y.; Li, Y.; Yang, X.; Li, C.; Wang, L.; Feng, J.; Chen, S.; Li, X.; Yang, Y. Effects of integrated biocontrol on bacterial wilt and rhizosphere bacterial community of tobacco. Sci. Rep. 2021, 11, 2653. [Google Scholar] [CrossRef]
- Shi, G.; Sun, H.; Calderon-Urrea, A.; Jia, X.; Yang, H.; Su, G. Soil Fungal Diversity Loss and Appearance of Specific Fungal Pathogenic Communities Associated With the Consecutive Replant Problem (CRP) in Lily. Front. Microbiol. 2020, 11, 1649. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhang, L.; Li, Y.; Hou, L.; Yang, H.; Shi, G. Silicon Fertilizer and Microbial Agents Changed the Bacterial Community in the Consecutive Replant Soil of Lilies. Agronomy 2022, 12, 1530. [Google Scholar] [CrossRef]
- Wang, T.; Cheng, K.; Huo, X.; Meng, P.; Cai, Z.; Wang, Z.; Zhou, J. Bioorganic fertilizer promotes pakchoi growth and shapes the soil microbial structure. Front. Plant Sci. 2022, 13, 1040437. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Shi, W.; Wu, D.; Tian, R.; Wang, B.; Lin, R.; Zhou, B.; Gao, Z. Biocontrol of potato common scab by Brevibacillus laterosporus BL12 is related to the reduction of pathogen and changes in soil bacterial community. Biol. Control 2021, 153, 104496. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Growth Indices | Yield (kg) | DI (%) | DR (%) | ||
|---|---|---|---|---|---|---|
| Height (cm) | Stem Thickness (cm) | Fresh Weight (g) | ||||
| BG | – | – | – | – | – | – |
| CK | 73.80 ± 6.38c | 8.07 ± 0.98b | 24.00 ± 8.61b | 8.67 ± 3.25b | 76.30 ± 4.63a | – |
| BF1 | 85.40 ± 1.14b | 9.76 ± 0.43a | 37.61 ± 10.83b | 11.21 ± 3.31b | 31.48 ± 3.90b | 58.74 ± 5.11 |
| Y37 | 96.20 ± 7.85a | 10.66 ± 0.73a | 60.07 ± 14.85a | 17.73 ± 5.09a | 23.70 ± 3.39b | 68.93 ± 4.45 |
| Treatment | Richness | Shannon | Simpson | ACE | Chao1 | Coverage (%) |
|---|---|---|---|---|---|---|
| BG | 638.50 ± 126.33b | 6.45 ± 0.48a | 0.966 ± 0.01a | 762.70 ± 124.66b | 780.30 ± 117.39b | 99.62 ± 0.00a |
| CK | 835.25 ± 90.15a | 6.87 ± 0.22a | 0.969 ± 0.01a | 1002.44 ± 96.53a | 1042.46 ± 99.65a | 99.48 ± 0.00b |
| BF1 | 843.25 ± 84.83a | 6.33 ± 0.69a | 0.936 ± 0.05a | 1020.11 ± 70.96a | 1027.33 ± 78.74a | 99.46 ± 0.00b |
| Y37 | 837.50 ± 40.17a | 6.62 ± 0.44a | 0.959 ± 0.02a | 1004.25 ± 73.17a | 1012.29 ± 86.99a | 99.47 ± 0.00b |
| Treatment | Richness | Shannon | Simpson | ACE | Chao1 | Coverage (%) |
|---|---|---|---|---|---|---|
| BG | 1186 ± 97c | 6.79 ± 0.08c | 0.943 ± 0.02b | 1521 ± 90c | 1531 ± 102c | 98.82 ± 0.08a |
| CK | 1469 ± 112b | 8.35 ± 0.26b | 0.991 ± 0.00a | 1829 ± 119b | 1851 ± 126b | 98.66 ± 0.07ab |
| BF1 | 1755 ± 142a | 8.74 ± 0.17a | 0.994 ± 0.00a | 2165 ± 216a | 2170 ± 235a | 98.42 ± 0.22c |
| Y37 | 1575 ± 182ab | 8.44 ± 0.33ab | 0.992 ± 0.00a | 1971 ± 207ab | 1982 ± 230ab | 98.54 ± 0.14bc |
| Treatment | Bacillus Genus Relative Abundance (%) |
|---|---|
| BG | 0.057 ± 0.028b |
| CK | 0.091 ± 0.044b |
| BF1 | 0.456 ± 0.295a |
| Y37 | 0.438 ± 0.103a |
| BG | CK | BF1 | Y37 | |
|---|---|---|---|---|
| pH | 7.06 ± 0.041ab | 7.02 ± 0.017bc | 7.07 ± 0.040a | 7.01 ± 0.012c |
| TN (g/kg) | 1.05 ± 0.303a | 1.15 ± 0.305a | 0.78 ± 0.096a | 0.78 ± 0.042a |
| TK (%) | 1.70 ± 0.002a | 1.72 ± 0.001a | 1.56 ± 0.001a | 1.51 ± 0.001a |
| TP (μg/g) | 1338.356 ± 170.327b | 902.740 ± 176.785c | 1873.973 ± 174.197a | 1793.151 ± 172.545a |
| NH4+-N (μg/g) | 12.0090 ± 2.594b | 8.1571 ± 0.489c | 18.6556 ± 2.710a | 19.6941 ± 1.751a |
| NO3−-N (μg/g) | 201.373 ± 10.083c | 174.759 ± 12.734d | 273.429 ± 14.243a | 228.978 ± 12.465b |
| AK (mg/kg) | 82.620 ± 1.118b | 67.686 ± 11.880c | 94.904 ± 2.944a | 91.071 ± 3.585ab |
| AP (μmol/g) | 24.767 ± 1.682b | 14.754 ± 1.561c | 36.259 ± 3.706a | 37.740 ± 1.818a |
| OP (μg/g) | 618.912 ± 119.298ab | 405.518 ± 265.598b | 811.473 ± 184.833a | 804.262 ± 175.493a |
| OC (g/kg) | 23.65 ± 2.073a | 24.02 ± 4.116a | 25.09 ± 5.774a | 24.84 ± 3.964a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, J.; Zhao, X.; Yang, Y.; Yi, Y.; Wang, H.; Wei, B.; Zeng, L. Two Bacillus spp. Strains Improve the Structure and Diversity of the Rhizosphere Soil Microbial Community of Lilium brownii var. viridulum. Microorganisms 2023, 11, 1229. https://doi.org/10.3390/microorganisms11051229
Tu J, Zhao X, Yang Y, Yi Y, Wang H, Wei B, Zeng L. Two Bacillus spp. Strains Improve the Structure and Diversity of the Rhizosphere Soil Microbial Community of Lilium brownii var. viridulum. Microorganisms. 2023; 11(5):1229. https://doi.org/10.3390/microorganisms11051229
Chicago/Turabian StyleTu, Jing, Xin Zhao, Yuanru Yang, Yongjian Yi, Hongying Wang, Baoyang Wei, and Liangbin Zeng. 2023. "Two Bacillus spp. Strains Improve the Structure and Diversity of the Rhizosphere Soil Microbial Community of Lilium brownii var. viridulum" Microorganisms 11, no. 5: 1229. https://doi.org/10.3390/microorganisms11051229
APA StyleTu, J., Zhao, X., Yang, Y., Yi, Y., Wang, H., Wei, B., & Zeng, L. (2023). Two Bacillus spp. Strains Improve the Structure and Diversity of the Rhizosphere Soil Microbial Community of Lilium brownii var. viridulum. Microorganisms, 11(5), 1229. https://doi.org/10.3390/microorganisms11051229