
Anti-Insect Properties of Penicillium Secondary Metabolites

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Penicillium Species as Insect Associates
3. Effects on Insect Viability and Development
3.1. Crude Culture Extracts
3.2. Purified Compounds
3.2.1. Mycotoxins
3.2.2. Other Products
4. Effects on the Nervous System
4.1. Acetylcholinesterase Inhibitors

{kind=link}
{kind=link}
{kind=link}
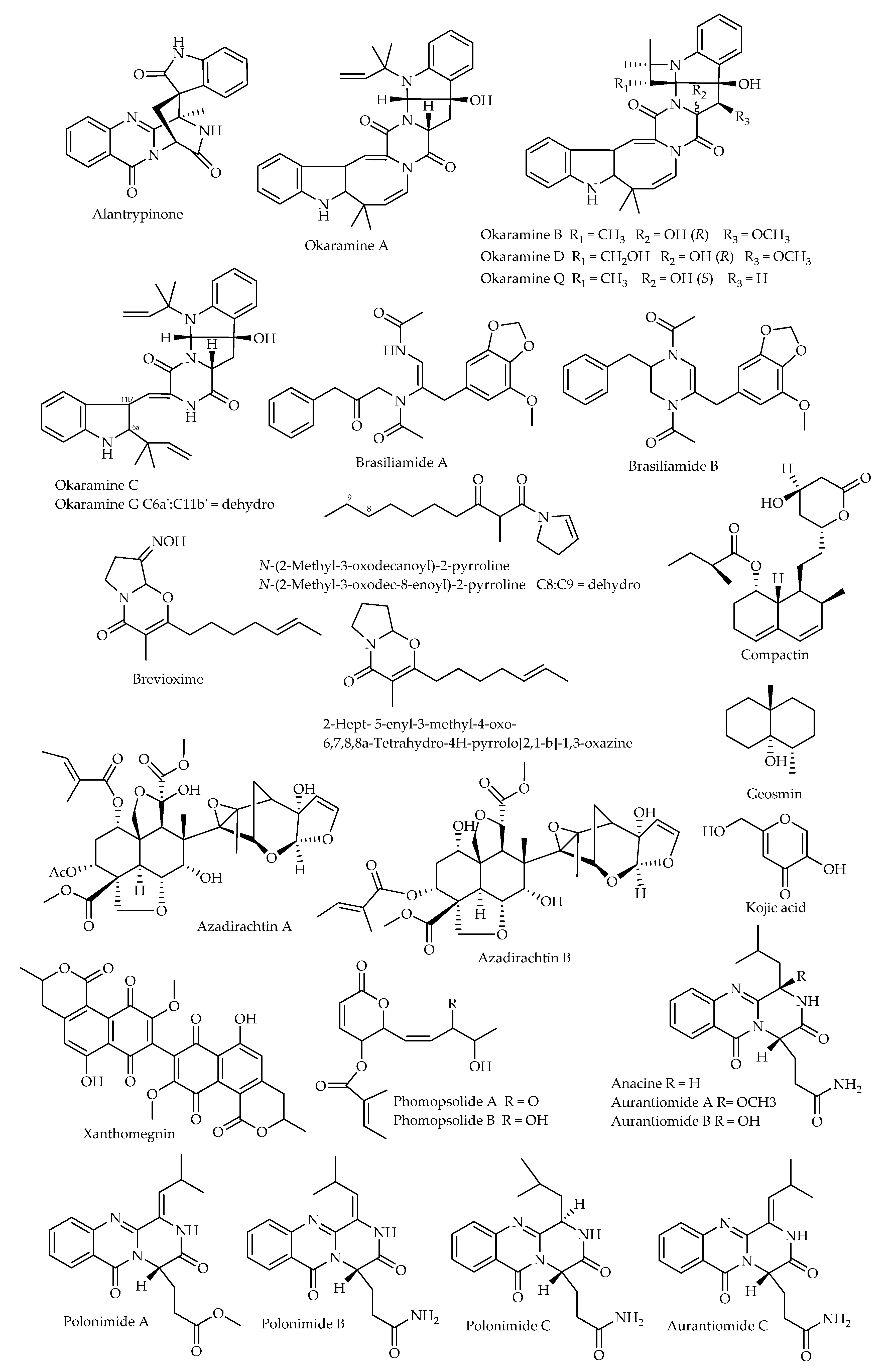{kind=link}
| Secondary Metabolite | Method | Activity | References |
|---|---|---|---|
| Aloe-emodin | Ellman | 42.5 µg/mL (IC50) | [147] |
| Arisugacin A | Ellman | 0.001 µM (IC50) | [133,148,149] |
| Arisugacin B | Ellman Ellman | 0.0258 µM (IC50) 3.03 µM (IC50) | [148,149] [150] |
| Arisugacin C | Ellman Ellman * | 2.5 µM (IC50) 1.4 µM (IC50) | [149] [122] |
| Arisugacin D | Ellman Ellman | 3.5 µM (IC50) 53.39 µM (IC50) | [149] [150] |
| Arisugacin F | Ellman | 0.37 µM (IC50) | [151] |
| Arisugacin I | Ellman | 0.64 µM (IC50) | [151] |
| Arisugacin L | Ellman * | 0.191 µM (IC50) | [122] |
| Arisugacin N | Ellman * | 3.9 µM (IC50) | [122] |
| Arisugacin O | Ellman * | 4.6 µM (IC50) | [122] |
| Arisugacin P | Ellman * | 66 µM (IC50) | [122] |
| 5-Bromosclerotiorin | Ellman | 200 µg/mL (23.87%) | [152] |
| Citreorosein | Ellman | 40.5 µg/mL (IC50) | [147] |
| Cyclo-(l-Pro–l-Val) | Marston | 10.0 µg (MIR) | [153] |
| Cyclopenin | Ellman | 2.04 µM (IC50) | [148] |
| Dechloroisochromophilone II | Marston | 10 ng (MIR) | [154] |
| 3′′-Deoxy-6′-O-desmethylcandidusin B | Ellman | 7.8 µM (IC50) | [155] |
| 6′-O-Desmethylcandidusin B | Ellman | 5.2 µM (IC50) | [155] |
| Dicitrinin A | Ellman modified | 42.0 µM (MIC) | [156] |
| (3R,4R)-3,4-Dihydro-4,6-dihydroxy-3-methyl-1-oxo-1H-isochromene-5-carboxylic acid | Marston | 3.0 µg (MIR) | [157] |
| (3R,4R)-4,7-Dihydroxymellein | Marston | 10.0 µg (MIR) | [157] |
| 4-(5,7-Dimethoxy-4-oxo-4H-chromen-2-yl)butanoic acid | Ellman | 93.2 µM (IC50) | [158] |
| 3-(5,7-Dimethoxy-4-oxo-4H-chromen-2-yl)propanoic acid | Ellman | 50.8 µM (IC50) | [158] |
| 3-Epiarisugacin E | Ellman | 38.23 µM (IC50) | [150] |
| 4-Hydroxymellein | Marston | 30.0 µg (MIR) | [153] |
| (R)-7-Hydroxymellein | Marston | 10.0 µg (MIR) | [157] |
| 13-Hydroxypalitantin | Ellman | 12 µM (IC50) | [142] |
| Isochromophilone II | Marston | 50 ng (MIR) | [154] |
| Isochromophilone III | Marston | 10 ng (MIR) | [154] |
| Isochromophilone IV | Marston | 100 ng (MIR) | [154] |
| Isochromophilone VIII | Marston | 50 ng (MIR) | [154] |
| Isocyclocitrinol B | Ellman modified | 166.0 µM (MIC) | [156] |
| Maritimin | Ellman | 75.3 µM (IC50) | [158] |
| Ochrephilone | Marston | 50 ng (MIR) | [154] |
| Orcinol | Marston | 60.0 µg (MIR) | [153] |
| (+)-Palitantin | Ellman | 0.079 µM (IC50) | [142] |
| Penaloidine A | Ellman | 14.85 µM (IC50) | [145] |
| Penaloidine B | Ellman | 41.27 uM (IC50) | [145] |
| Penicillar B | Ellman | 50 µg/mL (19.5%) | [159] |
| Penicillar C | Ellman | 50 µg/mL (21.3%) | [159] |
| Penicillic acid | Marston | Not determined | [160] |
| (+)-Penicilliumine | Ellman | 50 µM (32.4%) | [143] |
| (−)-Penicilliumine | Ellman | 50 µM (18.7%) | [143] |
| Penicinoline | Ellman | 87.3 µM (IC50) | [161] |
| Penicinoline E | Ellman | 68.5 µM (IC50) | [161] |
| Penicitrinol A | Ellman modified | 23.0 µM (MIC) | [156] |
| Penicitrinone B | Ellman | 38.96 µM (IC50) | [141] |
| Penicitrinone H | Ellman | 23.62 µM (IC50) | [141] |
| Penicnthene | Ellman | 28.03 µM (IC50) | [141] |
| Peniopyranone | Ellman | 0.0152 µM (IC50) | [162] |
| Pileotin B | Ellman | 13.9 µM (IC50) | [163] |
| Pinselin | Ellman | 45.9 µg/mL | [147] |
| Quinolactacin A1 | Ellman | 280 µM (IC50) | [144] |
| Quinolactacin A2 | Ellman | 19.8 µM (IC50) | [144] |
| Sclerotioramine | Ellman | 38.7 µM (IC50) | [158] |
| Sorbiterrin A | not reported | 25 µg/mL (IC50) | [164] |
| Terreulactone C | Ellman | 0.028 µM (IC50) | [150] |
| Territrem A | Ellman | 0.11 µM (IC50) | [165] |
| Territrem B | Ellman Ellman Ellman Ellman | 0.0076 µM (IC50) 0.00703 µM (IC50) 0.047 µM (IC50) 0.00036 µM (IC50) | [133,148] [142] [165] [146] |
| Territrem C | Ellman Ellman Ellman | 0.0068 µM (IC50) 0.23 µM (IC50) 0.045 µM (IC50) | [133,148] [150] [165] |
| Tetrahydrobisvertinolone | Ellman | 50 µg/mL (51.1%) | [166] |
| Tetrahydrotrichodimer ether | Ellman | 50 µg/mL (55.1%) | [166] |
4.2. Antagonists of Acetylcholine and γ-Aminobutyric Acid Receptors
5. Anti-Juvenile Hormone Activity
6. Effects on Immune Response
7. Behavioral Effects
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Berestetskiy, A.; Hu, Q. The chemical ecology approach to reveal fungal metabolites for arthropod pest management. Microorganisms 2021, 9, 1379. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, A.C.; Saari, S.; Moran-Diez, M.E.; Meyling, N.V.; Raad, M.; Glare, T.R. Beauveria bassiana as an endophyte: A critical review on associated methodology and biocontrol potential. BioControl 2017, 62, 1–17. [Google Scholar] [CrossRef]
- Peng, Z.-Y.; Huang, S.-T.; Chen, J.-T.; Li, N.; Wei, Y.; Nawaz, A.; Deng, S.-Q. An update of a green pesticide: Metarhizium anisopliae. All Life 2022, 15, 1141–1159. [Google Scholar] [CrossRef]
- Nicoletti, R.; Becchimanzi, A. Endophytism of Lecanicillium and Akanthomyces. Agriculture 2020, 10, 205. [Google Scholar] [CrossRef]
- Shikano, I. Evolutionary ecology of multitrophic interactions between plants, insect herbivores and entomopathogens. J. Chem. Ecol. 2017, 43, 586–598. [Google Scholar] [CrossRef]
- Francis, F.; Fingu-Mabola, J.C.; Ben Fekih, I. Direct and endophytic effects of fungal entomopathogens for sustainable aphid control: A review. Agriculture 2022, 12, 2081. [Google Scholar] [CrossRef]
- da Costa, G.L.; Lage de Moraes, A.M.; de Oliveira, P.C. Pathogenic action of Penicillium species on mosquito vectors of human tropical diseases. J. Basic Microbiol. 1998, 38, 337–341. [Google Scholar] [CrossRef]
- Ashtekar, N.; Anand, G.; Thulasiram, H.V.; Rajeshkumar, K.C. Genus Penicillium: Advances and application in the modern era. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2021; pp. 201–213. [Google Scholar]
- Nicoletti, R.; Buommino, E.; De Filippis, A.; Lopez-Gresa, M.P.; Manzo, E.; Carella, A.; Petrazzuolo, M.; Tufano, M.A. Bioprospecting for antagonistic Penicillium strains as a resource of new antitumor compounds. World J. Microbiol. Biotechnol. 2008, 24, 189–195. [Google Scholar] [CrossRef]
- Nicoletti, R. Antitumor and Immunomodulatory Compounds from Fungi. In Encyclopedia of Mycology, 1st ed.; CRC Press: Amsterdam, The Netherlands, 2021; pp. 683–709. [Google Scholar]
- Toghueo, R.M.K.; Boyom, F.F. Endophytic Penicillium species and their agricultural, biotechnological, and pharmaceutical applications. 3 Biotech 2020, 10, 107. [Google Scholar] [CrossRef]
- Vinale, F.; Salvatore, M.M.; Nicoletti, R.; Staropoli, A.; Manganiello, G.; Venneri, T.; Borrelli, F.; DellaGreca, M.; Salvatore, F.; Andolfi, A. Identification of the main metabolites of a marine-derived strain of Penicillium brevicompactum using LC and GC MS techniques. Metabolites 2020, 10, 55. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.-B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef]
- Li, X.; Guo, W.; Wen, Y.; Solanki, M.K.; Ding, J. Transmission of symbiotic fungus with a nonsocial leaf-rolling weevil. J. Asia-Pac. Entomol. 2016, 19, 619–624. [Google Scholar] [CrossRef]
- Salvatore, M.M.; Carraturo, F.; Salbitani, G.; Rosati, L.; De Risi, A.; Andolfi, A.; Salvatore, F.; Guida, M.; Carfagna, S. Biological and metabolic effects of the association between the microalga Galdieria sulphuraria and the fungus Penicillium citrinum. Sci. Rep. 2023, 13, 1789. [Google Scholar] [CrossRef]
- Yilmaz, N.; Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of the genus Talaromyces. Stud. Mycol. 2014, 78, 175–341. [Google Scholar] [CrossRef]
- Nicoletti, R.; Becchimanzi, A. Talaromyces–Insect relationships. Microorganisms 2022, 10, 45. [Google Scholar] [CrossRef]
- Poitevin, C.G.; Porsani, M.V.; Poltronieri, A.S.; Zawadneak, M.A.C.; Pimentel, I.C. Fungi isolated from insects in strawberry crops act as potential biological control agents of Duponchelia fovealis (Lepidoptera: Crambidae). Appl. Entomol. Zool. 2018, 53, 323–331. [Google Scholar] [CrossRef]
- Teixeira, M.F.N.P.; Souza, C.R.; Morais, P.B. Diversity and enzymatic capabilities of fungi associated with the digestive tract of larval stages of a shredder insect in cerrado and Amazon forest, Brazil. Braz. J. Biol. 2022, 82, e260039. [Google Scholar] [CrossRef] [PubMed]
- Gambino, P.; Thomas, G.M. Fungi associated with two Vespula (Hymenoptera: Vespidae) species. Pan-Pac. Entomol. 1988, 64, 107–113. [Google Scholar]
- Brooks, D.L. Entomogenous Fungi of Corn Insects in Iowa; Iowa State University: Ames, Iowa, 1962. [Google Scholar]
- Rashad, E.M.; Rashed, S.S.; Helal, G.A.; Hashem, F.M. Influence of Penicillium aurantiogriseum and its mycotoxin citrinin on haemolymph of Schistocerca gregaria (Forskål). Middle East J. Appl. Sci. 2020, 10, 673–682. [Google Scholar]
- Ismail, M.A.; Abdel-Sater, M.A. Fungi associated with the Egyptian cotton leafworm Spodoptera littoralis Boisdoval. Mycopathologia 1993, 124, 79–86. [Google Scholar] [CrossRef]
- Moraes, A.M.L.; Junqueira, A.C.V.; Costa, G.L.; Celano, V.; Oliveira, P.C.; Coura, J.R. Fungal flora of the digestive tract of 5 species of triatomines vectors of Trypanosoma cruzi, Chagas 1909. Mycopathologia 2001, 151, 41–48. [Google Scholar] [CrossRef]
- Heo, I.; Hong, K.; Yang, H.; Lee, H.B.; Choi, Y.-J.; Hong, S.-B. Diversity of Aspergillus, Penicillium, and Talaromyces species isolated from freshwater environments in Korea. Mycobiology 2019, 47, 12–19. [Google Scholar] [CrossRef]
- Turesson, G. The toxicity of moulds to the honeybee and the cause of bee paralysis. Svensk. Bot. Tidskr. 1917, 11, 33–38. [Google Scholar]
- Park, J.M.; You, Y.-H.; Back, C.-G.; Kim, H.-H.; Ghim, S.-Y.; Park, J.-H. Fungal load in Bradysia agrestis, a phytopathogen-transmitting insect vector. Symbiosis 2018, 74, 145–158. [Google Scholar] [CrossRef]
- Müller-Kögler, E.; Huger, A. Wundinfektionen Bei Raupen von Malacosoma neustria (L.) Durch Penicillium brevi-compactum Dierckx. Zeit. Angew. Entomol. 1960, 45, 421–429. [Google Scholar] [CrossRef]
- Peterson, S.W.; Pérez, J.; Vega, F.E.; Infante, F. Penicillium brocae, a new species associated with the coffee berry borer in Chiapas, Mexico. Mycologia 2003, 95, 141–147. [Google Scholar] [CrossRef]
- Visagie, C.M.; Renaud, J.B.; Burgess, K.M.N.; Malloch, D.W.; Clark, D.; Ketch, L.; Urb, M.; Louis-Seize, G.; Assabgui, R.; Sumarah, M.W.; et al. Fifteen new species of Penicillium. Persoonia 2016, 36, 247–280. [Google Scholar] [CrossRef]
- da Costa, G.L.; de Oliveira, P.C. Penicillium species in mosquitoes from two Brazilian regions. J. Basic Microbiol. 1998, 38, 343–347. [Google Scholar] [CrossRef]
- Burnside, C.E. Saprophytic fungi associated with the honey bee. Bee World 1929, 10, 42. [Google Scholar] [CrossRef]
- Gilliam, M.; Prest, D.B. Fungi isolated from the intestinal contents of foraging worker honey bees, Apis mellifera. J. Invertebr. Pathol. 1972, 20, 101–103. [Google Scholar] [CrossRef]
- Gilliam, M.; Prest, D.B.; Morton, H.L. Fungi isolated from honey bees, Apis mellifera, fed 2, 4-D and antibiotics. J. Invertebr. Pathol. 1974, 24, 213–217. [Google Scholar] [CrossRef]
- Rodrigues, K.F.; Costa, G.L.; Carvalho, M.P.; Epifanio, R. de A. Evaluation of extracts produced by some tropical fungi as potential cholinesterase inhibitors. World J. Microbiol. Biotechnol. 2005, 21, 1617–1621. [Google Scholar] [CrossRef]
- Yamoah, E.; Jones, E.E.; Weld, R.J.; Suckling, D.M.; Waipara, N.; Bourdôt, G.W.; Hee, A.K.W.; Stewart, A. Microbial population and diversity on the exoskeletons of four insect species associated with gorse (Ulex europaeus L.). Australian J. Entomol. 2008, 47, 370–379. [Google Scholar] [CrossRef]
- Russell, B.M.; Kay, B.H.; Shipton, W. Survival of Aedes aegypti (Diptera: Culicidae) eggs in surface and subterranean breeding sites during the Northern Queensland dry season. J. Med. Entomol. 2001, 38, 441–445. [Google Scholar] [CrossRef]
- Sen, S.K.; Jolly, M.S.; Jammy, T.R. A mycosis in the Indian tasar silkworm, Antheraea mylitta Drury, caused by Penicillium citrinum Thom. J. Invertebr. Pathol. 1970, 15, 1–5. [Google Scholar] [CrossRef]
- Maketon, M.; Amnuaykanjanasin, A.; Kaysorngup, A. A rapid knockdown effect of Penicillium citrinum for control of the mosquito Culex quinquefasciatus in Thailand. World J. Microbiol. Biotechnol. 2014, 30, 727–736. [Google Scholar] [CrossRef]
- Betts, A.D. The Fungi of the Bee-Hive; Cornell University Library: London, UK, 1912. [Google Scholar]
- Moraes, A.M.L.; de Figueiredo, A.R.; Junqueira, A.C.V.; da Costa, G.L.; Aguiar, R.K.; de Oliveira, P.C. Fungal flora of the digestive tract of Panstrongylus megistus (Reduviidae) used for experimental xenodiagnosis of Trypanosoma (Schizotripanum) cruzi Chagas, 1909. Rev. Iberoam. Micol. 2001, 18, 79–82. [Google Scholar]
- Foo, K.; Sathiya Seelan, J.S.; Dawood, M.M. Microfungi associated with Pteroptyx bearni (Coleoptera: Lampyridae) eggs and larvae from Kawang river, Sabah (Northern Borneo). Insects 2017, 8, 66. [Google Scholar] [CrossRef]
- Azeem, M.; Terenius, O.; Rajarao, G.K.; Nagahama, K.; Nordenhem, H.; Nordlander, G.; Borg-Karlson, A.-K. Chemodiversity and biodiversity of fungi associated with the pine weevil Hylobius abietis. Fungal Biol. 2015, 119, 738–746. [Google Scholar] [CrossRef]
- Vecchi, M.A. La microflora dell’ape mellifica. Microbiol. Enzimol. 1959, 9, 73–86. [Google Scholar]
- Batra, L.R.; Batra, S.W.T.; Bohart, G.E. The mycoflora of domesticated and wild bees (Apoidea). Mycopathol. Mycol. Appl. 1973, 49, 13–44. [Google Scholar] [CrossRef]
- Taniwaki, M.H.; Pitt, J.I.; Iamanaka, B.T.; Massi, F.P.; Fungaro, M.H.P.; Frisvad, J.C. Penicillium excelsum sp. nov from the Brazil nut tree ecosystem in the Amazon basin’. PLoS ONE 2015, 10, e0143189. [Google Scholar] [CrossRef]
- Azeem, M.; Rajarao, G.K.; Nordenhem, H.; Nordlander, G.; Borg-Karlson, A.K. Penicillium expansum volatiles reduce pine weevil attraction to host plants. J. Chem. Ecol. 2013, 39, 120–128. [Google Scholar] [CrossRef]
- Jaber, S.; Mercier, A.; Knio, K.; Brun, S.; Kambris, Z. Isolation of fungi from dead Arthropods and identification of a new mosquito natural pathogen. Parasit. Vectors 2016, 9, 491. [Google Scholar] [CrossRef]
- Marti, G.A.; García, J.J.; Cazau, M.C.; López Lastra, C.C. Fungal flora of the digestive tract of Triatoma infestans (Hemiptera: Reduviidae) from Argentina. Bol. Soc. Argent. Bot. 2007, 42, 175–179. [Google Scholar]
- Rivera, K.G.; Díaz, J.; Chavarría-Díaz, F.; Garcia, M.; Urb, M.; Thorn, R.G.; Louis-Seize, G.; Janzen, D.H.; Seifert, K.A. Penicillium mallochii and P. guanacastens, two new species isolated from Costa Rican caterpillars. Mycotaxon 2012, 119, 315–328. [Google Scholar] [CrossRef]
- Xu, B.; Zou, K.; Cheng, F. Alkaloids from Penicillium oxalicum, a fungus residing in Acrida cinerea. Adv. Mater. Res. 2014, 881–883, 442–445. [Google Scholar] [CrossRef]
- Kataria, S.K.; Singh, P.; Pandove, G.; Kalia, A.; Chandi, R.S. Penicillum oxalicum Spg1: A novel entomopathogenic fungus isolated from mummified Bemisia tabaci (Gennadius) of cotton. J. Appl. Nat. Sci. 2018, 10, 138–143. [Google Scholar] [CrossRef]
- Lamsal, K.; Kim, S.W.; Naeimi, S.; Adhikari, M.; Yadav, D.R.; Kim, C.; Lee, H.B.; Lee, Y.S. Three new records of Penicillium species isolated from insect specimens in Korea. Mycobiology 2013, 41, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Siedlecki, I.; Gorczak, M.; Okrasińska, A.; Wrzosek, M. Chance or necessity—The fungi co-occurring with Formica polyctena ants. Insects 2021, 12, 204. [Google Scholar] [CrossRef]
- Dângelo, R.A.C.; de Souza, D.J.; Mendes, T.D.; da Cruz Couceiro, J.; Della Lucia, T.M.C. Actinomycetes inhibit filamentous fungi from the cuticle of Acromyrmex leafcutter ants. J. Basic Microbiol. 2016, 56, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Calderón, R.A.; Rivera, G.; Sánchez, L.A.; Zamora, L.G. Chalkbrood (Ascosphaera apis) and some other fungi associated with africanized honey bees (Apis mellifera) in Costa Rica. J. Apic. Res. 2004, 43, 187–188. [Google Scholar] [CrossRef]
- Fernández-Marín, H.; Zimmerman, J.K.; Nash, D.R.; Boomsma, J.J.; Wcislo, W.T. Reduced biological control and enhanced chemical pest management in the evolution of fungus farming in ants. Proc. Royal Soc. B Biol. Sci. 2009, 276, 2263–2269. [Google Scholar] [CrossRef]
- Skou, J.-P.; Nørgaard Holm, S.; Haas, H. Preliminary investigations on diseases in bumble-bees (Bombus Latr.). In Yearbook 1963; Royal Veterinary and Agricultural College: Copenhagen, Denmark, 1963; pp. 27–41. [Google Scholar]
- Lamboni, Y.; Hell, K. Propagation of mycotoxigenic fungi in maize stores by post-harvest insects. Int. J. Trop. Insect Sci. 2009, 29, 31–39. [Google Scholar] [CrossRef]
- Kakinuma, N.; Iwai, H.; Takahashi, S.; Hamano, K.; Yanagisawa, T.; Nagai, K.; Tanaka, K.; Suzuki, K.; Kirikae, F.; Kirikae, T.; et al. Quinolactacins A, B and C: Novel quinolone compounds from Penicillium sp. EPF-6 I. Taxonomy, production, isolation and biological properties. J. Antibiot. 2000, 53, 1247–1251. [Google Scholar] [CrossRef]
- Kobayashi, C.; Fukasawa, Y.; Hirose, D.; Kato, M. Contribution of symbiotic mycangial fungi to larval nutrition of a leaf-rolling weevil. Evol. Ecol. 2008, 22, 711–722. [Google Scholar] [CrossRef]
- Batra, S.W.T. Social Behavior and nests of some nomiine bees in India (Hymenoptera, Halictidæ). Ins. Soc 1966, 13, 145–153. [Google Scholar] [CrossRef]
- Zarrin, M.; Vazirianzadeh, B.; Solary, S.; Mahmoudabadi, A.Z.; Rahdar, M. Isolation of fungi from housefly (Musca domestica) in Ahwaz, Iran. Pak. J. Med. Sci. 2007, 23, 917–919. [Google Scholar]
- Rosa, E.; Ekowati, C.N.; Handayani, T.T.; Ikhsanudin, A.; Apriliani, F.; Arifiyanto, A. Characterization of entomopathogenic fungi as a natural biological control of American cockroaches (Periplaneta americana). Biodiversitas 2020, 21, 5276–5282. [Google Scholar] [CrossRef]
- Henriques, J.; Inácio, L.; Sousa, E. Fungi associated to Platypus cylindrus Fab. (Coleoptera: Platypodidae) in cork oak. Rev. Ciências Agr. 2009, 32, 56–66. [Google Scholar]
- Zoberi, M.H.; Grace, J.K. Fungi associated with the subterranean termite Reticulitermes flavipes in Ontario. Mycologia 1990, 82, 289–294. [Google Scholar] [CrossRef]
- Luz, C.; Tai, M.H.H.; Santos, A.H.; Rocha, L.F.N.; Albernaz, D.A.S.; Silva, H.H.G. Ovicidal activity of entomopathogenic hyphomycetes on Aedes aegypti (Diptera: Culicidae) under laboratory conditions. J. Med. Entomol. 2007, 44, 799–804. [Google Scholar] [CrossRef]
- Kumari, P.; Sivadasan, R.; Jose, A. Microflora associated with the red flour beetle, Tribolium castaneum (Coleoptera: Tenebrionidae). J. Agr. Technol. 2011, 7, 1625–1631. [Google Scholar]
- Balogun, S.A.; Fagade, O.E. Entomopathogenic fungi in population of Zonocerus variegatus (l) in Ibadan, South West, Nigeria. Afr. J. Biotechnol. 2004, 3, 382–386. [Google Scholar]
- Cruz, J.S.; da Costa, G.L.; Villar, J.D.F. Biological activities of crude extracts from Penicillium waksmanii isolated from mosquitoes vectors of tropical diseases. World J. Pharm. Pharm. Sci. 2016, 5, 90–102. [Google Scholar]
- Barbosa, R.N.; Bezerra, J.D.P.; Souza-Motta, C.M.; Frisvad, J.C.; Samson, R.A.; Oliveira, N.T.; Houbraken, J. New Penicillium and Talaromyces species from honey, pollen and nests of stingless bees. Antonie Van Leeuwenhoek 2018, 111, 1883–1912. [Google Scholar] [CrossRef]
- Guswenrivo, I.; Nagao, H.; Lee, C.Y. The diversity of soil fungus in and around termite mounds of Globitermes sulphureus (Haviland) (Blattodea: Termitidae) and response of subterranean termite to fungi. In Sustainable Future for Human Security; Springer: Berlin/Heidelberg, Germany, 2018; pp. 37–52. [Google Scholar]
- Holighaus, G.; Rohlfs, M. Fungal Allelochemicals in insect pest management. Appl. Microbiol. Biotechnol. 2016, 100, 5681–5689. [Google Scholar] [CrossRef]
- Santamarina, M.P.; Roselló, J.; Llacer, R.; Sanchis, V. Antagonistic activity of Penicillium oxalicum Corrie and Thom, Penicillium decumbens Thom and Trichoderma harzianum Rifai isolates against fungi, bacteria and insects. Rev. Iberoam. Micol. 2002, 19, 99–103. [Google Scholar]
- Arunthirumeni, M.; Vinitha, G.; Shivakumar, M.S. Antifeedant and larvicidal activity of bioactive compounds isolated from entomopathogenic fungi Penicillium sp. for the control of agricultural and medically important insect pest (Spodoptera litura and Culex quinquefasciatus). Parasit. Int. 2023, 92, 102688. [Google Scholar] [CrossRef]
- Huang, X.; Luo, J.; Hu, M.; Liang, Y.; Ye, X.; Zhong, G. Isolation and bioactivity of endophytic fungi in Derris hancei. J. South China Agr. Univ. 2009, 30, 44–47. [Google Scholar]
- Hu, M.Y.; Zhong, G.H.; Sun, Z.T.; Sh, G.; Liu, H.M.; Liu, X.Q. Insecticidal activities of secondary metabolites of endophytic Pencillium sp. in Derris elliptica Benth. J. Appl. Entomol. 2005, 129, 413–417. [Google Scholar] [CrossRef]
- Ragavendran, C.; Manigandan, V.; Kamaraj, C.; Balasubramani, G.; Prakash, J.S.; Perumal, P.; Natarajan, D. Larvicidal, histopathological, antibacterial activity of indigenous fungus Penicillium sp. against Aedes aegypti L and Culex quinquefasciatus (Say) (Diptera: Culicidae) and its acetylcholinesterase inhibition and toxicity assessment of zebrafish (Danio rerio). Front. Microbiol. 2019, 10, 427. [Google Scholar] [PubMed]
- Magan, N.; Hope, R.; Cairns, V.; Aldred, D. Post-harvest fungal ecology: Impact of fungal growth and mycotoxin accumulation in stored grain. In Epidemiology of Mycotoxin Producing Fungi; Xu, X., Bailey, J.A., Cooke, B.M., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 723–730. [Google Scholar]
- Berenbaum, M.R.; Bush, D.S.; Liao, L.-H. Cytochrome P450-mediated mycotoxin metabolism by plant-feeding insects. Curr. Opin. Insect Sci. 2021, 43, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Wright, V.F.; Harein, P.K.; Collins, N.A. Preference of the confused flour beetle for certain Penicillium isolates 1. Environ. Entomol. 1980, 9, 213–216. [Google Scholar] [CrossRef]
- Wright, V.F.; De Las Casas, E.; Harein, P.K. Evaluation of Penicillium mycotoxins for activity in stored-product Coleoptera 1. Environ. Entomol. 1980, 9, 217–221. [Google Scholar] [CrossRef]
- Wright, V.F.; Harein, P.K. Effects of some mycotoxins on the Mediterranean flour moth. Environ. Entomol. 1982, 11, 1043–1045. [Google Scholar] [CrossRef]
- Dowd, P.F. Toxicity of naturally occurring levels of the Penicillium mycotoxins citrinin, ochratoxin a, and penicillic acid to the corn earworm, Heliothis zea, and the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Environ. Entomol. 1989, 18, 24–29. [Google Scholar] [CrossRef]
- Dowd, P.F.; Cole, R.J.; Vesonder, R.F. Toxicity of selected tremorgenic mycotoxins and related compounds to Spodoptera frugiperda and Heliothis zea. J. Antib. 1988, 41, 1868–1872. [Google Scholar] [CrossRef]
- González, M.C.; Lull, C.; Moya, P.; Ayala, I.; Primo, J.; Primo Yufera, E. Insecticidal activity of penitrems, including penitrem G, a new member of the family isolated from Penicillium crustosum. J. Agric. Food Chem. 2003, 51, 2156–2160. [Google Scholar] [CrossRef]
- Belofsky, G.N.; Gloer, J.B.; Wicklow, D.T.; Dowd, P.F. Antiinsectan alkaloids: Shearinines A-C and a new paxilline derivative from the ascostromata of Eupenicillium shearii. Tetrahedron 1995, 51, 3959–3968. [Google Scholar] [CrossRef]
- Nicholson, M.J.; Eaton, C.J.; Stärkel, C.; Tapper, B.A.; Cox, M.P.; Scott, B. Molecular cloning and functional analysis of gene clusters for the biosynthesis of indole-diterpenes in Penicillium crustosum and P. janthinellum. Toxins 2015, 7, 2701–2722. [Google Scholar] [CrossRef]
- Babu, J.V.; Popay, A.J.; Miles, C.O.; Wilkins, A.L.; di Menna, M.E.; Finch, S.C. Identification and structure elucidation of janthitrems A and D from Penicillium janthinellum and determination of the tremorgenic and anti-insect activity of janthitrems A and B. J. Agric. Food Chem. 2018, 66, 13116–13125. [Google Scholar] [CrossRef]
- Paterson, R.R.M.; Simmonds, M.S.J.; Blaney, W.M. Mycopesticidal effects of characterized extracts of Penicillium isolates and purified secondary metabolites (including mycotoxins) on Drosophila melanogaster and Spodoptora littoralis. J. Invert. Pathol. 1987, 50, 124–133. [Google Scholar] [CrossRef]
- Paterson, R.R.M.; Simmonds, M.J.S.; Kemmelmeier, C.; Blaney, W.M. Effects of brevianamide A, its photolysis product brevianamide D, and ochratoxin A from two Penicillium strains on the insect pests Spodoptera frugiperda and Heliothis virescens. Mycol. Res. 1990, 94, 538–542. [Google Scholar] [CrossRef]
- Shim, S.H.; Swenson, D.C.; Gloer, J.B.; Dowd, P.F.; Wicklow, D.T. Penifulvin A: A sesquiterpenoid-derived metabolite containing a novel dioxa[5,5,5,6]fenestrane ring system from a fungicolous isolate of Penicillium griseofulvum. Org. Lett. 2006, 8, 1225–1228. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, C.; Swenson, D.C.; Gloer, J.B.; Wicklow, D.T.; Dowd, P.F. Novel antiinsectan oxalicine alkaloids from two undescribed fungicolous Penicillium spp. Org. Lett. 2003, 5, 773–776. [Google Scholar] [CrossRef]
- Li, C.; Gloer, J.B.; Wicklow, D.T.; Dowd, P.F. Antiinsectan decaturin and oxalicine analogues from Penicillium thiersii. J. Nat. Prod. 2005, 68, 319–322. [Google Scholar] [CrossRef]
- Abe, M.; Imai, T.; Ishii, N.; Usui, M.; Okuda, T.; Oki, T. Isolation of an insecticidal compound oxalicine B from Penicillium sp. TAMA 71 and confirmation of its chemical structure by X-ray crystallographic analysis. J. Pest. Sci. 2007, 32, 124–127. [Google Scholar] [CrossRef]
- Hayashi, H.; Matsumoto, H.; Akiyama, K. New insecticidal compounds, communesins C, D and E, from Penicillium expansum Link MK-57. Biosci. Biotechnol. Biochem. 2004, 68, 753–756. [Google Scholar] [CrossRef]
- Bai, M.; Zheng, C.-J.; Nong, X.-H.; Zhou, X.-M.; Luo, Y.-P.; Chen, G.-Y. Four new insecticidal xanthene derivatives from the mangrove-derived fungus Penicillium sp. JY246. Mar. Drugs 2019, 17, 649. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.; Zheng, C.-J.; Huang, G.-L.; Mei, R.-Q.; Wang, B.; Luo, Y.-P.; Zheng, C.; Niu, Z.-G.; Chen, G.-Y. Bioactive meroterpenoids and isocoumarins from the mangrove-derived fungus Penicillium sp. TGM112. J. Nat. Prod. 2019, 82, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.; Huang, G.-L.; Mei, R.-Q.; Wang, B.; Luo, Y.-P.; Nong, X.-H.; Chen, G.-Y.; Zheng, C.-J. Bioactive lactones from the mangrove-derived fungus Penicillium sp. TGM112. Mar. Drugs 2019, 17, 433. [Google Scholar] [CrossRef] [PubMed]
- Geris, R.; Rodrigues-Fo, E.; Garcia da Silva, H.H.; Garcia da Silva, I. Larvicidal effects of fungal meroterpenoids in the control of Aedes aegypti L., the main vector of dengue and yellow fever. Chem. Biodivers. 2008, 5, 341–345. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, W.; Xu, Z.; Bai, Q.; Zhou, X.; Zheng, C.; Bai, M.; Chen, G. Biological secondary metabolites from the Lumnitzera littorea-derived fungus Penicillium oxalicum HLLG-13. Mar. Drugs 2023, 21, 22. [Google Scholar] [CrossRef]
- Cantín, Á.; Moya, P.; Castillo, M.-A.; Primo, J.; Miranda, M.A.; Primo-Yúfera, E. Isolation and synthesis of N-(2-methyl-3-oxodec-8-enoyl)-2-pyrroline and 2-(hept-5-enyl)-3-methyl-4-oxo-6,7,8,8a-tetrahydro-4h-pyrrolo[2,1-b]1,3-oxazine—Two new fungal metabolites with in vivo anti-juvenile-hormone and insecticidal activity. Eur. J. Org. Chem. 1999, 221–226. [Google Scholar] [CrossRef]
- Lopez-Gresa, M.P.; Gonzalez, M.C.; Ciavatta, L.; Ayala, I.; Moya, P.; Primo, J. Insecticidal activity of paraherquamides, including paraherquamide H and paraherquamide I, two new alkaloids isolated from Penicillium cluniae. J. Agric. Food Chem. 2006, 54, 2921–2925. [Google Scholar] [CrossRef]
- Wang, H.J.; Gloer, J.B.; Wicklow, D.T.; Dowd, P.F. Aflavinines and other antiinsectan metabolites from the ascostromata of Eupenicillium crustaceum and related species. Appl. Environ. Microbiol. 1995, 61, 4429–4435. [Google Scholar] [CrossRef]
- Abe, M.; Imai, T.; Ishii, N.; Usui, M.; Okuda, T.; Oki, T. Quinolactacide, a new quinolone insecticide from Penicillium citrinum Thom F 1539. Biosci. Biotechnol. Biochem. 2005, 69, 1202–1205. [Google Scholar] [CrossRef]
- Shao, C.-L.; Wang, C.-Y.; Gu, Y.-C.; Wei, M.-Y.; Pan, J.-H.; Deng, D.-S.; She, Z.-G.; Lin, Y.-C. Penicinoline, a new pyrrolyl 4-quinolinone alkaloid with an unprecedented ring system from an endophytic fungus Penicillium sp. Bioorg. Med. Chem. Lett. 2010, 20, 3284–3286. [Google Scholar] [CrossRef]
- Shields, M.S.; Lingg, A.J.; Heimsch, R.C. Identification of a Penicillium urticae metabolite which inhibits Beauveria bassiana. J. Invert. Pathol. 1981, 38, 374–377. [Google Scholar] [CrossRef]
- Castillo, M.A.; Moya, P.; Cantín, A.; Miranda, M.A.; Primo, J.; Hernández, E.; Primo-Yúfera, E. Insecticidal, anti-juvenile hormone, and fungicidal activities of organic extracts from different Penicillium species and their isolated active components. J. Agric. Food Chem. 1999, 47, 2120–2124. [Google Scholar] [CrossRef]
- Khokhar, I.; Mukhtar, I.; Mushtaq, S. Antifungal effect of Penicillium metabolites against some fungi. Archiv. Phytopathol. Plant Prot. 2011, 44, 1347–1351. [Google Scholar] [CrossRef]
- Klowden, M.J.; Pallai, S.R. Physiological Systems in Insects, 3rd ed.; Academic Press: San Diego, CA, USA, 2022; 682p. [Google Scholar]
- Jeschke, P. Status and outlook for acaricide and insecticide discovery. Pest Manag. Sci. 2021, 77, 64–76. [Google Scholar] [CrossRef]
- Nation, J.L.N. Insect Physiology and Biochemistry; CRC Press: Boca Raton, FL, USA, 2015; 586p. [Google Scholar]
- Silman, I.; Sussman, J.L. Acetylcholinesterase: How is structure related to function? Chemico-Biol. Interact. 2008, 175, 3–10. [Google Scholar] [CrossRef]
- Peter, R.; Josende, M.E.; da Silva Barreto, J.; da Costa Silva, D.G.; da Rosa, C.E.; Maciel, F.E. Effect of Illicium verum (Hook) essential oil on cholinesterase and locomotor activity of Alphitobius diaperinus (Panzer). Pest. Biochem. Physiol. 2022, 181, 105027. [Google Scholar] [CrossRef]
- Nillos, M.G.; Rodriguez-Fuentes, G.; Gan, J.; Schlenk, D. Enantioselective acetylcholinesterase inhibition of the organophosphorous insecticides profenofos, fonofos, and crotoxyphos. Environ. Toxicol. Chem. 2007, 26, 1949–1954. [Google Scholar] [CrossRef]
- Thapa, S.; Lv, M.; Xu, H. Acetylcholinesterase: A primary target for drugs and insecticides. Mini Rev. Med. Chem. 2017, 17, 1665–1676. [Google Scholar] [CrossRef]
- Su, J.; Liu, H.; Guo, K.; Chen, L.; Yang, M.; Chen, Q. Research advances and detection methodologies for microbe-derived acetylcholinesterase inhibitors: A systemic review. Molecules 2017, 22, 176. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres Jr, V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Marston, A.; Kissling, J.; Hostettmann, K. A Rapid TLC Bioautographic method for the detection of acetylcholinesterase and butyrylcholinesterase inhibitors in plants. Phytochem. Anal. 2002, 13, 51–54. [Google Scholar] [CrossRef] [PubMed]
- Ingkaninan, K.; De Best, C.M.; Van Der Heijden, R.; Hofte, A.J.P.; Karabatak, B.; Irth, H.; Tjaden, U.R.; Van der Greef, J.; Verpoorte, R. High-performance liquid chromatography with on-line coupled UV, mass spectrometric and biochemical detection for identification of acetylcholinesterase inhibitors from natural products. J. Chromatogr. A 2000, 872, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Sandoval, I.T.; Cai, S.; Smith, K.A.; Delacruz, R.G.C.; Boyd, K.A.; Mills, J.J.; Jones, D.A.; Cichewicz, R.H. Cholinesterase inhibitory arisugacins L–Q from a Penicillium sp. isolate obtained through a citizen science initiative and their activities in a phenotype-based zebrafish assay. J. Nat. Prod. 2019, 82, 2627–2637. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.C.; Ghosh, J.J. Effect of citreoviridin, a mycotoxin from Penicillium citreoviride, on kinetic constants of acetylcholinesterase and ATPase in synaptosomes and microsomes from rat brain. Toxicon 1981, 19, 555–562. [Google Scholar] [CrossRef]
- Houghton, P.J.; Ren, Y.; Howes, M.-J. Acetylcholinesterase inhibitors from plants and fungi. Nat. Prod. Rep. 2006, 23, 181–199. [Google Scholar] [CrossRef]
- Marucci, G.; Buccioni, M.; Dal Ben, D.; Lambertucci, C.; Volpini, R.; Amenta, F. Efficacy of acetylcholinesterase inhibitors in Alzheimer’s disease. Neuropharmacology 2021, 190, 108352. [Google Scholar] [CrossRef]
- Wang, Y.; Zeng, Q.G.; Zhang, Z.B.; Yan, R.M.; Wang, L.Y.; Zhu, D. Isolation and characterization of endophytic huperzine A-producing fungi from Huperzia serrata. J. Ind. Microbiol. Biotechnol. 2011, 38, 1267–1278. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, J.-Y.; Qian, C.; Fang, Q.; Ye, G.-Y. Venom of the parasitoid wasp Pteromalus puparum contains an odorant binding protein. Arch. Insect Biochem. Physiol. 2015, 88, 101–110. [Google Scholar] [CrossRef]
- Wang, C.-H.; Zhang, C.; Xing, X.-H. Xanthine dehydrogenase: An old enzyme with new knowledge and prospects. Bioengineered 2016, 7, 395–405. [Google Scholar] [CrossRef]
- Nie, Y.; Yang, W.; Liu, Y.; Yang, J.; Lei, X.; Gerwick, W.H.; Zhang, Y. Acetylcholinesterase inhibitors and antioxidants mining from marine fungi: Bioassays, bioactivity coupled LC–MS/MS analyses and molecular networking. Mar. Life Sci. Technol. 2020, 2, 386–397. [Google Scholar] [CrossRef]
- Friedli, M.J.; Inestrosa, N.C. Huperzine A and its neuroprotective molecular signaling in Alzheimer’s disease. Molecules 2021, 26, 6531. [Google Scholar] [CrossRef]
- Thi Minh Le, T.; Thi Hong Hoang, A.; Thi Bich Le, T.; Thi Bich Vo, T.; Van Quyen, D.; Hoang Chu, H. Isolation of endophytic fungi and screening of huperzine A–producing fungus from Huperzia serrata in Vietnam. Sci. Rep. 2019, 9, 16152. [Google Scholar] [CrossRef]
- Kang, X.; Liu, C.; Liu, D.; Zeng, L.; Shi, Q.; Qian, K.; Xie, B. The complete mitochondrial genome of huperzine A-producing endophytic fungus Penicillium polonicum. Mitochondrial DNA Part B 2016, 1, 202–203. [Google Scholar] [CrossRef]
- Omura, S.; Kuno, F.; Otoguro, K.; Sunazuka, T.; Shiomi, K.; Masuma, R.; Iwai, Y. Arisugacin, a novel and selective inhibitor of acetylcholinesterase from Penicillium sp. FO-4259. J. Antibiot. 1995, 48, 745–746. [Google Scholar] [CrossRef]
- Hu, J.; Okawa, H.; Yamamoto, K.; Oyama, K.; Mitomi, M.; Anzai, H. Characterization of two cytochrome P450 monooxygenase genes of the pyripyropene biosynthetic gene cluster from Penicillium coprobium. J. Antibiot. 2011, 64, 221–227. [Google Scholar] [CrossRef]
- Kandasamy, R.; London, D.; Stam, L.; von Deyn, W.; Zhao, X.; Salgado, V.L.; Nesterov, A. Afidopyropen: New and potent modulator of insect transient receptor potential channels. Insect Biochem. Mol. Biol. 2017, 84, 32–39. [Google Scholar] [CrossRef]
- Horikoshi, R.; Goto, K.; Mitomi, M.; Oyama, K.; Hirose, T.; Sunazuka, T.; Ōmura, S. Afidopyropen, a novel insecticide originating from microbial secondary extracts. Sci. Rep. 2022, 12, 2827. [Google Scholar] [CrossRef]
- Campora, M.; Francesconi, V.; Schenone, S.; Tasso, B.; Tonelli, M. Journey on naphthoquinone and anthraquinone derivatives: New insights in Alzheimer’s disease. Pharmaceuticals 2021, 14, 33. [Google Scholar] [CrossRef]
- Vanessa, V.V.; Mah, S.H. Xanthone: Potential acetylcholinesterase inhibitor for Alzheimer’s disease treatment. Mini Rev. Med. Chem. 2021, 21, 2507–2529. [Google Scholar] [CrossRef]
- Sharma, K. Chromone scaffolds in the treatment of Alzheimer’s and Parkinson’s disease: An overview. ChemistrySelect 2022, 7, e202200540. [Google Scholar] [CrossRef]
- Sunazuka, T.; Handa, M.; Nagai, K.; Shirahata, T.; Harigaya, Y.; Otoguro, K.; Kuwajima, I.; Ōmura, S. Absolute Stereochemistries and total synthesis of (+)-arisugacins A and B, potent, orally bioactive and selective inhibitors of acetylcholinesterase. Tetrahedron 2004, 60, 7845–7859. [Google Scholar] [CrossRef]
- Li, H.-T.; Sun, Y.; Xie, F.; Wang, M.; Chen, J.-Y.; Zhou, H.; Ding, Z.-T. Xanthene and citrinin derivatives from the endophytic fungus Penicillium sp. T2-11. Tetrahedron Lett. 2022, 90, 153626. [Google Scholar] [CrossRef]
- Huang, X.-S.; Yang, B.; Sun, X.-F.; Xia, G.-P.; Liu, Y.-Y.; Ma, L.; She, Z.-G. A new polyketide from the mangrove endophytic fungus Penicillium sp. Sk14JW2P. Helv. Chim. Acta 2014, 97, 664–668. [Google Scholar] [CrossRef]
- He, J.-B.; Ji, Y.-N.; Hu, D.-B.; Zhang, S.; Yan, H.; Liu, X.-C.; Luo, H.-R.; Zhu, H.-J. Structure and absolute configuration of penicilliumine, a new alkaloid from Penicillium commune 366606. Tetrahedron Lett. 2014, 55, 2684–2686. [Google Scholar] [CrossRef]
- Kim, W.-G.; Song, N.-K.; Yoo, I.-D. Quinolactacins Al and A2, new acetylcholinesterase inhibitors from Penicillium citrinum. J. Antibiot. 2001, 54, 831–835. [Google Scholar] [CrossRef] [PubMed]
- Gan, D.; Zhu, L.; Zhang, X.-R.; Li, C.-Z.; Wang, C.-Y.; Cai, L.; Ding, Z.-T. Penaloidines A and B: Two unprecedented pyridine alkaloids from Penicillium sp. KYJ-6. Org. Chem. Front. 2022, 9, 2405–2411. [Google Scholar] [CrossRef]
- Hu, Y.-W.; Chen, W.-H.; Song, M.-M.; Pang, X.-Y.; Tian, X.-P.; Wang, F.-Z.; Liu, Y.-H.; Wang, J.-F. Indole diketopiperazine alkaloids and aromatic polyketides from the Antarctic fungus Penicillium sp. SCSIO 05705. Nat. Prod. Res. 2023, 37, 389–396. [Google Scholar] [CrossRef]
- Xu, F.; Chen, W.; Ye, Y.; Qi, X.; Zhao, K.; Long, J.; Pang, X.; Liu, Y.; Wang, J. A new quinolone and acetylcholinesterase inhibitors from a sponge-associated fungus Penicillium sp. SCSIO41033. Nat. Prod. Res. 2022. [Google Scholar] [CrossRef]
- Kuno, F.; Otoguro, K.; Shiomi, K.; Iwai, Y.; Omura, S. Arisugacins A and B, novel and selective acetylcholinesterase inhibitors from Penicillium sp. FO-4259 I. Screening, taxonomy, fermentation, isolation and biological activity. J. Antibiot. 1996, 49, 742–747. [Google Scholar] [CrossRef]
- Otoguro, K.; Shiomi, K.; Yamaguchi, Y.; Arai, N.; Sunazuka, T.; Masuma, R.; Iwai, Y.; Omura, S. Arisugacins C and D, novel acetylcholinesterase inhibitors and their related novel metabolites produced by Penicillium sp. FO-4259-11. J. Antibiot. 2000, 53, 50–57. [Google Scholar] [CrossRef]
- Ding, B.; Wang, Z.; Huang, X.; Liu, Y.; Chen, W.; She, Z. Bioactive α-pyrone meroterpenoids from mangrove endophytic fungus Penicillium sp. Nat. Prod. Res. 2016, 30, 2805–2812. [Google Scholar] [CrossRef]
- Huang, X.; Sun, X.; Ding, B.; Lin, M.; Liu, L.; Huang, H.; She, Z. A new anti-acetylcholinesterase α-pyrone meroterpene, arigsugacin I, from mangrove endophytic fungus Penicillium sp. Sk5GW1L of Kandelia candel. Planta Med. 2013, 79, 1572–1575. [Google Scholar] [CrossRef]
- Wang, Z.; Zeng, Y.; Zhao, W.; Dai, H.; Chang, W.; Lv, F. Structures and biological activities of brominated azaphilones produced by Penicillium sclerotiorum E23Y–1A. Phytochem. Lett. 2022, 52, 138–142. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Silva, G.H.; Regasini, L.O.; Zanardi, L.M.; Evangelista, A.H.; Young, M.C.M.; Bolzani, V.S.; Araujo, A.R. Bioactive metabolites produced by Penicillium sp.1 and sp.2, two endophytes associated with Alibertia macrophylla (Rubiaceae). Zeit. Naturforsch. C 2009, 64, 824–830. [Google Scholar] [CrossRef]
- Hemtasin, C.; Kanokmedhakul, S.; Moosophon, P.; Soytong, K.; Kanokmedhakul, K. Bioactive azaphilones from the fungus Penicillium multicolor CM01. Phytochem. Lett. 2016, 16, 56–60. [Google Scholar] [CrossRef]
- Huang, H.; Feng, X.; Xiao, Z.; Liu, L.; Li, H.; Ma, L.; Lu, Y.; Ju, J.; She, Z.; Lin, Y. Azaphilones and p-terphenyls from the mangrove endophytic fungus Penicillium chermesinum (ZH4-E2) isolated from the South China Sea. J. Nat. Prod. 2011, 74, 997–1002. [Google Scholar] [CrossRef]
- Liu, Y.-F.; Zhong, R.; Cao, F.; Wang, C.; Wang, C.-Y. Citrinin derivatives and unusual C25 steroids from a sponge-derived Penicillium sp. fungus. Chem. Nat. Compds. 2016, 52, 548–551. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Regasini, L.O.; Silva, G.H.; Pfenning, L.H.; Young, M.C.M.; Berlinck, R.G.S.; Bolzani, V.S.; Araujo, A.R. Dihydroisocoumarins produced by Xylaria sp. and Penicillium sp., endophytic fungi associated with Piper aduncum and Alibertia macrophylla. Phytochem. Lett. 2011, 4, 93–96. [Google Scholar] [CrossRef]
- Yuan, W.-H.; Wei, Z.-W.; Dai, P.; Wu, H.; Zhao, Y.-X.; Zhang, M.-M.; Jiang, N.; Zheng, W.-F. Halogenated metabolites isolated from Penicillium citreonigrum. Chem. Biodivers. 2014, 11, 1078–1087. [Google Scholar] [CrossRef]
- Kong, F.D.; Zhou, L.M.; Ma, Q.Y.; Huang, S.Z.; Wang, P.; Dai, H.-F.; Zhao, Y.-X. Penicillars A–E from the marine animal endogenic fungus Penicillium sp. SCS-KFD08. Phytochem. Lett. 2016, 17, 59–63. [Google Scholar] [CrossRef]
- Schürmann, B.T.M.; Sallum, W.S.T.; Takahashi, J.A. Austin, dehydroaustin and other metabolites from Penicillium brasilianum. Quím. Nova 2010, 33, 1044–1046. [Google Scholar] [CrossRef]
- Chen, C.-M.; Chen, W.-H.; Pang, X.-Y.; Liao, S.-R.; Wang, J.-F.; Lin, X.-P.; Yang, B.; Zhou, X.-F.; Luo, X.-W.; Liu, Y.-H. Pyrrolyl 4-quinolone alkaloids from the mangrove endophytic fungus Penicillium steckii SCSIO 41025: Chiral resolution, configurational assignment, and enzyme inhibitory activities. Phytochemistry 2021, 186, 112730. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-S.; Tan, M.-H.; Lv, M.-M.; Liu, C.-X.; Guo, Z.-Y.; Zhang, L.; Zou, K.; Chen, H.-Y.; Proksch, P. New γ-pyranone and nucleoside derivatives from Penicillium sp. Phytochem. Lett. 2017, 20, 285–288. [Google Scholar] [CrossRef]
- Li, C.; Shao, Y.; Li, W.; Yin, T.; Li, H.; Yan, H.; Guo, X.; Liu, B.; He, B. Hybrid diterpenic meroterpenoids from an endophytic Penicillium sp. induced by chemical epigenetic manipulation. J. Nat. Prod. 2022, 85, 1486–1494. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhu, T.; Ding, Y.; Khan, I.A.; Gu, Q.; Li, D. Sorbiterrin A, a novel sorbicillin derivative with cholinesterase inhibition activity from the marine-derived fungus Penicillium terrestre. Tetrahedron Lett. 2012, 53, 325–328. [Google Scholar] [CrossRef]
- Dowd, P.F.; Peng, F.-C.; Chen, J.W.; Ling, K.H. Toxicity and anticholinesterase activity of the fungal metabolites territrems to the corn earworm, Helicoverpa zea. Entomol. Exp. Appl. 1992, 65, 57–64. [Google Scholar] [CrossRef]
- Pang, X.; Zhou, X.; Lin, X.; Yang, B.; Tian, X.; Wang, J.; Xu, S.; Liu, Y. Structurally various sorbicillinoids from the deep-sea sediment derived fungus Penicillium sp. SCSIO06871. Bioorg. Chem. 2021, 107, 104600. [Google Scholar] [CrossRef]
- de Oliveira, G.P.; Barreto, D.L.C.; Ramalho Silva, M.; Augusti, R.; Evódio Marriel, I.; Gomes de Paula Lana, U.; Takahashi, J.A. Biotic stress caused by in vitro co-inoculation enhances the expression of acetylcholinesterase inhibitors by fungi. Nat. Prod. Res. 2022, 36, 4266–4270. [Google Scholar] [CrossRef]
- Lima, M.T.N.S.; dos Santos, L.B.; Bastos, R.W.; Nicoli, J.R.; Takahashi, J.A. Antimicrobial activity and acetylcholinesterase inhibition by extracts from chromatin modulated fungi. Braz. J. Microbiol. 2018, 49, 169–176. [Google Scholar] [CrossRef]
- Matsuda, K.; Buckingham, S.D.; Kleier, D.; Rauh, J.J.; Grauso, M.; Sattelle, D.B. Neonicotinoids: Insecticides acting on insect nicotinic acetylcholine receptors. Trends Pharmacol. Sci. 2001, 22, 573–580. [Google Scholar] [CrossRef]
- Oliveira, R.A.; Roat, T.C.; Carvalho, S.M.; Malaspina, O. Side-effects of thiamethoxam on the brain and midgut of the africanized honeybee Apis mellifera (Hymenopptera: Apidae). Environ. Toxicol. 2014, 29, 1122–1133. [Google Scholar] [CrossRef]
- Shi, J.; Liao, C.; Wang, Z.; Zeng, Z.; Wu, X. Effects of sublethal acetamiprid doses on the lifespan and memory-related characteristics of honey bee (Apis mellifera) workers. Apidologie 2019, 50, 553–563. [Google Scholar] [CrossRef]
- Hatjina, F.; Papaefthimiou, C.; Charistos, L.; Dogaroglu, T.; Bouga, M.; Emmanouil, C.; Arnold, G. Sublethal doses of imidacloprid decreased size of hypopharyngeal glands and respiratory rhythm of honeybees in vivo. Apidologie 2013, 44, 467–480. [Google Scholar] [CrossRef]
- Blacquière, T.; Smagghe, G.; van Gestel, C.A.M.; Mommaerts, V. Neonicotinoids in bees: A review on concentrations, side-effects and risk assessment. Ecotoxicology 2012, 21, 973–992. [Google Scholar] [CrossRef]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef]
- Giordani, G.; Cattabriga, G.; Becchimanzi, A.; Di Lelio, I.; De Leva, G.; Gigliotti, S.; Pennacchio, F.; Gargiulo, G.; Cavaliere, V. Role of neuronal and non-neuronal acetylcholine signaling in Drosophila humoral immunity. Insect Biochem. Mol. Biol. 2023, 153, 103899. [Google Scholar] [CrossRef]
- Hirata, K.; Kataoka, S.; Furutani, S.; Hayashi, H.; Matsuda, K. A fungal metabolite asperparaline a strongly and selectively blocks insect nicotinic acetylcholine receptors: The first report on the mode of action. PLoS ONE 2011, 6, e18354. [Google Scholar] [CrossRef]
- Kataoka, S.; Furutani, S.; Hirata, K.; Hayashi, H.; Matsuda, K. Three austin family compounds from Penicillium brasilianum exhibit selective blocking action on cockroach nicotinic acetylcholine receptors. NeuroToxicology 2011, 32, 123–129. [Google Scholar] [CrossRef]
- Hayashi, H.; Mukaihara, M.; Murao, S.; Arai, M.; Clardy, J. Acetoxydehydroaustin, a new bioactive compound, and related compound neoaustin from Penicillium sp. MG-11. Biosci., Biotechnol. Biochem. 1994, 58, 334–338. [Google Scholar] [CrossRef]
- Hosie, A.; Sattelle, D.; Aronstein, K.; ffrench-Constant, R. Molecular biology of insect neuronal GABA receptors. Trends Neurosci. 1997, 20, 578–583. [Google Scholar] [CrossRef]
- Watanabe, T.; Arisawa, M.; Narusuye, K.; Alam, M.S.; Yamamoto, K.; Mitomi, M.; Ozoe, Y.; Nishida, A. Alantrypinone and its derivatives: Synthesis and antagonist activity toward insect GABA receptors. Bioorg. Med. Chem. 2009, 17, 94–110. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Sakaguchi, A. Okaramine G, a new okaramine congener from Penicillium simplicissimum ATCC 90288. Biosci. Biotechnol. Biochem. 1998, 62, 804–806. [Google Scholar] [CrossRef]
- Hayashi, H.; Fujiwara, T.; Murao, S.; Arai, M. Okaramine C, a new insecticidal indole alkaloid from Penicillium simplicissimum. Agric. Biol. Chem. 1991, 55, 3143–3145. [Google Scholar] [CrossRef]
- Hayashi, H.; Asabu, Y.; Murao, S.; Arai, M. New okaramine congeners, okaramines D, E, and F, from Penicillium simplicissimum ATCC 90288. Biosci. Biotechnol. Biochem. 1995, 59, 246–250. [Google Scholar] [CrossRef]
- Shiono, Y.; Akiyama, K.; Hayashi, H. New okaramine congeners, okamines J, K, L, M and related ccmpounds, from Penicillium simplicissimum ATCC 90288. Biosci. Biotechnol. Biochem. 1999, 63, 1910–1920. [Google Scholar] [CrossRef]
- Shiono, Y.; Akiyama, K.; Hayashi, H. Okaramines N, O, P, Q and R, new okaramine congeners, from Penicillium simplicissimum ATCC 90288. Biosci. Biotechnol. Biochem. 2000, 64, 103–110. [Google Scholar] [CrossRef]
- Shiono, Y.; Akiyama, K.; Hayashi, H. Effect of the azetidine and azocine rings of okaramine B on insecticidal activity. Biosci. Biotechnol. Biochem. 2000, 64, 1519–1521. [Google Scholar] [CrossRef]
- Furutani, S.; Nakatani, Y.; Miura, Y.; Ihara, M.; Kai, K.; Hayashi, H.; Matsuda, K. GluCl a target of indole alkaloid okaramines: A 25 year enigma solved. Sci. Rep. 2014, 4, 6190. [Google Scholar] [CrossRef]
- Matsuda, K. Okaramines and other plant fungal products as new insecticide leads. Curr. Opin. Insect Sci. 2018, 30, 67–72. [Google Scholar] [CrossRef]
- Fujita, T.; Makishima, D.; Akiyama, K.; Hayashi, H. New convulsive compounds, brasiliamides A and B, from Penicillium brasilianum Batista JV-379. Biosci. Biotechnol. Biochem. 2002, 66, 1697–1705. [Google Scholar] [CrossRef]
- Riddiford, L.M. Juvenile hormone action: A 2007 perspective. J. Insect Physiol. 2008, 54, 895–901. [Google Scholar] [CrossRef]
- Konopova, B.; Jindra, M. Juvenile hormone resistance gene methoprene-tolerant controls entry into metamorphosis in the beetle Tribolium castaneum. Proc. Natl. Acad. Sci. USA 2007, 104, 10488–10493. [Google Scholar] [CrossRef]
- Cusson, M.; Sen, S.E.; Shinoda, T. Juvenile hormone biosynthetic enzymes as targets for insecticide discovery. In Advanced Technologies for Managing Insect Pests; Ishaaya, I., Palli, S.R., Horowitz, A.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 31–55. [Google Scholar]
- Nagaoka, S.; Hara, Y. Compactin. In Drug Discovery in Japan: Investigating the Sources of Innovation; Nagaoka, S., Ed.; Springer: Singapore, 2019; pp. 15–34. [Google Scholar]
- Nicoletti, R.; Buommino, E.; Antonietta Tufano, M. Patenting Penicillium strains. Recent Pat. Biotechnol. 2012, 6, 81–96. [Google Scholar] [CrossRef]
- Monger, D.J.; Lim, W.A.; Kézdy, F.J.; Law, J.H. Compactin inhibits insect HMG-CoA reductase and juvenile hormone biosynthesis. Biochem. Biophys. Res. Commun. 1982, 105, 1374–1380. [Google Scholar] [CrossRef]
- Hiruma, K.; Yagi, S.; Endo, A. ML-236B (Compactin) as an inhibitor of juvenile hormone biosynthesis. Appl. Entomol. Zool. 1983, 18, 111–115. [Google Scholar] [CrossRef]
- Edwards, J.P.; Price, N.R. Inhibition of juvenile hormone III biosynthesis in Periplaneta americana with the fungal metabolite compactin (ML-236B). Insect Biochem. 1983, 13, 185–189. [Google Scholar] [CrossRef]
- Bellés, X.; Camps, F.; Casas, J.; Lloria, J.; Messeguer, A.; Piulachs, M.D.; Sánchez, F.J. In vivo and in vitro effects of compactin in liposome carriers on juvenile hormone biosynthesis in adult females of Blattella germanica. Pest. Biochem. Physiol. 1988, 32, 1–10. [Google Scholar] [CrossRef]
- Castillo, M.; Moya, P.; Couillaud, F.; Garcerá, M.D.; Martínez-Pardo, R. A heterocyclic oxime from a fungus with anti-juvenile hormone activity. Arch. Insect Biochem. Physiol. 1998, 37, 287–294. [Google Scholar] [CrossRef]
- Moya, P.; Cantín, Á.; Castillo, M.-A.; Primo, J.; Miranda, M.A.; Primo-Yúfera, E. Isolation, structural assignment, and synthesis of N-(2-methyl-3-oxodecanoyl)-2-pyrroline, a new natural product from Penicillium brevicompactum with in vivo anti-juvenile hormone activity. J. Org. Chem. 1998, 63, 8530–8535. [Google Scholar] [CrossRef]
- Guo, X.-C.; Zhang, Y.-H.; Gao, W.-B.; Pan, L.; Zhu, H.-J.; Cao, F. Absolute configurations and chitinase inhibitions of quinazoline-containing diketopiperazines from the marine-derived fungus Penicillium polonicum. Mar. Drugs 2020, 18, 479. [Google Scholar] [CrossRef] [PubMed]
- Caccia, S.; Astarita, F.; Barra, E.; Di Lelio, I.; Varricchio, P.; Pennacchio, F. Enhancement of Bacillus thuringiensis toxicity by feeding Spodoptera littoralis larvae with bacteria expressing immune suppressive DsRNA. J. Pest. Sci 2020, 93, 303–314. [Google Scholar] [CrossRef]
- Nicoletti, R.; Becchimanzi, A. Ecological and molecular interactions between insects and fungi. Microorganisms 2022, 10, 96. [Google Scholar] [CrossRef]
- Feng, P.; Shang, Y.; Cen, K.; Wang, C. Fungal biosynthesis of the bibenzoquinone oosporein to evade insect immunity. Proc. Natl. Acad. Sci. USA 2015, 112, 11365–11370. [Google Scholar] [CrossRef]
- Wei, G.; Lai, Y.; Wang, G.; Chen, H.; Li, F.; Wang, S. Insect pathogenic fungus interacts with the gut microbiota to accelerate mosquito mortality. Proc. Natl. Acad. Sci. USA 2017, 114, 5994–5999. [Google Scholar] [CrossRef] [PubMed]
- Marmaras, V.J.; Charalambidis, N.D.; Zervas, C.G. Immune response in insects: The role of phenoloxidase in defense reactions in relation to melanization and sclerotization. Arch. Insect Biochem. Physiol. 1996, 31, 119–133. [Google Scholar] [CrossRef]
- Lu, Z.; Jiang, H. Regulation of phenoloxidase activity by high-and low-molecular-weight inhibitors from the larval hemolymph of Manduca sexta. Insect Biochem. Mol. Biol. 2007, 37, 478–485. [Google Scholar] [CrossRef]
- DellaGreca, M.; De Tommaso, G.; Salvatore, M.M.; Nicoletti, R.; Becchimanzi, A.; Iuliano, M.; Andolfi, A. The issue of misidentification of kojic acid with flufuran in Aspergillus flavus. Molecules 2019, 24, 1709. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Anstee, J.H. Phenoloxidase and its zymogen from the haemolymph of larvae of the Lepidopteran Spodoptera littoralis (Lepidoptera: Noctuidae). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1995, 110, 379–384. [Google Scholar] [CrossRef]
- Dowd, P.F. Relative inhibition of insect phenoloxidase by cyclic fungal metabolites from insect and plant pathogens. Nat. Toxins 1999, 7, 337–341. [Google Scholar] [CrossRef]
- Zhang, H.-H.; Luo, M.-J.; Zhang, Q.-W.; Cai, P.-M.; Idrees, A.; Ji, Q.-E.; Yang, J.-Q.; Chen, J.-H. Molecular characterization of prophenoloxidase-1 (PPO1) and the inhibitory effect of kojic acid on phenoloxidase (PO) activity and on the development of Zeugodacus tau (Walker) (Diptera: Tephritidae). Bull. Entomol. Res. 2019, 109, 236–247. [Google Scholar] [CrossRef]
- Christensen, M.; Frisvad, J.C.; Tuthill, D. Taxonomy of the Penicillium miczynskii group based on morphology and secondary metabolites. Mycol. Res. 1999, 103, 527–541. [Google Scholar] [CrossRef]
- Houbraken, J.; Frisvad, J.C.; Samson, R.A. Taxonomy of Penicillium section Citrina. Stud. Mycol. 2011, 70, 53–138. [Google Scholar] [CrossRef]
- Mc Namara, L.; Dolan, S.K.; Walsh, J.M.D.; Stephens, J.C.; Glare, T.R.; Kavanagh, K.; Griffin, C.T. Oosporein, an abundant metabolite in Beauveria caledonica, with a feedback induction mechanism and a role in insect virulence. Fungal Biol. 2019, 123, 601–610. [Google Scholar] [CrossRef] [PubMed]
- El-Ghany, N.M.A. Semiochemicals for controlling insect pests. J. Plant Prot. Res. 2019, 59, 1–11. [Google Scholar] [CrossRef]
- Kusari, S.; Verma, V.C.; Lamshoeft, M.; Spiteller, M. An endophytic fungus from Azadirachta indica A. Juss. that produces azadirachtin. World J. Microbiol. Biotechnol. 2012, 28, 1287–1294. [Google Scholar] [CrossRef]
- Stierle, D.B.; Stierle, A.A.; Ganser, B. New phomopsolides from a Penicillium sp. J. Nat. Prod. 1997, 60, 1207–1209. [Google Scholar] [CrossRef]
- Stierle, A.A.; Stierle, D.B.; Mitman, G.G.; Snyder, S.; Antczak, C.; Djaballah, H. Phomopsolides and related compounds from the alga-associated fungus, Penicillium clavigerum. Nat. Prod. Commun. 2014, 9, 87–90. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Smedsgaard, J.; Larsen, T.O.; Samson, R.A. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud. Mycol. 2004, 49, 201–241. [Google Scholar]
- Xu, Y.; Vinas, M.; Alsarrag, A.; Su, L.; Pfohl, K.; Rohlfs, M.; Schäfer, W.; Chen, W.; Karlovsky, P. Bis-naphthopyrone pigments protect filamentous Ascomycetes from a wide range of predators. Nat. Commun. 2019, 10, 3579. [Google Scholar] [CrossRef]
- Yin, G.; Zhao, H.; Pennerman, K.K.; Jurick, W.M.; Fu, M.; Bu, L.; Guo, A.; Bennett, J.W. Genomic analyses of Penicillium species have revealed patulin and citrinin gene clusters and novel loci involved in oxylipin production. J. Fungi 2021, 7, 743. [Google Scholar] [CrossRef]
- Yin, G.; Padhi, S.; Lee, S.; Hung, R.; Zhao, G.; Bennett, J.W. Effects of three volatile oxylipins on colony development in two species of fungi and on Drosophila larval metamorphosis. Curr. Microbiol. 2015, 71, 347–356. [Google Scholar] [CrossRef]
- Macedo, G.E.; de Brum Vieira, P.; Rodrigues, N.R.; Gomes, K.K.; Martins, I.K.; Franco, J.L.; Posser, T. Fungal compound 1-octen-3-ol induces mitochondrial morphological alterations and respiration dysfunctions in Drosophila melanogaster. Ecotoxicol. Environ. Safety 2020, 206, 111232. [Google Scholar] [CrossRef]
- Almaliki, H.S.; Angela, A.; Goraya, N.J.; Yin, G.; Bennett, J.W. Volatile organic compounds produced by human pathogenic fungi are toxic to Drosophila melanogaster. Front. Fungal Biol. 2021, 1, 629510. [Google Scholar] [CrossRef]
- Inamdar, A.A.; Hossain, M.M.; Bernstein, A.I.; Miller, G.W.; Richardson, J.R.; Bennett, J.W. Fungal-derived semiochemical 1-octen-3-ol disrupts dopamine packaging and causes neurodegeneration. Proc. Natl. Acad. Sci. USA 2013, 110, 19561–19566. [Google Scholar] [CrossRef]
- Inamdar, A.A.; Bennett, J.W. A common fungal volatile organic compound induces a nitric oxide mediated inflammatory response in Drosophila melanogaster. Sci. Rep. 2014, 4, 3833. [Google Scholar] [CrossRef]
- Stensmyr, M.C.; Dweck, H.K.M.; Farhan, A.; Ibba, I.; Strutz, A.; Mukunda, L.; Linz, J.; Grabe, V.; Steck, K.; Lavista-Llanos, S.; et al. A conserved dedicated olfactory circuit for detecting harmful microbes in Drosophila. Cell 2012, 151, 1345–1357. [Google Scholar] [CrossRef]
- Melo, N.; Wolff, G.H.; Costa-da-Silva, A.L.; Arribas, R.; Triana, M.F.; Gugger, M.; Riffell, J.A.; DeGennaro, M.; Stensmyr, M.C. Geosmin attracts Aedes aegypti mosquitoes to oviposition sites. Curr. Biol. 2020, 30, 127–134. [Google Scholar] [CrossRef]
- Gu, F.; Ai, S.; Chen, Y.; Jin, S.; Xie, X.; Zhang, T.; Zhong, G.; Yi, X. Mutualism promotes insect fitness by fungal nutrient compensation and facilitates fungus propagation by mediating insect oviposition preference. ISME J. 2022, 16, 1831–1842. [Google Scholar] [CrossRef]
- Guo, H.-G.; Han, C.-Y.; Zhang, A.-H.; Yang, A.-Z.; Qin, X.-C.; Zhang, M.-Z.; Du, Y.-L. Penicillium fungi mediate behavioral responses of the yellow peach moth, Conogethes punctiferalis (Guenée) to apple fruits via altering the emissions of host plant VOCs. Arch. Insect Biochem. Physiol. 2022, 110, e21895. [Google Scholar] [CrossRef]
- Chen, Y.; Han, J.; Yang, H.; Qin, X.; Guo, H.; Du, Y. Different maize ear rot fungi deter the oviposition of yellow peach moth (Conogethes punctiferalis (Guenée)) by maize volatile organic compounds. Agronomy 2023, 13, 251. [Google Scholar] [CrossRef]
- Nicoletti, R.; Fiorentino, A.; Scognamiglio, M. Endophytism of Penicillium species in woody plants. Open Mycol. J. 2014, 8, 1–26. [Google Scholar] [CrossRef]




| Species | Host | Country | Source | References |
|---|---|---|---|---|
| P. adametzioides | unspecified insect | Brazil | dead or live insect | [19] |
| P. atrofulvum | Triplectides sp. | Brazil | gut | [20] |
| P. aurantiocandidum | Vespula vulgaris | California (USA) | gut of larva | [21] |
| P. aurantiogriseum | Ostrinia nubilalis | Iowa (USA) | dead adult and larva | [22] |
| Schistocerca gregaria | Egypt | dead adult | [23] | |
| Spodoptera littoralis | Egypt | eggs | [24] | |
| Triatoma brasiliensis | Brazil | digestive tract | [25] | |
| P. brasilianum | unidentified insect | South Korea | dead insect | [26] |
| P. brevicompactum | Apis mellifera | Sweden | midgut | [27] |
| Bradysia agrestis | South Korea | gut | [28] | |
| Malacosoma neustria | Germany | diseased larvae | [29] | |
| P. brocae | Hypothenemus hempei | Mexico | cuticle and frass of females | [30] |
| P. cairnsense | Triplectides sp. | Brazil | gut | [20] |
| P. camponotum | Camponotus herculeanus | Germany | buccal cavities | [31] |
| Camponotus pennsylvanicus | Canada | |||
| P. canescens | Aedes sp. | Brazil | adult | [32] |
| P. caseifulvum | Triplectides sp. | Brazil | gut | [20] |
| P. chermesinum | Vespula pennsylvanica | California (USA) | gut of larva | [21] |
| P. chrysogenum | A. mellifera | Michigan (USA) | dead adult | [33] |
| Arizona (USA) | gut | [34,35] | ||
| Culex nigripalpus | Brazil | diseased adult | [36] | |
| Culex sp. | Brazil | adult | [32] | |
| Cydia ulicetana | New Zealand | exoskeleton of live adult | [37] | |
| S. littoralis | Egypt | pupae and adults | [24] | |
| unspecified insect | Brazil | dead or live insect | [19] | |
| P. cinnamopurpureum | Triatoma pseudomaculata | Brazil | digestive tract | [25] |
| P. citreonigrum | Sericothrips staphylinus | New Zealand | exoskeleton of live adult | [37] |
| T. brasiliensis | Brazil | digestive tract | [25] | |
| P. citrinum | Aedes aegypti | Australia | eggs | [38] |
| Antheraea mylitta | India | diseased larvae | [39] | |
| A. mellifera | Arizona (USA) | gut | [35] | |
| B. agrestis | South Korea | gut | [28] | |
| Culex quinquefasciatus | Thailand | dead adult | [40] | |
| Culex sp. | Brazil | adult | [32] | |
| H. hempei | Mexico | cuticle and gut of females | [30] | |
| Nomia melanderi | western USA | diseased larva | [41] | |
| Parastrongylus megistus | Brazil | digestive tract | [42] | |
| Pteroptyx bearni | Sabah (Malaysia) | eggs | [43] | |
| T. brasiliensis, T. infestans | Brazil | digestive tract | [25] | |
| Triplectides sp. | Brazil | gut | [20] | |
| unspecified insect | Brazil | internal mycobiota | [19] | |
| V. vulgaris | California (USA) | larva | [21] | |
| P. coffeae | B. agrestis | South Korea | gut | [28] |
| P. commune | A. mellifera | Michigan (USA) | dead adults | [33] |
| Hylobius abietis | Sweden | frass | [44] | |
| P. corylophilum | Anopheles darlingi, Culex declarator, C. nigripalpus, C. saltanensis, Mansonia titilans | Brazil | larvae, adults | [7,32,36] |
| A. mellifera | Michigan (USA) | dead adults | [33] | |
| Arizona (USA) | gut | [35] | ||
| Musca domestica | Brazil | diseased adult/larva | [36] | |
| P. megistus | Brazil | digestive tract | [42] | |
| T. infestans, T. sordida, T. vitticeps, T. brasiliensis, T. pseudomaculata | Brazil | digestive tract | [25,36] | |
| V. pennsylvanica | California (USA) | adult | [21] | |
| P. costaricense | Rothschildia lebeau | Costa Rica | gut | [31] |
| P. crustosum | H. hempei | Mexico | cuticle, frass, gut of female | [30] |
| P. cyclopium | A. mellifera | Michigan (USA) | dead adults | [33] |
| Arizona (USA) | gut | [35] | ||
| Ostrinia nubilalis | Iowa (USA) | dead adult and larva | [22] | |
| P. decumbens | Aedes sp., Anopheles sp., Culex sp., Mansonia sp. | Brazil | adult | [32] |
| Helicoverpa zea | Iowa (USA) | dead larva and pupa | [22] | |
| Ostrinia nubilalis | Iowa (USA) | dead larva | [22] | |
| P. megistus | Brazil | digestive tract | [42] | |
| T. brasiliensis | Brazil | digestive tract | [25] | |
| V. pennsylvanica | California (USA) | gut of larva | [21] | |
| P. euglaucum | A. mellifera | Italy | dead/live adults/larvae | [45] |
| N. melanderi | northwestern USA | diseased larvae | [46] | |
| P. excelsum | bees, ants | Brazil | adults | [47] |
| P. expansum | A. mellifera | Michigan (USA) | dead adults | [33] |
| Anopheles sp., Culex sp. | Brazil | adult | [32] | |
| H. abietis | Sweden | frass | [44,48] | |
| T. brasiliensis | Brazil | digestive tract | [25] | |
| P. exsudans | Triplectides sp. | Brazil | gut | [20] |
| P. fellutanum | Aedes scapularis | Brazil | diseased adult | [36] |
| C. quinquefasciatus | Brazil | larvae, adults | [7,32] | |
| P. megistus | Brazil | digestive tract | [42] | |
| T. brasiliensis, T. infestans | Brazil | digestive tract | [25] | |
| P. freii | unidentified Pyralidae | Lebanon | dead moth | [49] |
| P. fundyense | C. pennsylvanicus | Canada | buccal cavities | [31] |
| P. glabrum | A. mellifera | Arizona (USA) | gut | [34,35] |
| T. brasiliensis | Brazil | digestive tract | [25] | |
| P. gladioli | B. agrestis | South Korea | gut | [28] |
| P. griseofulvum | A. mellifera | Arizona (USA) | gut | [34] |
| T. infestans | Brazil | digestive tract | [25] | |
| Argentina | digestive tract | [50] | ||
| P. guanacastense | Eutelia sp. | Costa Rica | gut | [51] |
| P. herquei | Euops chinensis | China | mycangia | [15] |
| P. infrabuccatum | C. pennsylvanicus | Canada | buccal cavities | [31] |
| P. janthinellum | Aedes fluvialitis, A. darlingi, C. quinquefasciatus | Brazil | larvae, adults | [7,32] |
| Anopheles sp., C. nigripalpus | Brazil | diseased adult | [36] | |
| P. megistus | Brazil | digestive tract | [42] | |
| T. infestans, T. brasiliensis | Brazil | digestive tract | [25] | |
| P. lanosum | V. pennsylvanica | California (USA) | gut of larva | [21] |
| P. lividum | H. abietis | Sweden | frass | [44] |
| P. mallochii | Citheronia lobesis, R. lebeau | Costa Rica | gut | [51] |
| Triplectides sp. | Brazil | gut | [20] | |
| unspecified insect | Brazil | dead or live insect | [19] | |
| P. maximae | Triplectides sp. | Brazil | gut | [20] |
| P. miczynskii | T. infestans | Brazil | digestive tract | [25] |
| P. ochrochloron | A. mellifera | Arizona (USA) | gut | [34] |
| P. olsonii | H. hempei | Mexico | cuticle and gut of females | [30] |
| P. oxalicum | Acrida bicolor | China | gut | [52] |
| Bemisia tabaci | India | mummified adult | [53] | |
| Mansonia sp. | Brazil | adult | [32] | |
| P. palitans | A. mellifera | Michigan (USA) | live adults | [33] |
| P. paxilli | Triplectides sp. | Brazil | gut | [20] |
| P. phoeniceum | V. pennsylvanica | California (USA) | gut of larva | [21] |
| P. polonicum | Culex sp. | Lebanon | dead adult | [49] |
| Muljarus japonicus | South Korea | dead adult | [54] | |
| P. restrictum | A. aegypti | Australia | eggs | [38] |
| V. pennsylvanica | California (USA) | gut of larva | [21] | |
| P. rolfsii | Triplectides sp. | Brazil | gut | [20] |
| P. roseopurpureum | A. aegypti | Australia | eggs | [38] |
| P. rubens | Triplectides sp. | Brazil | gut | [20] |
| P. rubidurum | B. agrestis | South Korea | gut | [28] |
| P. simplicissimum | A. aegypti | Australia | eggs | [38] |
| P. solitum | H. abietis | Sweden | frass | [44] |
| P. soppii | Formica polyctena | Poland | worker | [55] |
| Penicillium sp. | Acromyrmex balzani, A. niger, A. rugosus, A. subterraneus | Brazil | workers | [56] |
| A. aegypti | Australia | eggs | [38] | |
| A. darlingi | Brazil | diseased adult | [36] | |
| A. mellifera | Costa Rica | diseased larvae | [57] | |
| Atta colombica | Panama | queen cuticle | [58] | |
| Bombus sp. | Denmark | dead adult | [59] | |
| Carpophilus dimidiatus, Catarthus quadricollis, Cryptolestes ferrugineus, Gnathocerus cornutus, Palorus subdepressus, Prostephanus truncatus, Sitophilus zeamais, Tribolium castaneum | Benin | adults | [60] | |
| C. quinquefasciatus | Thailand | dead adult | [40] | |
| Diaphania (Margaronia) pyloalis | Japan | larva | [61] | |
| Euops lespedezae | Japan | mycangia | [62] | |
| F. polyctena | Poland | workers | [55] | |
| Halictus rubicundus | India | frass | [63] | |
| Lasioglossum zephyrum | India | dead larvae | [63] | |
| Lixus impressiventris | South Korea | dead insect | [54] | |
| M. domestica | Iran | adults | [64] | |
| M. domestica | Brazil | diseased adult/larva | [36] | |
| Periplaneta americana | Sumatra (Indonesia) | adult | [65] | |
| Platypus cylindrus | Portugal | exoskeleton, gut, mycangia | [66] | |
| Reticulitermes flavipes | Ontario (Canada) | live termites | [67] | |
| Scaptocoris carvalhoi | Brazil | adult or nymph | [68] | |
| T. brasiliensis, T. pseudomaculata, T. vitticeps | Brazil | digestive tract | [25] | |
| T. infestans | Argentina | digestive tract | [50] | |
| Tribolium castaneum | India | adults | [69] | |
| Zonocerus variegatus | Nigeria | dead adult | [70] | |
| P. spinulosum | H. abietis | Sweden | feces and frass | [44] |
| T. brasiliensis, T. pseudomaculata | Brazil | digestive tract | [25] | |
| P. steckii | Meloe proscarabaeus | South Korea | dead adult | [54] |
| P. megistus | Brazil | digestive tract | [42] | |
| T. sordida | Brazil | digestive tract | [25] | |
| V. vulgaris | California (USA) | larva | [21] | |
| P. vancouverense | Triplectides sp. | Brazil | gut | [20] |
| P. viridicatum | A. darlingi | Brazil | larvae, adults | [7,32] |
| T. brasiliensis | Brazil | digestive tract | [25] | |
| P. waksmanii | Aedes sp., Anopheles sp., C. quinquefasciatus, M. titilans | Brazil | larvae, adults | [7,32,71] |
| P. megistus | Brazil | digestive tract | [42] | |
| T. brasiliensis, T. pseudomaculata, T. vitticeps | Brazil | digestive tract | [25] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicoletti, R.; Andolfi, A.; Becchimanzi, A.; Salvatore, M.M. Anti-Insect Properties of Penicillium Secondary Metabolites. Microorganisms 2023, 11, 1302. https://doi.org/10.3390/microorganisms11051302
Nicoletti R, Andolfi A, Becchimanzi A, Salvatore MM. Anti-Insect Properties of Penicillium Secondary Metabolites. Microorganisms. 2023; 11(5):1302. https://doi.org/10.3390/microorganisms11051302
Chicago/Turabian StyleNicoletti, Rosario, Anna Andolfi, Andrea Becchimanzi, and Maria Michela Salvatore. 2023. "Anti-Insect Properties of Penicillium Secondary Metabolites" Microorganisms 11, no. 5: 1302. https://doi.org/10.3390/microorganisms11051302
APA StyleNicoletti, R., Andolfi, A., Becchimanzi, A., & Salvatore, M. M. (2023). Anti-Insect Properties of Penicillium Secondary Metabolites. Microorganisms, 11(5), 1302. https://doi.org/10.3390/microorganisms11051302